|
Link to research papers and articles: (http://wysinger.homestead.com/keita.html) |
|
Link to current African DNA research: (http://exploring-africa.blogspot.com/) |
|
Google Search- other data
Home page |
Home | Quotations | Misc Notes | Notes 2 | Hair | DemicDiff | Diversity | DNA| Asian IQ | Keita2008 data | Blood | Debunk3
Egypt
in Africa |
Black-Greek-DNA links | Notes
3 |Notes
4| Notes
5 | Notes
6 |
Notes 7 | Misc
news clips | Ethiopians
| Nubians
|
African Tmeline| Mesopotamia
| Egypt-
A Tropical Civ || KnolArchiv |
Notes8|
Notes
9 | Notes
10 | Notes
11
|
Link to research papers and articles: (http://wysinger.homestead.com/keita.html) |
|
Link to current African DNA research: (http://exploring-africa.blogspot.com/) |
|
Google Search- other data
Home page |
African
genetic diversity- Kittles and Keita
Rick Kitties1 and S. O. Y. Keita2
Interpreting African Genetic Diversity
African Archaeological Review, Vol. 16, No. 2, 1999
-----------------------------------------------------
Explanations of human biological variation in extant African populations have
historically been shaped by a racial paradigm. In this paradigm deep genetic differences
are assumed to exist between so-called "racial" groups or types, which are
viewed as being composed of nearly uniform individuals and are taken as "natural"
fundamental taxonomic units of Homo sapiens. These types were originally based
on specific aspects of external morphology. Carleton Coon (1962), a physical
anthropologist who contributed to this paradigm, proposed a theory of human
evolution which postulated the independent emergence of five primordial races or
subspecies from distinct, separate, Homo erectus ancestors. Two of these purported
primordial races originated in Africa. They were called Congoid ("Pygmies") and
Capoid (Khoisan or "Bushmen"). In Coon's conception other continental Africans,
which exhibit a broad range of biological diversity, were considered to be primarily
or only the result of various levels of admixture between these two groups with
each other or with immigrants deemed to be of European or Asian origin and generally
called "Caucasian." Although the causal aspects of Coon's theory have been
rejected, racial thinking still persists today using Coon's categories and is quite
evident in the methods and interpretation of genetic studies of African populations
(Keita and Kittles, 1997).
Here we briefly discuss the implications of modern genetic studies of African and
world populations for understanding African biohistory.
We propose an evolutionary biogeographical model as an alternative to
"racial" explanatory models of African biological diversity. A rational definition
of African in a biological sense should be derived from biogeography and not be
based on uninformed (and biased) traditions of practice.
Racial models, as traditionally presented, are static, and obligate one to postulate
gene flow as the primary explanation for variation. The racial paradigm
is manifested in the early writings on Africa by Seligman (1930) and Coon
(1962,1965) and is still operationalized in varying degrees by anthropologists and
geneticists, who otherwise reject Coon's thinking, to explain the vast biological
diversity observed in the continent. While it is a formidable task to evaluate the
extent of genetic diversity in Africa, it is quite evident that in many of these studies
there is the lack of an evolutionary perspective. This is especially the case when
northern Africa and the Horn are considered. The variation in these regions has
generally been explained as being due primarily to admixture between "Africans"
and "non-Africans." In fact, supra-Saharan African populations have frequently
been conceptualized as being derived from "European" ancestors and, hence, non-
African (see Seligman, 1930). The genetic profiles of supra-Saharan populations
are indeed in a relative sense "intermediate" to those of various sub-Saharan groups
(stereotypically defined) and "Eurasians" (see Guglielmino et al., 1987; Chibani
et al., 1985; Tartaglia et al., 1996; Merghoub et al., 1997). But is this genetic
"intermediateness" due primarily to supra-Saharan populations being foundationally
an admixed group (the result of gene flow between two distinct "races")? Or
is their biology reflective of factors acting on indigenous modern Homo sapien
populations in Africa?
Invoking admixture to explain human variation is related to another polemic
generally seen in many genetic studies of African populations. This is the persistence
of the socially constructed normative view of the African as only the "Forest
Negro" type, or the so-called "Pygmy" from Central Africa, in line with Coon's
(1962, 1965) thinking. These populations are used as the representative African
in many studies (see Bowcock et al., 1991, 1994; Cavalli-Sforza et al., 1994).
Northern African populations are rarely used in research under the designation
"African." Also, studies which utilize pooled samples of individuals from distinct
African populations as a representative "race" are problematic. If modern humans
evolved in Africa 150,000 to 200,000 years ago, and dispersed outside of Africa
only about 100,000 years ago, why should it be assumed that those populations
which remained in Africa became stagnant and homogeneous? While African populations
share a common ancestry, they also have evolved separately for a long time.
Evidence for isolation by distance operating in Africa is seen by genetic distinctions
among eastern, western, northern, central, and southern Africa (Excoffier
et al., 1987).
These genetic differences broadly correlate with geographic distances.
Various populations in Africa have interacted via migrations during past
history. One striking and most apparent signature of migration is the dramatic
eastern-to-western Africa cline of mitochondrial DNA (mtDNA) haplogroup L3a
frequencies (Watson et al., 1997). Haplogroup L3a is closely related to a mtDNA
haplotype common in European populations [the Cambridge Reference Sequence
(Anderson et al., 1981)]. A subgroup of related mtDNA haplotypes appears to be
East African specific and may represent a common ancestral sequence for most of
Europe and Eurasia. In other words, the mtDNA diversity observed in non-African
populations is a subset of African mtDNA variation. We note that while this group
of mtDNA haplotypes is common in Eastern Africa, it represents only a subset of
the total mtDNA diversity observed throughout the African continent. A similar
pattern is observed for nuclear (Tishkoff et al., 1996, 1998) and Y chromosome
(Passarino et al., 1998) variation in Eastern Africa. There are several implications
for these observations. First, it provides evidence for an African (specifically,
Eastern African) origin for Eurasians. Second it suggests that before major migrations
occurred out of the continent, populations were diverging. These observations
deconstruct racial thinking, especially the concept of "racial divergence."
The term racial divergence fails to describe the process responsible for producing
the variation that exists as a continuum in the human species. So-called
racial divergence dates reflect the times of differentiation of genes within populations
used in a given analysis and should be interpreted as such. "Racial divergence"
time estimates have been used to infer the age of the common ancestor between
sampled groups, but this is definitely not the time of origin for the so-called "racial
groups," which traditionally have been defined by morphology, nor is it the time
of "origin" for the sampled populations. Time, geography, and other data help elucidate
the larger meaning of genetic studies. A date of 156,000 years ago has been
suggested by Goldstein etal. (1995) for the separation of African (stereotypically
defined) and non-African populations. Given that there is no fossil evidence for
modern humans anywhere at 150,000 years ago except in Africa, this does not
represent an African/non-African "split." This date is actually an estimate of the
initial genetic divergence that occurred within the continent of Africa among our
modern human ancestors. After the expansion of modern humans out of Africa,
subsets of the resultant genetic variation were distributed throughout the other
continents. Most genetic variants observed outside of Africa are also found within
Africa at various frequencies (and are of indigenous origin); this clearly indicates
that "Africans" are not monomorphic. Northern African genetics, when this information
is considered carefully with palaeontological data, would not seem to be
explicable as simply "hybrids" or lost Eurasians. A different perspective, having
more explanatory power and consistent with the available data, is that northern and
Horn of Africa populations constitute gradients of differentiation largely reflective
of African biohistorical processes.
The data of Tishkoff et al. (1996, 1998) are best interpreted as suggesting
that a model of in situ evolutionary differentiation explains the bulk of variation in
Africa (which includes supra-Saharan, Saharan, and sub-Saharan regions). Markers
from mitochondrial DNA and the Y chromosome, which define maternal and
paternal lineages, respectively, also reveal significant differentiation of ancestral
African populations (Watson et al., 1996,1997; Passarino et al., 1998; Malaspina
et al., 1998). Another likely example of internal differentiation within Africa is to
be found in the "unusual" genetic makeup of the Khoisan, a southern African population
with a high frequency of purported "Asian" alleles (Cavalli-Sforza et al.,
1994). Some suggest admixture with Asians to explain the Khoisan genetic profile.
However, the Khoisan speakers may be descendants of a generalized ancestral human
population from which modern Asian and African populations were ultimately
derived. The Khoisan have been shown to be less diverse than other African populations
for a variety of genetic loci (Excoffier et al., 1987). This may be due to two
reasons: historical isolation and a smaller effective population size. The Khoisan
have historically remained relatively isolated from other African populations due
to cultural and linguistic differences. Also, the smaller effective population size is
likely the result of the hunter-gatherer culture of the Khoisan. These cultural and
subsistence patterns could promote significant differentiation of the population.
The overall pattern of genetic variation that exists within and outside of Africa
suggests an African origin of modern humans and a recent common origin of non-
African populations. The pattern of human genetic diversity supportive of this
position is observed in studies which examine genetic signatures left behind from
population expansions and population bottlenecks (Rogers and Harpending, 1992;
Harpending et al., 1993, 1998; Shriver et al., 1997; Reich and Goldstein, 1998;
Kimmel etal., 1998; Jorde et al., 1997). These studies consistently reveal an initial
population expansion within Africa prior to out-migration into other continents.
What is actually a population expansion has been mistakenly termed "racial divergence,"
which implies morphological differentiation into the recognizable entities
now labeled "races." In reality, it represents the early genetic divergence of ancestral
Homo sapiens. From the genetic data we find evidence of in situ differentiation
(or genetic divergence) of mid-Pleistocene populations in Africa and subsequent
migrations out of Africa into Europe and Asia, with continued drift due to isolation
by distance and founder effects, which abruptly end when expansion in population
size and frequent migrations occurred. Also, numerous environmental zones
exerted their own selective pressures on the populations. The populations remaining
in Africa are the primary ancestors of present Africans. This model is more
consistent with archaeological and molecular evidence and can be tested using various
data sets. This biogeographical perspective better explains the main source of
variation within Africa and obviates the need for racial thinking and its associated
baggage.
REFERENCES CITED
Anderson, S., Bankier, A., Barrell, B., de Bruijn, M,, Coulson, A., Drouin, J., Eperon, I., Nierlich, D.,
Roe, B., and Young, I. (1981). Sequence and organization of the human mitochondrial genome.
Nature 290: 457-465.
Bowcock, A., Kidd, J., Mountain, J., Herbert, J., and Cavalli-Sforza, L. L. (1991). Drift, admixture, and
selection in human evolution: A study with DNA polymorphisms. Proceedings of the National
Academy of Sciences USA 88: 839-843.
Bowcock, A..,etal.(\ 994). High resolution of human evolutionary trees with polymorphic microsatellites.
Nature 368:455^t57.
Cavalli-Sforza, L. L., Piazza, A., Menozzi, P., and Mountain J. (1988). Reconstruction of human evolution:
Bringing together genetic archaeological, and linguistic data. Proceedings of the National
Academy of Sciences USA 85: 6002-6006.
Cavalli-Sforza, L. L., Menozzi, P., and Piazza, A. (1994). The History and Geography of Human Genes,
Princeton University Press, Princeton, NJ.
Chibani, J., Lefranc, G., and Constans, J. (1985). Serum protein polymorphism among Tunisian Berbers:
Haptoglobin, transferrin and group-specific component subtypes, C3 and BF types. Annals of
Human Biology 5:449-462.
Clark, J. D. (1989). The Origin and Spread of Modern Humans: A broad perspective on the
African evidence. In Mellars, P., and Stringer, C. (eds.), The Human Revolution: Behavioral
and Biological Perspectives on the Origins of Modern Humans., Edinburgh University Press,
Edinburgh, pp. 565-588.
Coon, C. (1962). The Origin of Races, Alfred Knopf, New York.
Coon, C. (1965). The Living Races of Man, Alfred Knopf, New York.
Excoffier, L., Pellegrini, B., Sanchez-Mazas, A., Simon, C., and Langaney, A. (1987). Genetics and
history of sub-saharan Africa. Yearbook of Physical Anthropology 30: 151-194.
Goldstein, D., Ruiz-Linares, A., Cavalli-Sforza, L. L., and Feldman, M. (1995). Genetic absolute
dating based on microsatellites and the origin of modern humans. Proceedings of the National
Academy of Sciences USA 92: 6723-6727.
Guglielmino, CR., Menozzi, P., Piazza, A., and Cavalli-Sforza, L. L. (1987). Measures of genetic
admixture for North African populations. Atti Associazione Genetica Italiana 33: 177-178.
Harpending, H., Sherry, S., Rogers, A., and Stoneking, M. (1993). The genetic structure of ancient
human populations. Current Anthropology 34:483-496.
Harpending, H., Batzer, M., Gurven, M., Jorde L., Rogers A., and Sherry S. (1998). Genetic traces of
ancient demography. Proceedings of the National Academy of Sciences USA 95: 1961-1967.
Jorde, L., Rogers, A., Bamshad, M., Watkins, A., Krakowiak, P., Sung, S., Kere, J., and Harpending,
H. (1997). Microsatellite diversity and the demographic history of modern humans. Proceedings
of the National Academy of Sciences USA 94: 3100-3103.
Keita, S. O. Y., and Kittles, R. A. (1997). The persistence of racial thinking and the myth of racial
divergence. American Anthropologist 99(3): 534-544.
Kimmel, M., Chakraborty, R., King, J., Bamshad, M., Watkins, W., and Jorde, L. (1998). Signatures
of population expansion in microsatellite repeat data. Genetics 148: 1921-1930.
Malaspina, P., et al. (1998). Network analyses of Y chromosome types in Europe, Northern Africa,
and Western Asia reveal specific patterns of geographic distribution. American Journal of Human
Genetics 63: 847-860.
Merghoub, T., Sanchez-Mazas, A., Tamouza, R., Lu, C., Bouzid, K., Ardjoun, F., Labie, D.,
Lapoumeroulie, C., and Elion, J. (1997). Haemoglobin D-Ouled Rabah among the Mozabites: A
relevant variant to trace the origin of Berber-speaking populations. European Journal of Human
Genetics S: 390-396.
Nei, M., and Ohta, T. (1991). Evolutionary relationships of human populations at the molecular level.
In Osawa, S., and Honjo, T. (eds.). Evolution of Life, Springer, Tokyo, pp. 415-428.
Nei, M., and Roychoudhury, K. (1993). Evolutionary relationships of human populations on a global
scale. Molecular Biology and Evolution 10(5): 927-943.
Passarino, G., Semino, O., Quintana-Murci, L., Excoffier, L., Hammer, M., and Santachiara-
Benerecetti, S. (1998). Different genetic components in the Ethiopian population, identified by
mtDNA and Y-chromosome polymorphisms. American Journal of Human Genetics 62:420-434.
Reich, D., and Goldstein, D. (1998). Genetic evidence for a Paleolithic human population expansion
in Africa. USA. Proceedings of the National Academy of Sciences 95: 8119-8123.
Rogers, A., and Harpending, H. (1992). Population growth makes waves in the distribution of pairwise
genetic differences. Molecular Biology and Evolution 9(3): 552-569.
Seligman, C. (1930). The Races of Africa, Oxford University Press, London.
Shriver, M., Jin, L., Ferrell, R., and Deka, R. (1997). Microsatellite data support an early population
expansion in Africa. Genome Research 7: 586-591.
Tartaglia, M., Scano, G., and De Stefano, G. (1996). An anthropogenetic study on the Oromo and
Amhara. American Journal of Human Biology 8: 505-516.
Tishkoff, S., el al. (1996). Global patterns of linkage disequilibrium at the CD4 locus and modern
human origins. Science 271: 1380-1387.
Tishkoff, S., et al. (1998). A global haplotype analysis of the mytonic dystrophy locus: implications
for the evolution of modern humans and for the origin of mytonic dystrophy mutations. American
Journal of Human Genetics 62: 1389-1402.
Watson, E., Bauer, K., Aman, R., Weiss, G., von Haeseler, A., and Paabo, S. (1996). mtDNA sequence
diversity in Africa. American Journal of Human Genetics 59:437-444.
Watson, E., et al. (1997). Mitochondrial footprints of human expansions in Africa. American Journal
of Human Genetics 61:693-704.
Modern DNA analysis debunks simplistic "race" models
quote:
"It also does not mean that no differences whatsoever exist between continental groups. In fact, what Rosenberg et al. (2002) have shown is that given enough markers and the extraordinary power of Structure, the tiny amounts of genetic differences that exist between continents can also be discerned. However, this should not obscure the fact that on a worldwide scale, clines are a better representation of the human diversity than clades, and that continents do not represent more substantial discontinuities in such clines than many other geographical and cultural barriers. That clines are a more adequate representation of human genetic diversity than clades is not unexpected in view of earlier works that show that most genetic variation is found among individuals rather than among continents (e.g., Boyd 1950; Livingstone 1962; Lewontin 1972; Cavalli-Sforza et al. 1994).
In fact, also in the current data set, 87.6% percent of the total diversity is found among individuals and only 9.2% among continents
(Excoffier and Hamilton 2003), in agreement with many previous studies (e.g. Lewontin 1972; Owens and King 1999; but see also Edwards 2003). The current results are also not unexpected in view of the fact that identical DNA sequences of several kilobases are found on different continents
(Kaessmann et al. 1999; Gabriel et al. 2002). In fact, as much as a third of the entire human diversity of common haplotypes may be contained within single individuals
(Pääbo 2003). However, in spite of this, there is a great tendency in the literature to use a few populations from the extremes of continental landmasses (such as in Fig. 1A) to make worldwide inferences about substructures in the human gene pool. In fact, because human genetic diversity tends to be distributed
clinally, it is especially problematic to sample the extremes of continents because this will create the impression of sharp discontinuities in the distribution of genetic variants...
Clustering in the human gene pool is of practical importance. It has recently been claimed that the greatest genetic structure that exists in the human population occurs at the racial level
(Risch et al. 2002). Our results show that this is not the case, and we see no reason to assume that races represent any units of relevance for understanding human genetic history. In clinical practice, the classification of people into races, as recently suggested
(Risch et al. 2002; Burchard et al. 2003), could perhaps have some justification as a proxy for differences in environmental and other factors of relevance for public health or to help identify rare disease alleles
(Phimister 2003). However, in the absence of other knowledge, most alleles influencing susceptibility to disease or outcome of medical interventions cannot be expected to show significantly different frequencies between races.
An exception may be genes where different selection regimes have acted in different geographical regions. .. even in those cases, the genetic discontinuities seen are generally not racial or continental in nature but depend on historical and cultural factors that are more local in nature. For example, the hemoglobin S allele that causes resistance to malaria occurs not only in sub-Saharan Africa but also in southern Europe, the Middle East, and India
(Cavalli-Sforza et al. 1994). Lactose tolerance occurs both in Europe and in Africa
(Sahi 1994), and the deleted allele of CCR5 that confers resistance to human immunodeficiency virus occurs in Europe as well as in Asia (Martinson et al. 1997). Thus, even for a rapid and rough evaluation of genetic risk factors, racial background is of limited use, and direct analysis of the relevant gene is the only reliable way to evaluate genetic risk in an individual (Cooper et al. 2003). "
Source:
-- Evidence for Gradients of Human Genetic Diversity Within and Among Continents -- David Serre and Svante Pääbo Genome Research. (2004) 14: 1679-1685
mtDNA DATA in North Africa and Egypt

Important African mtDNA lineages, such as L3, which gave rise to mutations in Asia, were already undergoing expansion and mutation within Africa well PRIOR to the out of Africa. The rise of outside haplogroups such as M or N are a continuation of a process
already begun inside Africa.
"Past population size can be estimated from modern genetic diversity using coalescent theory. Estimates of ancestral human population dynamics in sub-Saharan Africa can tell us about the timing and nature of our first steps towards colonizing the globe. Here, we combine Bayesian coalescent inference with a dataset of 224 complete human mitochondrial DNA (mtDNA) sequences to estimate effective population size through time for each of the four major African mtDNA haplogroups (L0L3). We find evidence of three distinct demographic histories underlying the four
haplogroups. Haplogroups L0 and L1 both show slow, steady exponential growth from 156 to 213 kyr ago. By contrast, haplogroups L2 and L3 show evidence of substantial growth beginning 1220 and 6186 kyr ago, respectively. These later expansions may be associated with contemporaneous environmental and/or cultural changes. The timing of the L3 expansion812 kyr prior to the emergence of the first non-African mtDNA lineagestogether with high L3 diversity in eastern Africa, strongly supports the proposal that the human exodus from Africa and subsequent colonization of the globe was prefaced by a major expansion within Africa, perhaps driven by some form of cultural innovation."
--Atkinson, et al (2009). Bayesian coalescent inference of major human mitochondrial DNA haplogroup expansions in Africa. Proc Biol
Sci. 276(1655):367-73.
2007 DNA study finds that ancient mtDNA haplogroups M and N had an African origin, and expanded from a possible location in East Africa to areas within Africa and the rest of the world.
"Studies of human mitochondrial (mt) DNA genomes demonstrate that the root of the human phylogenetic tree occurs in Africa. Although 2 mtDNA lineages with an African origin
(haplogroups M and N) were the progenitors of all non-African haplogroups, macrohaplogroup L (including haplogroups L0-L6) is limited to sub-Saharan Africa. Several L haplogroup lineages occur most frequently in eastern Africa (e.g., L0a, L0f, L5, and L3g), but some are specific to certain ethnic groups, such as haplogroup lineages L0d and L0k that previously have been found nearly exclusively among southern African "click" speakers. Few studies have included multiple mtDNA genome samples belonging to haplogroups that occur in eastern and southern Africa but are rare or absent elsewhere. This lack of sampling in eastern Africa makes it difficult to infer relationships among mtDNA haplogroups or to examine events that occurred early in human history. We sequenced 62 complete mtDNA genomes of ethnically diverse Tanzanians, southern African Khoisan speakers, and Bakola Pygmies and compared them with a global pool of 226 mtDNA genomes.. We propose that a large and diverse human population has persisted in eastern Africa and that eastern Africa may have been an ancient source of dispersion of modern humans both within and outside of Africa."
-- Gonder, M, et al (2007). Whole mtDNA Genome Sequence Analysis of Ancient African Lineages. Mol Biol
Evol. 24(3):757-68.

Berber populations in Egypt more related to tropical Africans than Europeans or Middle Easterners based on mtDNA.
"The mitochondrial DNA variation of 295 Berber-speakers from Morocco (Asni, Bouhria and Figuig) and the Egyptian oasis of Siwa was evaluated.. A clear and significant genetic differentiation between the Berbers from Maghreb and Egyptian Berbers was also observed. The first are related to European populations as shown by haplogroup H1 and V frequencies, whereas the latter share more affinities with East African and Nile Valley populations as indicated by the high frequency of M1 and the presence of L0a1, L3i, L4*, and L4b2 lineages. Moreover, haplogroup U6 was not observed in Siwa. We conclude that the origins and maternal diversity of Berber populations are old and complex, and these communities bear genetic characteristics resulting from various events of gene flow with surrounding and migrating populations."
-- Coudray et al. (2008). The Complex and Diversified Mitochondrial Gene Pool of Berber Populations. Annals of Human Genetics. Volume 73 Issue 2, Pages 196 - 214
Pale skin color of Europeans and Asiatics a recent event dating to only 6-12kya

Modern humans did not begin to lighten in skin color immediately
after entering Europe some 35,000 years ago. In fact, these ancestral Europeans
remained brown-skinned for tens of thousands of years. This is the conclusion
now emerging from studies of skin color loci.
In 2005, a team of Japanese researchers found that the depigmentation of
European skin was partly due to a relatively recent allele at the SLC45A2 (AIM1)
gene. They dated the allele to c. 11,000 BP and concluded that it had rapidly
supplanted the original allele through positive selection (Soejima et al.,
2005).
Then last year, at the Annual Meeting of the American Association of Physical
Anthropologists, a molecular anthropologist at the University of Arizona, Heather
Norton, presented evidence that Europeans have a similarly recent allele at
another skin color gene, SLC24A5. The new allele is dated to 12,000 3,000
BP. As she stated during her talk: "The [evolution of] light skin occurred
long after the arrival of modern humans in Europe." (Norton
& Hammer, 2007).
Either way, the implication is that our
European ancestors were brown-skinned for tens of thousands of years--a
suggestion made 30 years ago by Stanford University geneticist L. Luca
Cavalli-Sforza. He argued that the early immigrants to Europe, who were
hunter-gatherers, herders, and fishers, survived on ready-made sources of
vitamin D in their diet. But when farming spread in the past 6000 years, he
argued, Europeans had fewer sources of vitamin D in their food and needed to
absorb more sunlight to produce the vitamin in their skin. Cultural factors such
as heavier clothing might also have favored increased absorption of sunlight on
the few exposed areas of skin, such as hands and faces, says paleoanthropologist
Nina Jablonski of PSU in State College. (Gibbons,
2007)
From:
Gibbons, A. (2007). American Association Of
Physical Anthropologists Meeting: European Skin Turned Pale Only Recently, Gene
Suggests. Science 20 April 2007: 316. no. 5823, p. 364
and
Norton, H.L. & Hammer, M.F. (2007). Sequence variation in the pigmentation candidate gene SLC24A5 and evidence for independent evolution of light skin in European and East Asian populations. Program of the 77th Annual Meeting of the American Association of Physical Anthropologists, p. 179.

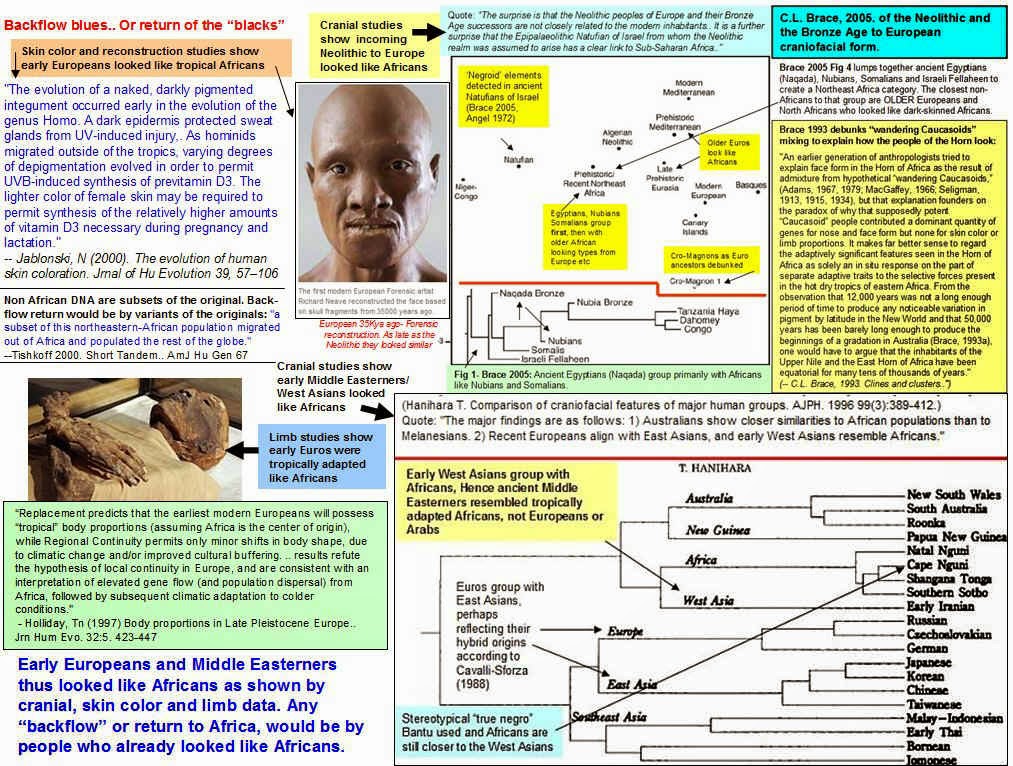
Reconstruction of early European based
on scientific data about skin color
First modern Europeans looked like Africans
Scientists reveal face of the first European
The face of the first European has been recreated from bone fragments by scientists.
By Urmee Khan, Digital and Media Correspondent
Published: 8:22PM BST 04 May 2009
The first modern European Forensic artist Richard Neave reconstructed the face based on skull fragments from 35000 years ago. The head was rebuilt in clay based on an incomplete skull and jawbone discovered in a cave in the south west of the Carpathian Mountains in Romania by
potholers.
Using radiocarbon analysis scientists say the man or woman, it is still not possible to determine the sex, lived between 34,000 and 36,000 years ago.
Europe was then occupied by both Neanderthal man, who had been in the region for thousands of years, and anatomically-modern humans Homo sapiens.
The skull appears very like humans today, but it also displays more archaic traits, such as very large molar teeth, which led some scientists to speculate the skull may belong to a hybrid between Homo sapiens and Neanderthals an idea discounted by other experts.
Erik Trinkaus, professor of anthropology at Washington University in Missouri, said the jaw was the oldest, directly-dated modern human fossil. "Taken together, the material is the first that securely documents what modern humans looked like when they spread into Europe," he said.
The model was created by Richard Neave, a forensic artist, for a BBC programme about the origins of the human race and evolution.
Evolution shows that the original human skin color is dark and humans until quite recently had dark skin, until they migrated from the tropics.
"The evolution of a naked, darkly pigmented integument occurred early in the evolution of the genus Homo. A dark epidermis protected sweat glands from UV-induced injury, thus insuring the integrity of somatic thermoregulation. Of greater significance to individual reproductive success was that highly melanized skin protected against UV-induced photolysis of folate.. As hominids migrated outside of the tropics, varying degrees of depigmentation evolved in order to permit UVB-induced synthesis of previtamin D3. The lighter color of female skin may be required to permit synthesis of the relatively higher amounts of vitamin D3 necessary during pregnancy and lactation."
-- Jablonski, N (2000). The evolution of human skin coloration. Journal of Human Evolution 39, 57106
"Humans skin is the most visible aspect of the
human phenotype. It is distinguished mainly by its naked appearance, greatly
enhanced abilities to dissipate body heat through sweating, and the great range
of genetically determined skin colors present within a single species. Many
aspects of the evolution of human skin and skin color can be reconstructed using
comparative anatomy, physiology, and genomics. Enhancement of thermal sweating
was a key innovation in human evolution that allowed maintenance of homeostasis
(including constant brain temperature) during sustained physical activity in hot
environments. Dark skin evolved pari passu with the loss of body hair and was
the original state for the genus Homo. Melanin pigmentation is adaptive and has
been maintained by natural selection."
-- Jablonski N (2004)THE EVOLUTION OF HUMAN SKIN AND SKIN COLOR. Annual
Review of Anthropology Vol. 33: 585-623
Early Europeans and Middle Easterners thus looked like Africans as shown by cranial, skin color and limb data. Any backflow or return to Africa, would be by people who already looked like Africans.

Other researchers note weakness based on sampling in North African mtDNA study: [quote]
The North African patchy mtDNA landscape has no parallel in other regions of the world and increasing the number of sampled populations has not been accompanied by any substantial increase in our understanding of its phylogeography. Available data up to now rely on sampling small, scattered populations.. It is therefore doubtful that this picture truly represents the complex historical demography of the region rather than being just the result of the type of samplings performed so far.
-- Cherni et al 2005. Female gene pools of Berber and Arab .. microstructure of mtDNA variation in North Africa. Hum Biol. 2005 Feb;77(1):61-70.
Home | Quotations | Misc Notes | Notes 2 | Hair | DemicDiff |
DiversityEgypt in Africa | Black-Greek-DNA links | Notes 3 |Notes 4| Misc news clips | Ethiopians
|
Link to research papers and articles: (wysinger.homestead.com/keita.html) |
|
Link to current African DNA research: (http://exploring-africa.blogspot.com/) |
|
Google Search- other data |
