
|
SEMINARIO 10 |
CINETICA ENZIMATICA I
Conceptos Básicos y Modelo de Michaelis-Menten
1) Se realizó un estudio exhaustivo de una cepa de Saccharomyces cereviceae (D273-10B) y resultó de interés analizar el comportamiento cinético de la enzima directamente comprometida en una mutación, la porfobilinogenasa (PBGasa), la cual presenta una cinética michaeliana y no se detecta efectos cooperativos entre las dos proteínas que forman el complejo de la PBGasa (deamidasa e isomerasa).
Para todos los estudios cinéticos se utilizaron las mismas cantidades de células (absorbancia medida a 540 nm). La mezcla de reacción se llevó a cabo en condiciones óptimas de temperatura y pH. En cada caso la enzima se inactivó tomando 1 ml de mezcla de reacción (que contenía una dilución 1:1000 del extracto enzimático) y agregando 4 ml de reactivo B4C (inactivador coloreado), posteriormente, se leyó la absorbancia a 500 nm.
El primer experimento se refleja en la Tabla 3.1, donde se entregan resultados de concentración de producto expresados en el tubo de colorimetría, para diferentes concentraciones de sustrato.
Tabla 3.1
Se realizó un segundo experimento: un estudio cinético en diferentes fases de crecimiento celular, encontrándose los resultados expresados en la Tabla 3.2:
Tabla 3.2
| Fase de crecimiento | |||
| 1 | 2 | 3 | |
| Vmax (umoles min-1) | 180 | 300 | 350 |
| Km (mM) | 15.1 | 15.3 | 15.2 |
En un tercer y último experimento se seleccionaron las células que se encontraban en la fase 2 de crecimiento y se realizó una cinética en la cual se agregó a la mezcla de reacción ácido 5-aminolevúlico obteniéndose valores de Vmax=300 m moles min
-1 y Km=5,1 mM.a) Determine los parámetros cinéticos utilizando los datos del primer experimento.
b) Interprete los resultados del segundo experimento y si es posible, hágalo numéricamente.
c)¿Qué conclusiones puede obtener del tercer experimento?
2) Se realizó un estudio cinético de una enzima proteolítica, cuya actividad se siguió utilizando como sustrato un compuesto sintético, del tipo CBZ-p-nitro fenil éster, que al ser hidrolizado libera un producto coloreado (p-nitrofenol o p-nitrofenolato) que absorbe a una longitud de onda de 400nm. Se obtuvieron los resultados que se muestran en la Tabla 3.3:
Tabla 3.3
|
Sustrato (M) |
velocidad inicial (m mol/min) |
|
5 x 10 -2 |
0.25 |
|
5 x 10 -3 |
0.25 |
|
5 x 10 -4 |
0.25 |
|
5 x10 -5 |
0.20 |
|
5 x10 -6 |
0.071 |
|
5 x10 -7 |
0.0096 |
a.- ¿Cuál es la Vmáx de la reacción? No utilice gráficos, responda analíticamente.
b.- ¿Cuál es el Km de la enzima? No utilice gráficos, responda analíticamente.
c.- Demuestre si esta reacción sigue la cinética de Michaelis Menten.
d.- ¿Cuál es la velocidad utilizando una concentración de sustrato de 1 x 10
-6 M y 1 x10-10 M?e.- Calcular la cantidad de producto formado, durante los primeros cinco minutos de reacción a S = 2 x10
-3 Mf.- Suponga que la concentración de enzima aumenta en un factor de 4. ¿Cuál es el valor de Km? ¿De Vmáx?. En estas condiciones, ¿qué valor de vo se obtiene si S = 5 x10
-6 M?.g.- Defina por lo menos dos unidades de Actividad Enzimática.
Para la determinación de la cantidad de proteína presente en el extracto enzimático, se utilizó el método de Lowry, para lo cual se confeccionó una curva de calibración utilizando un patrón de albúmina 4 mg/ml, el cual se diluyó 1/10. Los dosajes se realizaron con los volumenes indicados de la dilución en un volumen final de 1,3 ml (Tabla 3.4)
Tabla 3.4
|
Tubo |
1 |
1’ |
2 |
2’ |
3 |
3’ |
4 |
4’ |
|
Alb (ml) |
0 |
0 |
0,1 |
0,1 |
0,15 |
0,15 |
0,20 |
0,20 |
|
DO670nm |
0,014 |
0,016 |
0,08 |
0,088 |
0,116 |
0,100 |
0,156 |
0,140 |
h.- El valor de DO670nm de la determinación de una alícuota de 0,2 ml del extracto enzimático en estudio fue 0,095. Calcule, si es posible, la concentración de proteína en mg/ml. Calcule la actividad específica del extracto.
3) Para estudiar la relación entre la estructura de los tRNA y la especificidad con que se incorporan los aminoácidos en las proteínas, se sintetizó artificialmente dos tRNA de levadura con la secuencia del tRNA
Asp, sin modificaciones en sus bases. Uno de ellos llevaba la secuencia no-modificada del tipo salvaje (U1-A72) y el otro llevaba guanina (G) en su extremo 5' y citosina en su extremo 3' (G1-C72). La secuencia nucleotídica de estos tRNA se muestra en la Figura 3.1.Con cada tRNA se midió la aminoacilación in vitro por aspartil-tRNA sintetasa (con [
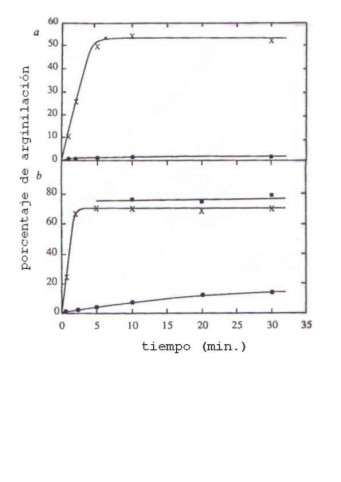
3H]aspartato como sustrato) o por arginil-tRNA sintetasa (con [3H]arginina como sustrato), evaluándose la incorporación de marca en el tRNA. En la Tabla 3.5 se muestran los resultados del análisis cinético de cada enzima con cada tRNA, y en la Figura 3.2 la actividad de arginilación (como porcentaje frente a la aspartilación) del transcripto G1-C72 (X), del U1-A72 (n ) y del tRNAAsp modificado (nativo) (l ) en presencia de un extracto enzimático crudo de levadura (a) o de arginil tRNA sintetasa purificada (b).Tabla 3.5. Aspartilación y arginilación del tRNA
Asp de levadura nativo o no modificado (variantes U1-A72 o G1-C72, ver Fig. 4.11) por dos aminoacil-tRNA sintetasas.| tRNA | Km (nM) | |
| Aspartil-tRNA-sintetasa | Arginil-tRNA-sintetasa | |
| tRNAAsp nativo | 44 | 8600 |
| tRNAAsp G1-C72 | 33 | 390 |
| tRNAAsp U1-A72 | 28 | 110 |
| tRNAArg/UCU | - | 7,3 |
a) ¿Cómo cree que se obtuvo los tRNA no modificados?
b) Señale las regiones más importantes en el esquema de la Figura 3.1 en relación a la función de los tRNA.
c) ¿Cuál fue el efecto de suprimir las modificaciones en las bases del tRNA
Asp?d) Proponga una hipótesis que explique estos resultados.
Figura 3.1
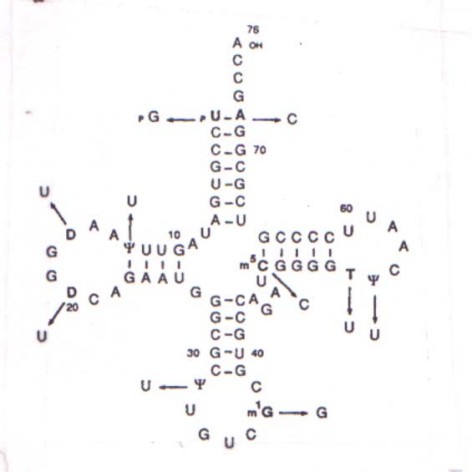
Figura 3.2
4) La arginina sintetasa (AS) cataliza la reacción:
Arginilsuccínico « Arginina + Ac. Fumárico
AS
En un experimento se midió la cantidad de arginina producida en función del tiempo, a diferentes concentraciones iniciales de sustrato. En todas las mezclas de incubación las condiciones de reacción fueron las mismas, y en cada mezcla de incubación se adicionó 5 m l de una preparación de enzima. El volumen final de cada mezcla de reacción fue de 10 ml. Los resultados obtenidos se muestran en la Tabla 3.6

a) ¿Por qué las velocidades que se determinan son valores iniciales?
b) Esquematice brevemente un protocolo que permita obtener los valores de v
o de la tabla (en forma cualitativa)c) Sabiendo que una unidad enzimática se define como la cantidad de enzima que cataliza la producción de 1 m mol de Arg/10 min, en las condiciones de reacción del ensayo, y a concentración de sustrato saturante, ¿cuál es la actividad de la preparación de enzima, en UE/ml?
d) ¿Qué datos requeriría para calcular la actividad específica de la preparación de enzima?. ¿Qué datos para calcular el número de recambio?
e) Calcule el Km. ¿Cuál es el significado físico del parámetro Km?. ¿Considera posible disminuír el Km para este par enzima-sustrato?.
5) Se determinaron las constantes cinéticas de la b -galactosidasa para la lactosa, siendo:: Km=20 mM y No de recambio=400 s
-1. Otra enzima capaz de hidrolizar la lactosa, la b -glicosidasa, mostró los siguientes valores para lactosa: Km=5,1 y No de recambio=50 s-1.a) Para una mezcla equimolar de ambas enzimas, y en condiciones óptimas de ensayo, ¿cuál es la relación de velocidades de reacción?. ¿Cuál es el máximo valor de esa relación y cuál el mínimo?.
b) Cómo determinaría el Km de la b -galactosidasa para otro sustrato, el PPNP-gal, incluyendo concentraciones de sustrato, volúmenes y tiempos, sabiendo que:
i) De acuerdo a las referencias bibliográficas de otras b -galactosidasas, es posible que su enziima tenga una afinidad por el PNP-gal alrededor de 20 veces superior a la afinidad por lactosa.
ii) Una unidad enzimática es la cantidad de enzima que cataliza la producción de 1 m mol de 4-nitrofenol/min, en condiciones óptimas de ensayo (pH=7,0; temperatura=37oC).
iii) La detección del producto se realiza volcando la muestra sobre un volumen 2,5 veces inferior de Na
2CO3 1 M, y determinando la absorbancia a 420 nm. (Rango de validez de la Ley de Lambert-Beer: 0,02-0,30 m moles de 4-nitrofenol/ml final).iv) Ud. dispone de una solución stock de buffer (con los iones requeridos) veinte veces concentrada respecto a la necesaria para el ensayo. Dispone también de una solución stock de PNP-gal 50 mM.
6) Se estudió la acción de la lactato deshidrogenasa (LDH) obtenida a partir de hígado y de corazón. Esta enzima es capaz de oxidar al L-lactato mediante una reacción acoplada a la reducción del NAD+, produciendo piruvato y NADH.
Se colocaron 0,5 ml del extracto enzimático, 0,3 ml de la solución de NAD+ (10 mM), 0,2 ml de una solución de hemicarbazida (30 mM) y 1 ml de buffer fosfatos pH=7,6. A cada mezcla de reacción se le adicionó agua y finalmente, una solución de L-lactato 100 mM, en las cantidades indicadas en la Tabla 3.7. El volumen total de cada mezcla fue de 3 ml. Inmediatamente después de la adición de lactato se mezcló y se siguió la reacción a través de la medida de absorbancia a 340 nm. El NAD+ tiene una absorción a 340 nm pequeñísima, y su forma reducida tiene un e
340=6,220.10-3 M-1.cm-1. Las reacciones catalizadas por deshidrogenasas en la dirección de reducción del NAD+ a pH neutro son termodinámicamente desfavorables, pero puede conseguirse un buen desplazamiento con reactivos que atrapen a los aldehídos o cetonas producidos.Tabla 3.7. Actividades de la lactato deshidrogenasa de hígado y de corazón.
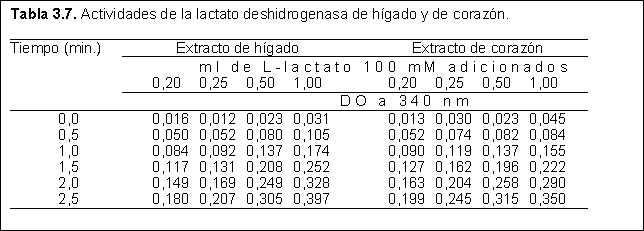
a) ¿Cuál es la actividad de cada extracto?. Utilice al menos dos unidades de actividad diferentes.
b) ¿Cuál es la afinidad de cada LDH por el lactato?
c) ¿Cuál es la cantidad de cada enzima en cada extracto?
d) Tal como ha caracterizado ambas LDH, ¿cree que dichas características son adecuadas a sus respectivas funciones?
e) Explique mediante el uso de fórmulas y ecuaciones cómo es posible llevar a cabo las medidas de NADH.
7) Para purificar y caracterizar invertasa de levadura, Ud. obtuvo un extracto crudo.
En la deteminación de la actividad de invertasa en el mismo utilizó la mezcla de reacción de la Tabla 3.8
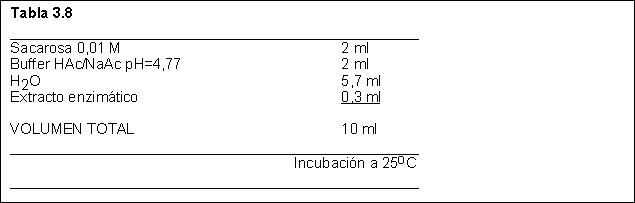
Tomó muestras de esta mezcla de incubación a los siguientes tiempos:
1 minuto 5 minutos 15 minutos
Desproteinizó estas muestras llevando a un volumen total de 5 ml. Las filtró y del filtrado tomó muestras de 0,8 ml para efectuar la colorimetría de azúcares reductores.
Obtuvo las siguientes lecturas de DO a 640 nm:
TUBO 1 min. 5 min. 15 min.
DO - Blanco 0,084 0,388 0,470
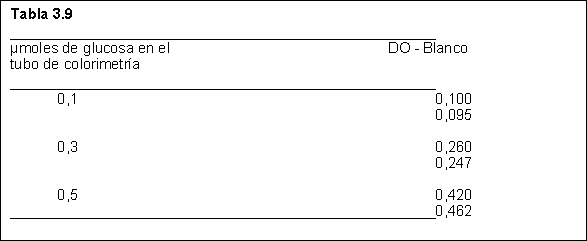
Paralelamente efectuó una curva de calibración, obteniendo los resultados de la Tabla 3.9
Calcule la actividad de la invertasa, expresada en número de unidades de invertasa por ml y en catales por ml ambas referidas al extracto enzimático.
Unidad de invertasa: cantidad de enzima que en las condiciones óptimas de reacción hidroliza 10 m moles de sacarosa por minuto.
8) La bacteria Gram negativa Haemophilus influenzae, utiliza la fructosa como principal fuente de carbono y energía. La fructosa es transportada desde el medio por una proteína transportadora específica (PTS) que libera fructosa-6-fosfato en el interior celular. A partir de la fructosa-6-fosfato H. influenzae obtiene energía mediante fermentación. En ausencia de fructosa en el medio de cultivo, H. influenzae fermenta otros hidratos de carbono, entre ellos glicerol, el cual es incorporado del medio al interior celular por una proteína transportadora específica codificada por el gen ptg (proteína transportadora de glicerol), que libera glicerol-3-fosfato.
Para estudiar a la proteína transportadora de glicerol (PTG), 2 litros de un medio mínimo, conteniendo glicerol como única fuente de carbono, fueron inoculados con 20 ml de un cultivo de H. influenzae en crecimiento exponencial y cultivados 48 hs a 37ºC. Cuando la DO600nm del cultivo fue de 0,8 (fase logarítmica tardía), las bacterias fueron centrifugadas y resuspendidas en 100 ml de buffer fosfato pH=7. Se agregó lisozima y se incubó 2 horas a 37ºC. Se centrifugó y con el sobrenadante (extracto crudo) se inició la purificación. La purificación se resume en la Tabla 3.10 (lamentablemente se han perdido algunos datos, pero se conoce el protocolo realizado). Para la determinación de proteínas luego de hacer el intercambio iónico, se tomaron 0,3 ml y se los llevó a un volumen final de 5 ml, luego se realizó una dilución 1:500 y finalmente se midió proteínas por el método de Bradford usando 1 ml de muestra para la reacción colorimétrica. Esta determinación se realizó por duplicado obteniéndose 0,53 y 0,55 como valores de DO590nm. La curva de calibración para el método de Bradford se realizó como se detalla en la Tabla 3.11. Para la determinación de actividad luego de la exclusión molecular, se tomaron 0,5 ml y se los llevó a un volumen final de 5 ml, luego se realizó una dilución 1:15 y de allí se tomó 0,1 ml, al que se le agregó 0,9 ml de glicerol 2 mM y un exceso del dador de grupos fosfato en buffer fosfato 5mM pH 7,5 y se incubó a 37ºC. De esta mezcla de reacción se midió producto a diferentes tiempos, obteniéndose una velocidad inicial de 0,16 m mol de glicerol-3-fosfato/ml minuto. El Km bibliográfico de esta enzima para el glicerol es de 5 mM.
a) Complete los datos faltantes de la Tabla 3.10. Indique cómo realiza las diluciones indicadas en el texto para realizar las determinaciones de proteínas y la medida de actividad. Para realizar las diluciones usted dispone del siguiente material: pipetas 2-20 m l, 20-200 m l, 200-1000m l, 1-5 ml y 1-10 ml, tubos eppendorf de 1,5 ml de volumen máximo y matraces aforados de 10 ml y 25 ml.
b) Calcule rendimiento, recuperación y cuántas veces se purificó la proteína PTG.
Tabla 3.10. Resumen de la purificación de la proteína PTG.
|
Fracción |
Volumen (ml) |
Proteínas (mg/ml) |
Actividad (UE/ml) * |
|
Extracto Crudo |
100 |
50,0 |
47,0 |
|
Precipitación con (NH4)2SO4 al 60% |
10 |
67,0 |
470,0 |
|
Diálisis |
12 |
55,8 |
387,5 |
|
Q-Sepharosa (intercambio iónico) |
3 |
1500,0 |
|
|
Sephadex 200HR (exclusión molecular) # |
5 |
50,0 |
*
Unidad Enzimática: Cantidad de enzima que cataliza la formación de 1 m mol de glicerol-3-fosfato/ml minuto, a 37ºC en buffer fosfato 5mM pH 7,5# En la cromatografía de exclusión molecular la proteína PGT salió en un volumen de elución correspondiente a 200 KDa
Tabla 3.11. Curva de calibración del método de Bradford.
|
m g de albúmina |
0 |
5 |
10 |
15 |
20 |
|
DO590nm |
0,006 |
0,160 |
0,342 |
0,520 |
0,665 |
|
DO590nm |
0,009 |
0,189 |
0,342 |
0,495 |
0,680 |
![]()
Seminarios
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
![]()
Home Docentes Cronograma Programa Materia Programa TP Seminarios Links Publicaciones Bibliografia