| Effects of Landscape Variation on the Ecology of Black Bears |
| Chhatra M. Sharma Agricultural University of Norway, Noragric, P.O.Box 5003, N-1432 �s, Norway |
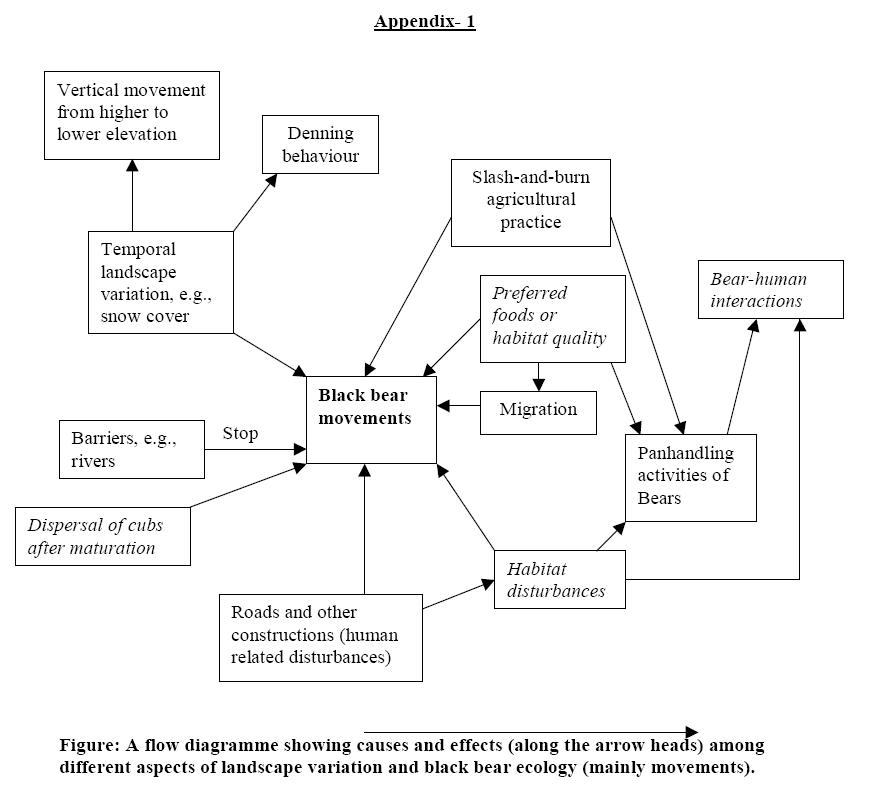
The paper deals with the variation in landscape and its effects on the ecology of black bears. Black bears are omnivorous animals found in the temperate region of the world and Asian black bears (Selenarctos thibetanus) are ecologically very similar to the American black bears (Ursus americanus) (Schaller et al. 1989, Reid et al. 1991). The main target of this paper is to find out the relationships of landscape change (temporal as well as spatial) with feeding habits, habitat choices, movements, and causes of denning behaviours of black bears. At the same time, bear-human interactions are also discussed. Main foods of black bears include fruits and berries, acorns, grass, buds and blossoms, nuts, tubers and roots, new bamboo shoots, and farm crops etc. But they are very much adapted to consume salmons wherever available (mainly American black bears). Therefore, they prefer the landscape that mainly consists of mixed broad-leaved temperate forests having sufficient water, cover, denning sites and least human related disturbances. They show vertical migration from higher to lower elevations each year due to temporal change in the landscape where their main habitat is covered with snow. The spatial change in the landscape due to slash-and-burn agricultural practices, permanent agricultural practices and roads also affect the movement patterns of black bears. Rivers act as the barrier for black bear movement and dispersal, therefore, translocation of some female bears can help to recover the fragmented population. Human related disturbances are mainly responsible for the bear-human-interactions that some times cause death of the people. The overall findings indicate that the panhandling activities of bears are mainly due to human related activities that affect the ecological behaviour of black bears. Therefore, management of black bear habitat should avoid human activities to a certain level. Key words: Black bear, Landscape variation, Habitat preferences, Food habit, Movements, Denning, Barrier, Bear-human interactions, Habitat disturbances. Himalayan black bear (Selenarctos thibetanus) is distributed in a wide range of temperate habitats in Asia from southern China to Russian Manchuria, and Iran to Japan (Reid et al. 1991). The average weight of an adult male bear ranges from 90-160 kg and that of an adult female from 70-130 kg whereas some well-fed individuals may weigh as much as 200 kg (males) and 170 kg (females) (Walker 1975, MacDonald 1984). It is general biological expectation that male bears are larger at all ages than female bears (McRoberts et al. 1998). The coat colour is generally jet black, with a distinct while crescent shaped mark on the chest (Shrestha et al. 1997). The life expectancy is said to be approximately 24 years in the wild (MacDonald 1984). Ecologically, Himalyan black bear is equivalent to American black bear (Ursus americanus) in term of the body form, size and food habits (Schaller et al. 1989, Reid et al. 1991). Therefore, this paper mainly aims to study the ecological behaviour of black bears based on the landscape variability, irrespective of the Himalayan (Asiatic) or American black bears since both are considered as very similar in most of the aspects. ?The actual relationships of individuals, populations, and species to their habitat, however, is much more complex and variable, spatially and temporally, than can be portrayed by a written definition? (Schoen 1990). Therefore, habitat preferences of black bear depend on many factors and we cannot claim only few factors responsible for it. As bears are wide-ranging species of landscapes, habitat relationships must be evaluated in a broader concept than habitat types, e.g., human activities and land uses must be incorporated into bear habitat relationships (Schoen 1990). Landscape variability has also some effects on the habitat selection, food preferences, movements, denning behaviour, and other behavioural activities of black bears that can be justified on the basis of following studies by many different authors. The components of prime black bear habitat includes the landscape with sufficient food (Pelchat and Ruff 1986; Grenfell and Brody 1986), water, cover, denning sites, and least human related disturbances, therefore, mixed broad-leaved temperate forest is an ideal habitat for black bears (Shrestha et al. 1997, Saitoh et al. 2001). Hazumi and Maruyama (1986) mention dense forests, heavy undergrowth, and complex topography as the natural black bear habitat. It is suggested that large hardwood swamp systems, heterogeneous landscapes, saw timber stands and road less areas are important habitat components to maintain a viable bear population (Mykytka and Pelton 1990). Black bear is omnivorous in habit. Spawning and migrating salmonids are a major food of black bears in coastal Alaska (Reinhart and Mattson 1990); but bears rely heavily on small, wild fruits such as berries to provide energy reserves necessary for reproduction and winter hibernation, especially in areas where energy-dense food (e.g. salmon) are scarce or unavailable (Welch et al. 1997). It requires relatively nutrient rich food such as grass, buds and blossoms, acorns, nuts, fruits and berries, tubers and roots, new bamboo shoots, and farm crops (e.g. maize) (Shrestha et al. 1997). However, areas of concentrated food need not be large to attract bears (Noyce and Coy 1990). Depending on the food availability, bears may also feed on carrion, insects, eggs of birds etc. And they are also known to feed on corn in the agriculture farms (Shrestha et al. 1997). According to Welch et al. (1997), bears are non-ceacal monogastrics that cannot digest fibres efficiently, therefore, small wild fruits play an important role in sustaining many bear populations. Habitat utilization by black bears seems to depend on the seasons. Fall core areas of bears were related to food distribution, primarily acorns which were found to be the most important food during this season (Hazumi and Maruyama 1986). But the summer core areas, as they mention, were apparently related to cover, not food availability. The seasonal changes in habitat use patterns resulting from the temporal distribution of key foods (Young and Beecham 1986). Kansas and Raine (1990) recognize some factors as year-to-year food availability, bear densities, elevation, cover and fire history influencing black bear habitat use. Powell and Seaman (1990) pointed out that patchiness of food production might be as important as the total production in analysing the effects on bear biology. The Florida black bears occurring around large swamps and surrounding pinelands is explained on the basis of food availability and presence of escape cover (Mykytka and Pelton 1990). Heterogeneity, a difference between locations, is regarded as a fundamental cause of species movements that is explained as the 'species flow principle' of landscape ecology (Forman and Gordon 1986). The home range of individual black bears usually covers more than several square kilo meters and it would not be difficult for them to cross a shallow river less than 100 m wide (Saitoh et al. 2001). They further analyse and argue that such rivers appear to be a barrier for gene flow among the bear populations. In a landscape with dense rivercourses male bears more frequently cross the rivers than female bears (White et al. 2000). The home range for black bears was suspected to be more than 200 ha in general (Hazumi and Maruyama 1986) which probably change in size and location in response to the availability of food in their habitats. American black bears were estimated to cover about 39 sq.km (by females)-102 sq. km (by males) (Pelchat and Ruff 1986). Black bear movements within their home range are regarded as their behavioural manifestations of basic biological requirements as food, shelter, and mates (Seaman and Powell 1990). Saitoh et al. (2001) write that it may be difficult for bears that disperse from their natal sites to settle and breed in a new habitat. Black bears are found to be wandering from one core to another core area ranging from 20-115 ha in Nikko, Japan, therefore, home ranges may be composed of several core areas (Hazumi and Maruyama 1986). Black bears maintain their territory by urinating on small trees and rubbing on large trees; and they also investigated the marks of other bears (Rogers and Wilker 1990). Some winter-summer animal movement between higher and lower elevations (vertical movement) in mountains is considered as transitional between migration and home range movements (Forman and Gordon 1986). Black bears movement is affected by the availability of the preferred food (Pelchat and Ruff 1986; Raine and Kansas 1990; and Shrestha et al. 1997). Pelchat and Ruff (1986) found that bears were engaged in foraging excursions away from their home ranges in response to the shortage of preferred food and occupied areas larger than in the years when food were abundant. Bears invade agricultural fields and raid crops when natural foods become scarce (Pelchat and Ruff 1986; Shrestha et al. 1997, Saitoh et al. 2001). Noyce and Coy (1990) write that seasonal movements of black bears to dumps, oak stands, pine nut stands, agricultural fields, and productive berry patches are reported by many researchers. Smith and Pelton (1990), after reviewing many studies, noted that the space utilization by black bears may be related to sex, age, reproductive condition, kinship, social rank and population density; however, food abundance and distribution are the most important in determining the black bears' movements. The denning of Ursids is generally interpreted as a strategy to avoid unfavourable environmental conditions and reduce energy expenditures (Watts 1990). After consulting many literatures, Watts (1990) wrote that the black and grizzly bears are characterized by denning during the winter by both sexes and all age classes, whereas, the polar bear is limited primarily to maternity denning. Black bears mostly go under hibernation during harsh winters; and remain immobile in winter dens for 3 to 5 months (Floyed and Nelson 1990). But black bears of some places do not necessarily under go hibernation; winter active bears use lower and warmer places consuming herbaceous foods and acorns remaining on the ground (Graber 1990). Smith (1986) asserts that the length of hibernation (denning) of black bears increases with the length, and possibly, the severity of winters. Wathen et al (1986) reviewed several studies of den selection and concluded that bears exhibit considerable versality in den selection in various parts of their range; and most dens are excavations beneath standing trees, stumps, and fallen logs, or excavations directly into the hillside. Den sites are located in sub alpine areas where deep snows accumulate and temperatures remain cold (Hazumi and Maruyama 1986). Wathen et al. (1986) found tree cavities high above ground as the dens used by black bears in south eastern United States (oaks and northern oaks are the primary species outside the GSMNP). Smith and Van Daele (1990) reported very less disturbance of denning and permanent denning habitat loss during the construction of hydroelectric development than had been predicted. Mack (1990) recorded most of the dens on northern aspects on relatively steep slopes which he tried to explain that steep slopes provide better soil drainage and the northern aspect reduces the probability of water entering the dens. In contrast, Miller (1990) found that there was no apparent selection for aspect in black bear dens; however, brown bear preferred south aspects. A population of black bear can be fragmented into two or more subpopulations due to barriers, e.g., rivers. Populations of black bears in south eastern North America are very fragmented due to possible barrier effects of rivers to normal bear movements and dispersal (White et al. 2000). Similar results were obtained by Saitoh et al. (2001) while studying fragmented populations of Asian black bears in Japan where human activities along large rivers are the main causes for population fragmentation in relatively short time scale. Subpopulation sizes of black bears in some parts of Japan are too small to be self sustaining, therefore, connection of these subpopulations through corridors is very important (Saitoh et al. 2001). Mississippi river acted as semi-permeable barrier to bear movements and dispersal patterns where male bears were less influenced than female bears, therefore, female bear translocation across rivers is recommended by White et al. (2000) to recover the fragmented bear populations. The polar bear (Ursus maritimus) is the largest non-aquatic carnivores on earth, and is by far the most predaceous of all living bears whereas the black bear (Ursus americanus) is normally tolerant of people and reclusively dwells in the forest (Herrero and Fleck 1990). Mattson (1990) studied about the human impacts on bear habitat and mentioned that bears and humans are flexible, adaptable omnivores and are natural competitors. He further argues that humans have disrupted otherwise stable, dynamically equilibrated processes in virtually all wild land ecosystems occupied by bears. Timber harvest is regarded as one of the detrimental effects on bears mainly due to disturbances (e.g., roads, development, noise) that might interfere with exploitation of the habitat by individual bears (Grenfell and Brody 1986). McLean and Pelton (1990) suggested a clue that the habitat quality may dictate the amount of panhandler activity; and assumed that increase in the production of natural foods would not only boost the carrying capacity of the local habitat, but conceivably, decrease the amount of such panhandler activity. After reviewing many studies on panhandler activities of bears, McLean and Pelton (1990) concluded that panhandler activity in the smoky mountains is common because of variety of factors: the violation of national park service regulations regarding food and bears, visitor density, bear density, and habitat condition etc., for example, park officials reported an average of 158 incidents per year of personal damage caused by black bears. Similar incidents were reported with grizzly bears in Kluane National Park, where during 1981-84, only 3 serious incidents between bears and people occurred which dramatically increased in 1985 and reached 10 serious incidents due to a change in the behaviour of some grizzlies towards people (Leonard and Breneman 1990). According to the local people in MBNPCA, black bears are shy and try to run away from the view of the people. But they are found to raid their crops during night when there is shortage of natural foods for bear in nearby forest (personal communication). Such nuisance behaviour of bears may be due to the habitat loss or change in the landscape structure favourable to them. Kasworn and Manley (1990) write that avoidance of human activities by wildlife has become an important consideration in habitat management. They found that road has a great influence on the habitat use by black bears because they avoided habitat within 274m of open roads. According to Young and Beecham (1986), avoidance of roads is mainly shown by female black bears in proportion to availability. Black bears are found to be more tolerant than grizzly bears towards the disturbance due to roads (Kasworn and Manley 1990). One of the major factors was the road density that affected bear vulnerability during the 1976 through 1978 hunting seasons in Florida (Mykytka and Pelton 1990). Young and Beecham (1986) studied habitat use by both the sexes of bears and stated the tendency for male black bears to use roads in proportion to availability and female black bears to avoid roads has strong management implications?. Smith and Van Daele (1990), in their study on the impacts of hydroelectric development on brown bears, found out that bears residing near the construction areas made only minor shifts to areas with dense cover during construction; however, the long term impacts are expected to have much more serious implications to brown bear habitat management than the immediate effects of construction. Slash-and-burn agricultural practices in temperate regions nearby the black bear habitats also influence the ecology of black bears because such practices create patches in the forests (Shrestha et al. 1997). Under-stories are also destroyed completely in such areas and the place becomes unsuitable for black bear habitat. Though black bears are omnivorous in habit and consume many different kinds of foods, they mostly rely on some energy rich foods, therefore, they mostly move to those places where there are sufficient energy rich foods. This indicates their spatial movements in search of the good habitat and their home range contains many core areas (Hazumi and Maruyama 1986). In winter, they start to move towards lower elevations to escape themselves from the harsh environment (Shrestha et al. 1997). From above mentioned descriptions, it can be realised that landscape variability is one of the important causes for the movement of black bears within or across the landscape for different purposes, e.g., feeding, hiding (denning), dispersing etc. Food availability is the number one affecting factor for the movement of black bears (Pelchat and Ruff 1986, Raine and Kansas 1990, and Shrestha et al. 1997). The dispersal of new ones from the natal area to establish their own territory is one of the difficult tasks for black bears (Saitoh et al. 2001). Vertical movement of black bears during winter period is mainly for the avoidance of unfavourable condition due to change in landscape (snow cover) as indicated by Forman and Gordon (1986). As indicated by Watt (1990), denning, one of the important behaviours of black bears, is essential for them to reduce the energy expenditure in the harsh winter periods when there is temporal variation in their habitat as the habitat is completely covered with the snow. Denning is considered to be optional for them depending in the condition of the landscape and climate change (Smith 1986, Wathen et al. 1986, Hazumi and Maruyama 1986, Mack 1990, Miller 1990). Rivers sometimes act as barriers (or semi-barriers) that can separate a population to subpopulations. Human related activities are the most important factors for the bear-human interactions that can sometimes be fatal. Panhandling activities of bears emerged mainly due to the change in landscape of bear habitat by human created activities, e.g., change in agricultural practices, construction works etc. (Leonard and Breneman 1990, McLean and Pelton 1990, Grenfell and Brody 1986). It is suggested that avoidance of human activities can be an important management practice to conserve black bear habitat (Kasworn and Manley 1990, Smith and Van Daele 1990). I am highly indebted to my teacher Gary Fry, Department of Biology and Nature Conservation, Agricultural University of Norway, for his guidance during the preparation of this paper. I also thank all the classmates of Landscape Ecology for pointing out the relevant suggestions during the presentation of this paper. Floyed, T., and R.A. Nelson. 1990. Bone Metabolism in Black Bears. Int. Conf. Bear Res. and Manage.8:135-137. Forman, R.T.T., and M. Gordon. 1986. Landscape Ecology. John Wiley and Sons, Inc, USA. Graber, D.M. 1990. Winter Behavior of Black Bears in the Sierra Neveda, California. Int. Conf. Bear Res. and Manage. 8: 269-272. Grenfell, W.E., and A.J. Brody. 1986. Black Bear Habitat Use in Tahoe National Forest, California. Int. Conf. Bear Res. and Manage.6:65-72. Hazumi, T., and N. Maruyama. 1986. Movements and Home Ranges of Japanese Black Bears in Nikko. Int. Conf. Bear Res. and Manage.6:137-143. Herrero, S., and S. Fleck. 1990. Injury to People Inflicted by Black, Grizzly or Polar Bears: Recent Trends and New Insights. Int. Conf. Bear Res. and Manage. 8:25-32. Kansas, J.L., and R.M. Raine. 1990. Methodologies Used to Assess the Relative Importance of Ecological Land Classification Units to Black Bears in Banff National Park, Alberta. Int. Conf. Bear Res. and Manage.8:155-160. Kasworn, W.F., and T.L. Manley. 1990. Road and Trail Influences on Grixxly Bears and Black Bears in Northwest Montana. Int. Conf. Bear Res. and Manage.8:79-84. Leonard, R.D., R. Breneman, and R. Frey. 1990. A Case History of Grizzly Bear Management in the Slims River Area, Kluane National Park Reserve, Yukon. Int. Conf. Bear Res. and Manage. 8:113-123. MacDonald, D. (ed.). 1984. The Encyclopedia of Mammals. Facts on File Publications, New York.895pp + appendices. Mack, J.A. 1990. Black bear Dens in the Beartooth Face, South-Central Montana. Int. Conf. Bear Res. and Manage. 8:273-277. Mattson, D.J. 1990. Human Impacts on Bear Habitat Use. Int. Conf. Bear Res. and Manage.8:33-56. McLean, P.K., and M.R. Pelton. 1990. Some Demographic Comparisons of Wild and Panhandler Bears in the Smoky Mountains. Int. Conf. Bear Res. and Manage. 8:105-112. McRoberts, R.E., R.T. Brooks, and L.L. Rogers. 1998. Using Non-Linear Mixed Effects Models to Estimate Size-Age Relationships for Black Bears. Can. J. Zool.76: 1098-1106. Miller, S.D. 1990. Denning Ecology of Brown Bears in Southcentral Alaska and Comparisons with a Sympatric Black Bear Population. Int. Conf. Bear Res. and Manage. 8:279-287. Mykytka, J.M., and M.R. Pelton. 1990. Management Strategies for Florida Black Bears Based on Home Range Habitat Composition. Int. Conf. Bear Res. and Manage.8:161-167. Noyce, K.V., and P.L. Coy. 1990. Abundance and Productivity of Bear Food Species in Different Forest Types of Northcentral Minnesota. Int. Conf. Bear Res. and Manage. 8:169-181. Pelchat, B.O., and R.L. Ruff. 1986. Habitat and Spatial Relationships of Black Bears in Boreal Mixedwood Forest of Alberta. Int. Conf. Bear Res. and Manage.6:81-92 Powell, R.A., and D.E. Seaman. 1990. Production of Important Black Bear Foods in the Southern Appalachians. Int. Conf. Bear Res. and Manage. 8:183-187. Raine, R.M., and J.L. Kansas. 1990. Black Bear Seasonal Food Habits and Distribution by Elevation in Banff National Park, Alberta. Int. Conf. Bear Res. and Manage. 8:297-304. Reid, D., M. Jiang, Q. Teng, Z. Qin, and J. Hu. 1991. Ecology of the Asiatic Black Bear (Ursus thibetanus) is Sichuan, China. Mammalia. 55(2): 221-237. Reinhart, D.P., and D.J. Mattson. 1990. Bear Use of Cutthroat Trout Spowning Streams in Yellowstone National Park. Int. Conf. Bear Res. and Manage. 8:343-350. Rogers, L.L., and G.W. Wilker. 1990. How to Obtain Behavioral and Ecological Data from Free Ranging, Researcher-habituated Black Bears. Int. Conf. Bear Res. and Manage. 8:321-327. Saitoh T., Y. Ishibanshi, H. Kanamori, and E. Kitahara. 2001. Genetic Status of Fragmented Populations of the Asian Black Bear (Ursus thibetanus) in Western Japan. Popul. Ecol., 43: 221-227. Schaller, G.B., T. Qitao, K.G. Johnson, W. Xiaoming, S. Heming, and H. Jinchu. 1989. The Feeding Ecology of Giant Pandas and Asiatic Black Bears in the Tangjiahe Reserve, China. Pp. 212-241 In: J. L. Gittleman (ed.) Carnivore Behavior, Ecology and Evolution. Cornell University Press, Ithaca, NY. 620 pp. Schoen, J.W. 1990. Bear Habitat Management: A Review and Future Perspective. Int. Conf. Bear Res. and Manage. 8:143-154. Seaman, D.E., and R.A. Powell. 1990. Identifying Patterns and Intensity of Home Range Use. Int. Conf. Bear Res. and Manage.8:243-249. Shrestha, M.K., C.M. Sharma, and A. Bhattarai. 1997. Wildlife Depredation of Crop and Livestok: A Study in the Makalu-Barun National Park and Conservation Area. A Draft Report of People Wildlife Project (USAID/DNPWC/TMI), Kathmandu, Nepal. Smith, R.B., and L.J. Van Daele. 1990. Impacts of Hydroelectric Development on Brown Bears, Kodiac Island, Alaska. Int. Conf. Bear Res. and Manage.8:93-103. Smith, T.R. 1986. Activity and Behavior of Denned Black Bears in the Lower Mississippi River Valley. Int. Conf. Bear Res. and Manage.6:137-143. Smith, T.R., and M.R. Pelton. 1990. Home Ranges and Movements of Black Bears in a Bottomland Hardwood Forest in Arkansas. Int. Conf. Bear Res. and Manage.8:213-218. Walker, E.P. 1975. Mammals of the World. Volume 11. Third edition, John Hopkins University. Baltimore. 1500pp. Wathen, W.G., K.G. Johnson, and M.R.Pelton. 1986. Characteristics of Black Bear Dens in the Southern Appalachian Region. Int. Conf. Bear Res. and Manage.6:119-127 Watts, P.D. 1990. Comparative Weight Loss in Three Species of Ursus under Simulated Denning Conditions. Int. Conf. Bear Res. and Manage.8: 139-141. Welch C.A., J. Keay, K.C. Kendall, and C.T. Robbins. 1997. Constraints on Frugivory by Bears. Ecology, 78(4): 1105-1119. White Jr., T.H., J.L. Bowman, B.D. Leopold, H.A. Jacobson, W.P. Smith, and F.J. Vilella. 2000. Influence of Mississippi Alluvial Valley Rivers on Black Bear Movements and Dispersal: Implications for Louisiana Black bear Recovery. Biological Conservation.95: 323-331. Young, D.D., and J.J. Beecham. 1986. Black Bear Habitat Use at Priest Lake, Idaho. Int. Conf. Bear Res. and Manage.6:73-80. |
| Home | Family | Curriculum | Thesis TU | Thesis NLH | Publications | Term Papers | Recent Paper | Proposal | Nepali Poems |
 |
 |
| You are here: Home>>Term Papers>> Effects of ........ |
| Abstract |
| Introduction |
| Habitat and Food Preferences |
| Movements within or across Landscape |
| Denning Behaviour |
| River as a Barrier |
| Bear-Human Interactions |
| Habitat Disturbances and its Management |
| Discussion/Conclusion |
| Acknowledgement |
| References |