LA FOSSILISATION DES OEUFS DE DINOSAURES
LA CONDUCTANCE DES COQUILLES D'OEUFS
L'ATTRIBUTION TAXONOMIQUE DES OEUFS
LES OEUFS ET L'EXTINCTION DES DINOSAURES
LES AUTRES GISEMENTS EUROPEENS
FOSSILES ET FOSSILISATION GENERALITES

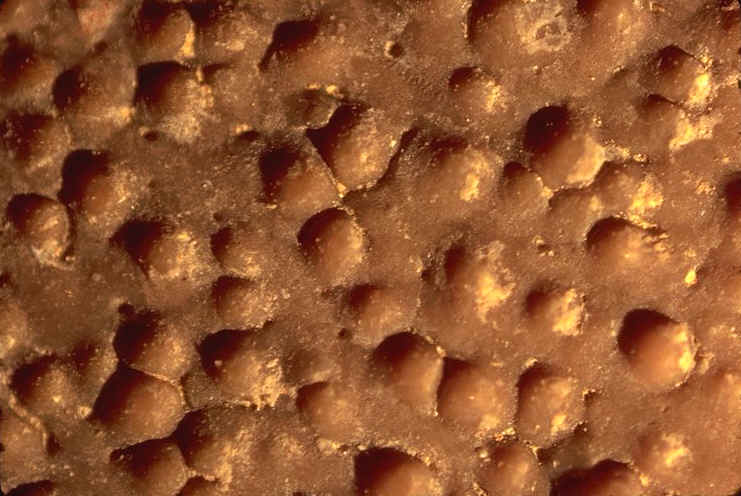
Cairanoolithus dughii ( Rognacien inf�rieur , Maastrichtien sup�rieur du gisement de La Cairanne / La Colline des Chapeliers , commune de Rousset-sur-Arc , Bouches-du-Rh�ne , France ) . Surface externe d'une coquille attribu�e � un Dinosaure . Des nodes ( petits nodules arrondis ) pars�ment la surface de la coquille ( le diam�tre de nodes varie de 0, 5 � 1 mm ) .On distingue clairement de petits crat�res de forme sub-circulaires . Ce sont des pores qui permettaient les �changes gazeux avec le milieu ext�rieur , n�cessaires � la survie et au d�veloppement de l'embryon.
Les oeufs fossiles r�colt�s dans la plupart des gisements ne contiennent pas de restes osseux et il est tr�s al�atoire d'attribuer un morphotype de coquille � telle ou telle cat�gorie de Dinosaure.C'est la raison pour laquelle une parataxonomie a �t� r�cemment �tablie. Quelques d�couvertes spectaculaires ont permis de d�couvrir en association des oeufs et des nouveaux-n�s ( Horner et Makela , 1979 ; Currie et Horner , 1988 ), des oeufs renfermant des embryons ( Horner et Weishampel , 1988 ; Norell et al. , 1988 ) et m�me un Dinosaure en train de prot�ger et de couver ses oeufs ( Norell et al. , 1995 ). En Europe un gisement d�couvert dans le bassin de Hateg en Roumanie ( Grigorescu et al. , 1994 ) a livr� un oeuf non �clos dont l'examen tomographique laisse suppose la pr�sence d'un embryon . La d�couverte la plus int�r�ssante est survenue en 1993 au Portugal o� une pal�ontologiste amateur Isabel Mateus d�couvrit sur le sol de la localit� de Paimogo , non loin de Lourinha , � 60 kilom�tres au nord-ouest de Lisbonne , de nombreux fragments de coquilles dans un niveau du Kimm�ridgien - Tithonique inf�rieur ( Jurassique sup�rieur , 140 millions d'ann�es ) . Des fouilles pratiqu�es en 1995 - 1996 permirent la mise au jour de nombreuses pontes tr�s compl�tes comptant jusqu'� 34 oeufs , ainsi que de vestiges d'embryons attribu�s � des Dinosaures Th�ropodes ( Megalosaurus sp. ? ) ( Mateus et al. , 1997 ) .
Dans le Midi m�diterran�en , ainsi d'ailleurs que dans le Cr�tac� sup�rieur de l'ensemble de l'Europe ( � la seule exception assez discutable de la Roumanie ( cf. supra ) ) , aucun reste d'embryons ou de juv�niles, int�gr�s ou associ�s � des oeufs fossiles, n'a jamais �t� signal� , en d�pit du nombre important d'oeufs entiers et de pontes compl�tes. De ce fait toutes les hypoth�ses formul�es � ce jour , y compris celles dont nous faisons �tat ici , restent hautement sp�culatives .
Dans la litt�rature pal�ontologique relative aux oeufs fossiles du Sud de la France les essais d'attribution taxonomique sont fr�quents ( Matheron , 1869 ; Gervais , 1877 ; Lapparent , 1947 et 1957 ;Dughi et Sirugue , 1976 ;Voss-Foucart , 1968 ; Kurten , 1968 ; Case , 1978 ; Erben et alii , 1979 ; Ginsburg , 1980 ; Seymour , 1980 ; Kerourio , 1982 ; Penner , 1983 ; Williams , Seymour et Kerourio , 1984 ) mais aucune des attributions taxonomiques �tablie au niveau g�n�rique, comme au niveau sp�cifique , ne peut �tre retenue en l'absence de preuves satisfaisantes .La quasi-totalit� des hypoth�ses repose sur de pures sp�culations , rarement et / ou vaguement �tay�es de comparaisons avec des ootypes retrouv�s en d'autres points du Monde et parfois associ�s � des embryons.
La microstructure de la plupart des oeufs fossiles du bassin d'Aix diff�re de celle des oeufs de crocodiliens et de ch�loniens fossiles et actuels .Les oeufs de ch�loniens sont compos�s d'unit�s de base sph�rolithiques avec des cristaux d'aragonite �troitement agglom�r�s et en forme d'aiguilles ( " needle-like " , Hirsch , 1983 ) .M�me sous leur forme calcitique diag�n�tique , ils n'offrent aucune ressemblance avec les sp�cimens de coquilles d'oeufs fossiles retrouv�s dans la r�gion d'Aix-en-Provence ( Young , 1950 ; Hirsch , 1983 ).Quant aux oeufs de crocodiliens , ils sont compos�s d'unit�s de base claviformes visibles en lumi�re naturelle et se d�veloppant en s'�largissant vers la surface externe de la coquille ( Ferguson , 1982 ; 1985 ; Hirsch , 1985 ).Des fragments de coquilles d'oeufs de crocodiliens ont �t� d�couvertes par l'auteur en diff�rents niveaux du Maastrichtien continental du bassin d'Aix ( Kerourio , 1987 ). Leur microstructure est ais�ment reconnaissable et elle diff�re radicalement de celle des autres types d'oeufs fossiles retrouv�s dans le m�me secteur et d�crits ici .
Une ressemblance morphologique et microstrucuturale peut �tre �tablie entre des oeufs aviens fossiles et actuels et certains des ootypes fossiles retrouv�s dans la r�gion d'Aix-en-Provence.Dans l'oeuf avien la coquille est organis�e en deux ou trois couches morphologiques horizontales : une couche form�e par la juxtaposition des unit�s mamillaires suivie d'une ou de deux couches sup�rieures ( ou prismatiques ) d�pourvues , en lumi�re naturelle , d'unit�s de croissance distinctes ( Terepka , 1963 ; Tyler , 1969 ; Simons , 1971 ; Pooley , 1979 ).Cette premi�re caract�ristique distingue clairement l'oeuf avien des oeufs du oogenre Prismatoolithus et apparent�s .Chez ces derniers les unit�s de croissance de forme columnaire sont visibles en lumi�re naturelle dans toute l'�paisseur de la coquille ( Kerourio , 1981 ).Par ailleurs nous sommes enclins � consid�rer comme un trait sp�cifiquement reptilien la pr�sence de petits nodes isol�s et bien individualis�s � la surface externe de la coquille. La surface des oeufs aviens actuels est g�n�ralement d�pourvue d'une ornementation analogue .Certains sp�cimens fossiles ( oeufs attribu�s � des ratites ) peuvent pr�senter une surface externe comportant des vallonnements et des cr�tes ( Dughi et Sirugue , 1962 ; Touraine , 1960 ; Sauer , 1966 et 1972 ).
L'hypoth�se d'une appartenance avienne de certains types d'oeufs fossiles du bassin d'Aix ne peut �tre totalement �cart�e.Elle est quasi-certaine pour un nouveau ootype d'affinit�s clairement ratites et que nous avons d�couverts en diff�rents points du Maastrichtien sup�rieur et de la base du Montien ( niveaux sous-jacents et sus-jacents � la formation dite " Poudingue de La Galante )et qui pourrait �tre attribu� � Gargantuavis philoinos, un grand oiseau apt�re de la taille d'une autruche actuelle, attest� dans le Maastrichtien de l'Aude ( Buffetaut et Le Loeuff , 1998 ).L'appartenance avienne est aussi probable pour d'autres ootypes non encore d�crits que nous figurons ici. La pr�sence d'une avifaune diversifi�e est par ailleurs confirm�e par la d�couverte r�cente de restes osseux dans diff�rents gisements du Campanien et du Maastrichtien du Midi m�diterran�en ( gisement de Cruzy dans l'H�rault , gisement de Campagne-sur-Aude / Esp�raza dans l'Aude , gisement de Fox-Amphoux dans le Var ( Buffetaut , 1998 ; Buffetaut et Le Loeuff , 1998 ; Buffetaut et alii , 1995 )).
Le rattachement des oeufs fossiles du bassin d'Aix-en-Provence , et de mani�re plus g�n�rale de l'ensemble du Midi m�diterran�en fran�ais , � des esp�ces dinosauriennes clairement identifi�es se heurte � l'absence g�n�ralis�e de restes d'embryons ou de formes juv�niles directement associ�es aux aires de pontes . Seules des comparaisons morphologiques et microstructurales effectu�es avec des oeufs fossiles provenant d'autres r�gions du Monde et associ�s � des vestiges embryonnaires peuvent nous �clairer sur l'appartenance taxonomique des oeufs fossiles du Cr�tac� Sup�rieur proven�al. C'est ainsi que l'oogenre Megaloolithus se retrouve dans le Cr�tac� sup�rieur de la Patagonie argentine ( Gisement de Auca Mahuevo , provincia de Neuquen , Argentine ) . Les oeufs de forme sph�rique ( 13 � 15 centim�tres de diam�tre ) pr�sentent d'�troites similitudes morphom�triques , ornementales et microstructurales avec Megalolithus mamillare du Rognacien sup�rieur du bassin d'Aix-en-Provence. Certains des oeufs fossiles retrouv�s dans le gisement d'Auca Mahuevo contenaient des restes d'embryons attribu�s � des dinosauriens sauropodes du genre Titanosaurus ( Chiappe et alii , 1998.- Nature , vol. 396 , p. 258 - 261 ).Quant aux oeufs fossiles de la famille des Prismatoolithidae , fr�quents dans le Cr�tac� sup�rieur du Midi m�diterran�en , leur appartenance � des dinosauriens th�ropodes semble confirm�e par la d�couverte d'oeufs de ce type contenant des embryons dans le Kimm�ridgien -Tithonique ( Jurassique sup�rieur ) des environs de Lourinha au Portugal ( Mateus et al. , 1997 ).Une fouille m�thodique de ce gisement a r�v�l� plusieurs pontes compl�tes contenant des oeufs oblongs dot�s d'une coquille mince ( 0,7 � 1 mm ) � surface externe lisse. Plusieurs dizaines de petits os ont �t� r�colt�s � la proximit� , autour ou � la surface des oeufs fractur�s .Certaines caract�ristiques morphologiques de ces fragments osseux ( forme de la t�te f�morale , forme du plateau tibial , diaphyse creuse des f�murs et des tibias , etc ... ) sont caract�ristiques des Th�ropodes ( Mat�us et al. , 1997 , p. 75 ).
 Retour au Menu
Retour au Menu{kind=link}
![]() Cliquez ici pour revenir en d�but
de page
Cliquez ici pour revenir en d�but
de page