|
SEMINARIO 12 |
CINETICA ENZIMATICA III
Regulación de la Actividad Enzimática: Enzimas Alostéricas y Modificación Covalente.
1) Un dímero alostérico tiene un factor de interacción a de 0,2 cuando es analizado de acuerdo al modelo de interacción secuencial (es decir, la unión de la primera molécula de sustrato incrementa la constante de unión al sitio vacante en un factor de 5).
a) ¿Cuál es la distribución relativa de las distintas formas de la enzima a una S = 0,3 Ks?
b) ¿Cuál es la velocidad específica a S = 0,3 Ks?
c) ¿El valor de nap calculado es igual a 2?
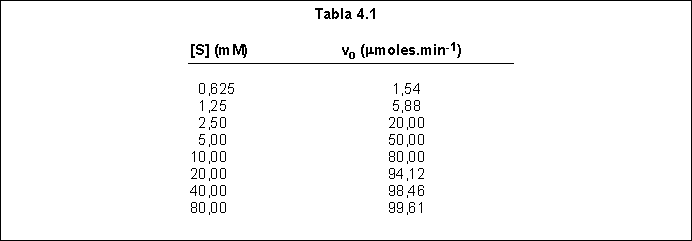
2) A partir de los datos de la Tabla 4.1, determine si la enzima que cataliza esta reacción obedece a una cinética hiperbólica o sigmoidal y calcule o estime las constantes cinéticas apropiadas (Km y Vmax o K', S0,5, nap Vmax)
3) Calcule el valor de nap de una enzima alostérica si S0,9 / S0,1 adquiere los siguientes valores: 4,0; 6,5; 9,0. ¿Qué le indican estos cálculos?
4) La Tabla 4.2 contiene datos para las regiones intermedias de las curvas de unión de oxígeno de tres hemoglobinas en solución bajo condiciones idénticas. La concentración de oxígeno se reporta como presión parcial de oxígeno, pO2, y la fracción de la velocidad máxima de unión (vo/Vmax) como la fracción de saturación, Y. Las tres hemoglobinas son: A (hemoglobina humana adulta normal), Hiroshima (His146b ® Asp) y Kansas (Asp102b ® Thr)
Grafique estos datos como para obtener los valores de nap de cada hemoglobina e interprete los efectos de la cooperatividad de la unión del oxígeno en términos de los modelos que Ud. conoce, sabiendo que las mutaciones mencionadas se encuentran en las interfaces de unión de las subunidades de hemoglobina y no en los sitios de unión del oxígeno.
TABLA 4.2
|
A |
Hiroshima |
Kansas |
|||
|
PO2 |
Y |
PO2 |
Y |
PO2 |
Y |
|
0.67 |
0.044 |
0.133 |
0.045 |
1.33 |
0.193 |
|
1.33 |
0.243 |
0.267 |
0.179 |
2.67 |
0.371 |
|
2.00 |
0.500 |
0.667 |
0.620 |
4.00 |
0.500 |
|
2.67 |
0.691 |
1.330 |
0.822 |
5.33 |
0.592 |
|
4.00 |
0.874 |
2.00 |
0.948 |
6.67 |
0.660 |
5) La bacteria Bordetella pertussis, agente causal de la tos convulsa, posee un sistema de dos componentes para inducir la expresión de sus genes de virulencia. El sistema se muestra en la Figura 4.1, donde BvgS representa a la proteína sensora y BvgA a la reguladora de respuesta. La proteína sensora BvgS contiene dominios distinguibles delineados por secuencias transmembrana (TM) y regiones ricas en alanina/prolina (A/P). Se muestra con una línea la porción citoplásmica de BvgS (´BvgS), que se ha utilizado para los experimentos que se detallan más abajo. También se muestran los aminoácidos conservados en los dominios transmisor y receptor, junto con sus posiciones, para BvgS y BvgA. En el dominio C- terminal de BvgS se encuentra una histidina conservada en la posición 1172 y en el C- terminal de BvgA, un motivo hélice-vuelta-hélice (helix-turn-helix, HTH).
Para determinar los sitios de fosforilación y los requerimientos estructurales para la transmisión de la señal, se realizaron dos experimentos. En la Fig. 4.2 se muestra el resultado de un PAGE-SDS donde se separó los productos de una incubación de ´BvgS con ciertos péptidos en presencia de [-32P]ATP. Los péptidos adicionados a la incubación fueron: la región C- terminal de BvgS (GST-C) correspondiente a una cepa salvaje (línea 2) o la misma región pero proveniente de una cepa donde se introdujo una mutación puntual en la histidina 1172, cambiándose a glutamina (línea 3). En la línea 1 se sembró ´BvgS sin el agregado de otro péptido, y en las líneas 4 y 5, los mismos péptidos que en 2 y 3 pero sin ´BvgS. El gel se reveló por autorradiografía.
En la Fig. 4.3 se muestra otra electroforesis revelada por autorradiografía donde ´BvgS se incubó en presencia de [-32P]ATP, sin péptidos (S) o con el agregado de BvgA (S+A) o con el agregado de la porción C- terminal de BvgS (S+C). Dado que se sabe que los enlaces fosfoamidil son estables al álcali y lábiles al ácido mientras que los enlaces fosfoacil son lábiles tanto al álcali como al ácido, las incubaciones se realizaron en buffer Tris 50 mM o en HCl 1N o en NaOH 3N para corroborar si la modificación covalente se realiza en residuos de histidina.
a) ¿En qué consiste un sistema de dos componentes? ¿Qué función cumple?
b) De acuerdo al resultado de la Fig. 4.2, explique cómo se modifica la región C- terminal de BvgS.
c) ¿Cómo corrobora la respuesta anterior con el resultado de la Fig. 4.3?. ¿Qué alternativas había?
d) Explique cómo BvgA produce la respuesta del sistema, especificando qué dominios intervienen y qué funciones cumplen.
EMBO J. 15: 1028-1036 (1996)
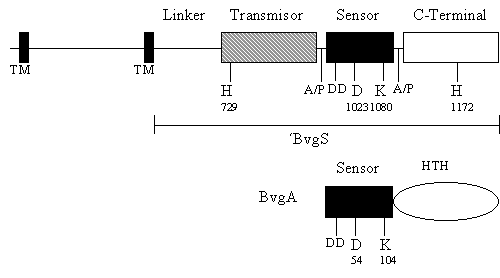
Figura 4.1
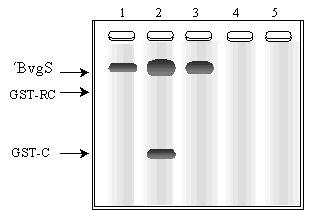
Figura 4.2
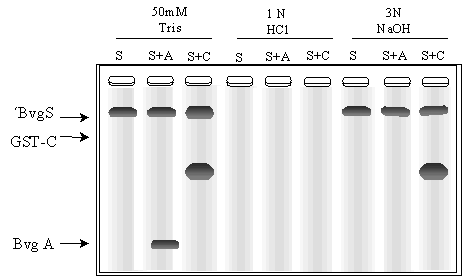
Figura 4.3
6) En un estudio del metabolismo de las plantas se aisló y caracterizó la sacarosa sintetasa de zanahoria, que cataliza la interconversión reversible de sacarosa + UDP para formar UDP-glucosa + fructosa (Fig. 4.4). La actividad de dicha enzima se mide a través de la formación de UDP-glucosa, la cual se cuantifica en presencia de un exceso de UDP-glucosa deshidrogenasa y NAD+, registrándose la formación de NADH. En la caracterización de dicha actividad, se realizaron experimentos para obtener los parámetros cinéticos y determinar la actividad inhibitoria de la fructosa (Tabla 4.3). También se determinó la temperatura óptima de la reacción (Fig. 4.5A) y la estabilidad térmica de la enzima (Fig.4.5B).
Tabla 4.3 Velocidad inicial (moles NADH/min) de la reacción de la sacarosa sintetasa con diversas concentraciones de sacarosa, en presencia o ausencia de fructosa 10 mM.

Además, mediciones de la actividad enzimática in vivo revelaron que la misma disminuye un 80% en 5 minutos si las plantas se pasan a una atmósfera de argón (ausencia de oxígeno).
a) Describa cómo se realiza la cuantificación de la actividad de esta enzima, qué reacciones ocurren y qué clase de mediciones se practican.
b) Explique cómo se realizaron los experimentos de la Figura 4.5 y qué información obtiene de ellos.
c) Calcule los parámetros cinéticos de la sacarosa sintetasa con los datos de la Tabla 4.3, como así también la Ki de la fructosa y explique qué clase de inhibidor es dicho metabolito, cómo actúa sobre la enzima y qué rol cree que cumple esta inhibición en el metabolismo carbonado de la zanahoria.
d) Explique por qué disminuyó la actividad enzimática en ausencia de oxígeno. Ayúdese con la Figura 4.4 para evaluar las siguientes posibilidades:
d1) En ausencia de oxígeno se activa un inhibidor de la expresión del gen que codifica para la sacarosa sintetasa, reprimiendo la transcripción del mismo.
d2) La fructosa ejerce una actividad inhibitoria por feedback
d3) Aumenta la concentración de un segundo mensajero que induce la fosforilación (o defosforilación) de la sacarosa sintetasa, que de esta manera pasa de un estado activo a uno inactivo.
d4) La enzima se inactiva por una proteólisis selectiva.
Diga si alguna de estas posibilidades es improbable dadas las condiciones en que se produce la inhibición y plantee experimentos (la menor cantidad posible) para detectar cuál opera en realidad.
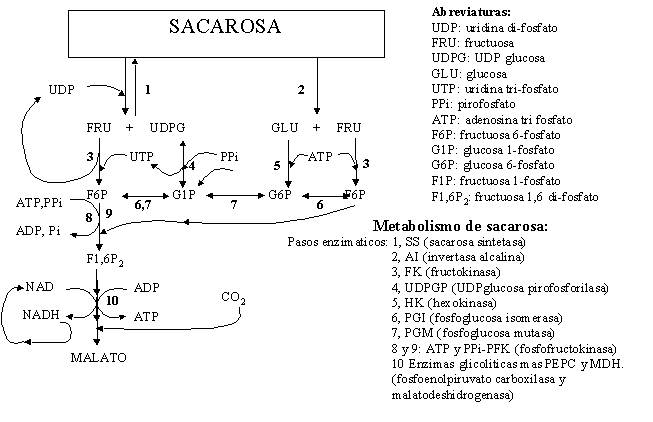
Figura 4.4
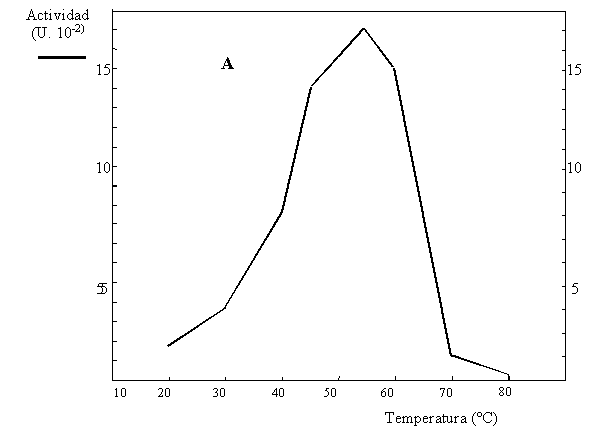
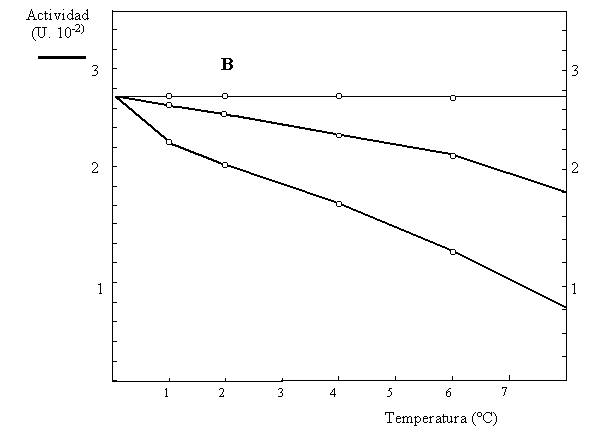
Figuras 4.5
7) En las plantas de soja la bacteria Bradyrhizobium japonicum forma nódulos radiculares dentro de los cuales es capaz de reducir el N2 atmosférico a NH3, el cual puede ser usado por la planta para cubrir todas sus necesidades de N. Dicha actividad sólo puede ser llevada a cabo por una forma muy diferenciada de los rizobios, llamada bacteroide. Los bacteroides dependen totalmente de la planta para sus requerimientos de C y energía, los que pueden ser satisfechos únicamente con succinato y malato. Para ello, la fosfoenol piruvato carboxilasa (PEPC) de origen vegetal presente en grandes cantidades en el nódulo juega un papel fundamental en convertir el fofoenol piruvato en oxalacetato, el cual luego va a ser transformado en malato por la malato deshidrogenasa. Para estudiar la regulación de la PEPC, preparaciones de PEPC parcialmente purificada de extractos de nódulos de soja se incubaron con 32P y se observó la fosforilación de PEPC por autorradiografía de geles de poliacrilamida (Fig. 4.6) Antes de realizar la extracción, las plantas se mantuvieron en la oscuridad por 20 horas (línea 1), o 20 horas seguidas de 1 hora de luz (línea 2) o 3 horas de luz (línea 3). En la parte A se muestra el resultado de una tinción con coomasie blue y en la B la autorradiografía.
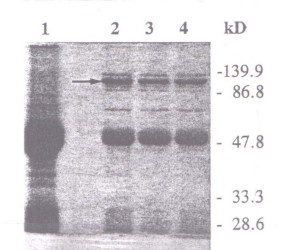
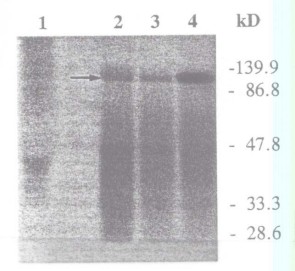
Figura 4.6
L1 F=L2P; L2F=L3P; L3F=L4P
Se sabía que bajo ciertas condiciones la PEPC es inhibida por el malato. Para intentar correlacionar los resultados de los estudios de modificación covalente con este fenómeno, se realizó el experimento de la Tabla 4.4 donde se midió la sensibilidad al malato como la cantidad de malato requerida para producir una inhibición del 50% (I50) de preparaciones de PEPC provenientes de plantas mantenidas en la luz (light) o en la oscuridad (dark) seguida (followed) de luz, con o sin remoción del floema (girdling). Los ensayos se realizaron a pH 7,3 (normal) o a pH 8,0 (donde no se observa actividad inhibitoria del malato).
Tabla 4.4
|
Pretreatment of Plants |
I50 (L-malate)a |
Activityb |
|
|
PH 7.3 |
PH 8.0 |
||
|
mM |
Units/mg of protein |
||
|
Light (control) |
0.121 |
0.23 |
0.36 |
|
Dark (26h) |
0.074 |
0.18 |
0.38 |
|
Dark plus girdling (26h) |
0.059 |
0.16 |
0.37 |
|
Dark (26h) followed by light (3h) |
0.124 |
0.24 |
0.37 |
|
Dark (26h) followed by light plus girdling (3h) |
0.06 |
0.15 |
0.38 |
a
Assayed under suboptimal conditions (pH 7.3, 0.5mM PEP) in the presence of 0.01 to 0.25mM L-malate (pH ajusted).b
PEPC activity was assayed under suboptimal (pH 7.3, 0.5mM PEP) or optimal (pH 8.0, 2.5mM PEP) conditions. Results are the means of three separate experiments (se = ± 5 – 10%)a) Explique el resultado de la Fig 4.6 y calcule el peso molecular de la PEPC.
b) ¿Cómo se correlaciona este resultado con el de la Tabla 4.4?. ¿En qué etapas de la modificación covalente juega un papel la parte aérea de la planta?.
c) ¿Cuál es la razón fisiológica de que el malato inhiba a la PEPC en las condiciones observadas?
Plant Physiol. 108: 1561-68 (1995)
8) La glutamino sintetasa (GS) cataliza la formación de glutamina a partir de glutamato y NH3:
glutamato + NH3 + ATP ® glutamina + ADP + Pi
Dado que el glutamato y la glutamina son los aminoácidos precursores de la biosíntesis de todos los demás compuestos nitrogenados, esta enzima juega un rol central en la asimilación del nitrógeno. Así, la regulación de la GS es muy compleja. Esta enzima es un dodecámero, cada una de cuyas subunidades puede adenilarse por la acción de una adenil transferasa específica. El estado de adenilación (número de subunidades adeniladas) modifica la actividad de la GS. Para la reacción de biosíntesis de glutamina, la actividad decrece a medida que se adenila, siendo la glutamina un factor que modifica la adenilación a través de una acción sobre la adenil transferasa.
La GS cataliza, además, la reacción g glutamil transferasa (g GT):
glutamina + hidroxilamina ® g glutamil hidroxamato + NH3
Esta reacción se utiliza para estudiar el estado de adenilación, ya que a diferencia de la reacción biosintética, ambas formas son activas, aunque con diferente pH óptimo. Además, el MgCl2 es un inhibidor de la actividad de las subunidades adeniladas, con lo cual la incubación en presencia o ausencia de Mg2+ indica el estado de adenilación.
Se estudió la GS de Klebsiella aerogenes a través de la reacción g GT a partir de cultivos que habían sido incubados en medios limitados en NH3 adicionados o no con glutamina. Los extractos de estos cultivos se incubaron en un rango de pH durante 30 minutos, luego de los cuales se realizó la reacción g GT (Fig. 4.7, círculos llenos: medio con glutamina; círculos vacíos: medio sin glutamina). En otro experimento, a un extracto de células crecidas en medio sin glutamina se le adicionó este aminoácido, y a diferentes tiempos luego de la adición se tomó muestras con las que se llevó a cabo la reacción g GT a pH=7,15 en presencia o ausencia de Mg2+ (Fig 4.8). Finalmente, y para comprobar que las modificaciones observadas se deben a la adenlación, se incubó GS adenilada por distintos tiempos con fosfodiesterasa de veneno de serpiente (que cataliza la remoción de adenilos) en presencia o ausencia de Mg2+ luego de los cuales se midió la actividad g GT a pH=7,5 (Fig. 4.9).
a) ¿En qué consiste la adenilación y cómo esta puede modificar el comportamiento cinético de la GS?.
b) ¿Cómo se hizo el experimento de la Fig 4.7?. ¿Qué información obtiene de él?
c) ¿Qué efecto tiene la glutamina (inhibitorio o estimulatorio) sobre la actividad de la adenil transferasa?. ¿Es esta respuesta 100% segura o requiere de más experimentos?. En ese caso, ¿cuáles?.
d) De acuerdo a su respuesta c), ¿Cuál es la ventaja fisiológica de la modificación producida por la glutamina sobre la actividad de la GS?
J. Bacteriol. 129: 1001-9 (1977)
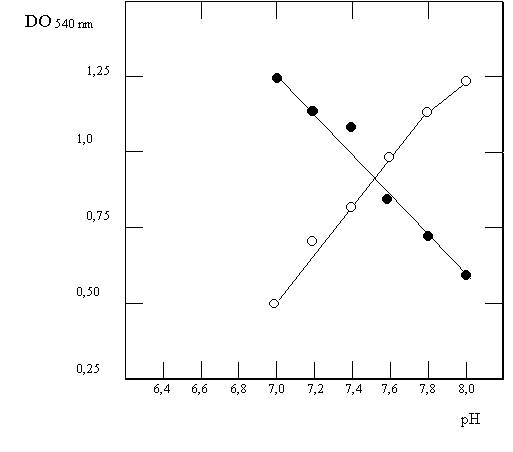
Figura 4.7
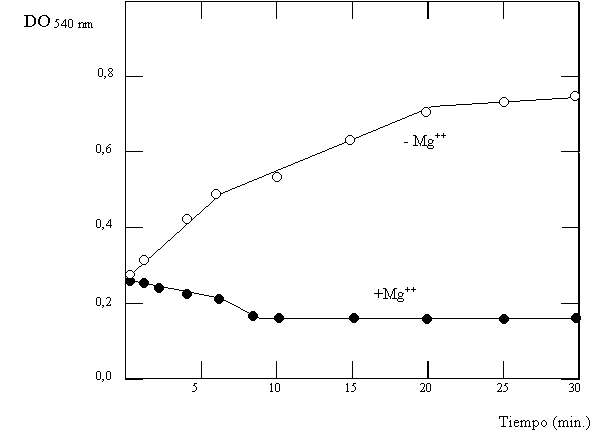
Figura 4.8
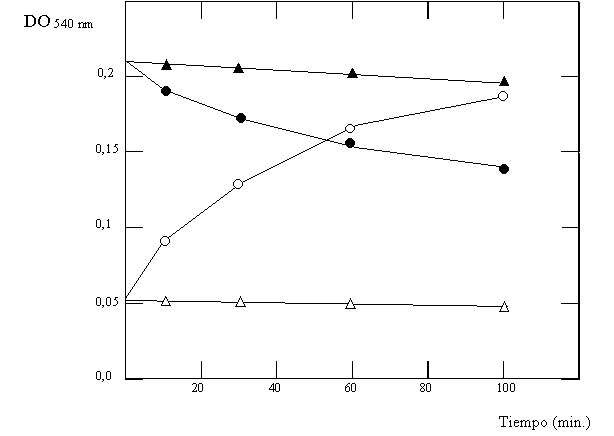
Figura 4.9
![]()
Seminarios
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
![]()
Home Docentes Cronograma Programa Materia Programa TP Seminarios Links Publicaciones Bibliografia