Knochen
www.j-seegers.de
1. Baumaterial
1.1 Knochenzellen
Vorläuferzellen
liegen in der Nähe von äußerer
und innerer Knorpeloberfläche sowie in den Havers-Kanälen.
Sie können sich zu Osteoblasten
umwandeln und zeitlebens bei Knochenbrüchen aktiviert werden.
Osteoblasten
synthetisieren die organischen Bestandteile der
Knochengrundsubstanz (Kollagen, Proteogylcane, Glycoproteine). Sie liegen
epithelähnlich einshichtig iso- bis hochprismatisch (ûø Synthesephase)
bzw. flach (ûø abnehmende Aktivität) an der Oberfläche von Knochenbälkchen.
In der Synthesephase sind sie logischerweise basophil und PAS-positiv.
Syntheseprodukte werden nur an die mit der Knochengrundsubstanz in Berührung
stehende Fläche abgegeben (= Polarisation). Diese noch nicht verkalkte
Grundsubstanz heißt Osteoid.
Osteozyten
nennt man Osteoblasten, die ringsherum von Knochengrundsubstanz
umgeben sind (meist in verkalkter Grungsubstanz und in Schichten angeordnet).
Sie stehen durch Gap
junctions miteinander in Verbindung.
Osteoklasten
sind große, stark verzweigte, bewegliche Zellen (stammen von
Monozytenvorläuferzellen
ab und gehören zum Phagozytosesystem).
-
erheblich vergrößerter azidophiler (=eosinophiler) Zelleib
-
50 oder mehr Kerne
Sie sind in der Lage, Knochensubstanz abzubauen. Ihre Fraßspur heißt
Howshipsche
Lakune. Ein Osteoklast baut die Knochensubstanz ab, die 100-150 Osteoblasten
in der gleichen Zeit aufbauen.
Steuerung und Aktivierung erfolgt über Hormone: |
 |
Vitamin D3 erhöht Ca++ im
Blut (ûø Knochenabbau bei Calciummangel), Parathormon (Nebenschilddrüse!)
aktiviert Osteoklasten (ûø Knochenabbau ûø Ca++
erhöht sich im Blut), Calcitonin hemmt Osteoklasten (gegenteiliger
Effekt).
1.2 Intercellularsubstanzen
Besteht zu 50% aus Mineralien (Phosphat, Calcium, ...), zu 25% aus organischen
Verbiondungen (Kollagenfasern!)
und zu 25% aus Hydratationswasser.
2. Knochenarten
2.1 Geflechtknochen
Bei jeder Knochenneubildung wird dieses Stadium
durchlaufen. Er enthält einen zufälligen, orientierungslosen
Verlauf an teils groben, teils feinen Kollagenfaserbündels, Lamellen
fehlen. Der Osteozytenbestand ist höher, der Mineraliengehalt niedriger
aus bei Lamellenknochen. In der Regel wird er aus mechanisch-belastbaren
Gründen in Lamellenknochen umgebaut.
2.2 Lamellenknochen
| Für den Erwachsenen typisch. Sie enthalten
ein feines Netz von Kollagenfasern
(in dem es eine Hauptfaserrichtung gibt) sowie Osteozyten,
die in den Lamellen sowie an ihren Grenzen liegen: Sie befinden sich in
abgeflachten Knochenhöhlen, von denen radiär Knochenkanälchen
(V) für Osteozytenfortsätze ausgehen. Diese Volkmann-Kanälchen
(V) stehen miteinander in Verbindung und bekommen Anschluß an den
Zentralkanal (Z), welcher Blutgefäße,
Nerven und Bindegewebe enthält. Von hier erfolgt die Ernährung.
Schaltlamellen sind reste einer früheren Osteonengeneration und haben
keinen direkten Gefäßanschluß mehr. Dahingegen begrenzen
Generallamellen den Knochen: Die äußeren Generallamellen liegen
unter dem Periost, die inneren zur Knochenhöhle hin. |
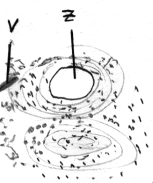
|
3. Periost und Endost
Äußere und innere Knochenoberfläche
werden von Bindegewebe bedeckt, Periost (außen) und Endost (innen).
Beide haben die Fähigkeit, Knochen neu zu bilden. Fehlen sie, wird
der Knochen durch Osteoklasten
wieder abgebaut. Im Periost kommen neben zahlreichen (sensorischen!) Nerven
und Gefäßen auch Fibroblasten
und Vorläuferzellen vor.
Bei Knochenbrüchen geht die Knochenneubildung
vom Periost und Endost aus. Zunächst kommt es an der Bruchstelle
zum Bluterguß, dann bauen Makrophagen das an der Bruchstelle zugrunde
gegangene Gewebe ab. Anschließend kommt es zu Knorpelbildung und
dann zu enchondraler (s. u. 4.2.2), aber auch zu desmaler (s. u. 4.1) Ossifikation.
4. Knochenentwicklung
4.1 Desmale Ossifikation
Direkte Knochenentwicklung, häufigste Art
der Knochenentwicklung. Seltenst jedoch entstehen ganze Knochen durch desmale
Ossifikation allein (nur z. B. einige Teile der Schädelknochen). Desmale
Ossifikation läßt am Schaft aller Röhrenknochen eine Knochenmanschette
entstehen: Sie beginnt mit der Verdichtung einiger Mesenchymzellen.
Das Wachstum angelegter Knochen erfolgt apositionell, d. h. an der einen
(äußeren) Oberfläche wird neuer Knochen gebildet, an der
anderen (inneren) wird er abgebaut.
4.2 Chondrale
Ossifikation
4.2.1 Perichondrale Ossifikation
Knochenneubildung geht vom Perichondrium
aus und wird vom Periost weitergeführt (also ein Vorgang, der von
außen
für außen stattfindet ûø es wird quasi der Rahmen abgesteckt,
das ist der entscheidende Unterschied zu 4.2.2). Der hyaline Knorpel entwickelt
sich über Säulenknorpel (Hypertrophie der Chondrozyten
durch Ernährungsmangel und Vergrößerung der Knorpelhöhlen)
zu Blasenknorpel (fast Ø Intercellularsubstanz mehr vorhanden),
der von Chondroklasten abgebaut wird.
Osteoklasten
fressen Löcher in die Oberfläche dieser gebildeten perichondralen
Knochenmanschette, Löcher, in die Blutgefäße und Mesenchymzellen
aus dem Periost einwachsen. Durch verschiedene Umbauvorgänge umschließt
der Knochenschaft einen weitgehend knochen- und knorpelfreien Raum, die
primäre Markhöle, den Blutgefäße und Mesenchymzellen,
die zu Stammzellen verschiedenster Art werden, ausfüllen. Ab dem 5.
Embryonalmonat ist der blutbildende Stammzellentyp vorherrschend, ab diesem
Zeitpunkt spricht man von sekundärer Markhöle.
4.2.2 Enchondrale Ossifikation
Der Knorpel des Knochenmodells wird abgebaut
und durch Geflechtknochen ersetzt (Vorgang, der innen stattfindet, Unterschied
zu 4.2.1). Dieser Vorgang beginnt von innen an der perichondralen Knochenmanschette.
Solange der Knochen wächst, bleiben die Epiphysenfugen vorhanden.
Man unterscheidet die Reservezone (in der deutlich hyaliner Knorpel
vorliegt) von der Poliferationszone (in der der hyaline Knorpel
als Säulenknorpel vorliegt und weniger Intercellularsubstanz vorhanden
ist). In der Resorptionszone liegt der Knorpel aufgrund von Nahrungsmangel
als Blasenknorpel vor (siehe 4.2.1). In der Verknöcherungszone gehen
die Knorpelzellen zugrunde, die Knorpelhölen werden durch Chondroklasten
eröffnet (ûø "Eröffnungszone"), Einwachsen von Blutkapillaren
und undifferenzierten Zellen, die sich zum Osteoblasten differenzieren
und dann Knochengrundsubstanz bilden. Verkalkter Knorpel ist basophil,
Knochengewebe azidophil (=eosinophil).
Ohne Gewähr. Irrtümer, Fehler sowie Unvollständigkeit
vorbehalten.
www.j-seegers.de