| 1° |
2° |
2° |
2° |
2° |
3° |
|
T ou U |
C |
A |
G |
|
|
Phenylanine |
Serine |
Tyrosine |
Cysteine |
U |
| U |
Phenylanine |
Serine |
Tyrosine |
Cysteine |
C |
|
Leucine |
Serine |
x |
x |
A |
|
Leucine |
Serine |
x |
Tryptophane |
G |
|
1 et 2 |
6 |
10 |
17 et 18 |
|
|
|
| 1° |
2° |
2° |
2° |
2° |
3° |
|
T ou U |
C |
A |
G |
|
|
Leucine |
Proline |
Histidine |
Arginine |
U |
| G |
Leucine |
Proline |
Histidine |
Arginine |
C |
|
Leucine |
Proline |
Glutamine |
Arginine |
A |
|
Leucine |
Proline |
Glutamine |
Arginine |
G |
|
2 |
7 |
11 et 12 |
19 |
|
|
|
| 1° |
2° |
2° |
2° |
2° |
3° |
|
T ou U |
C |
A |
G |
|
|
Isoleucine. |
Threonine |
Asparagine |
Serine |
U |
| A |
Isoleucine, |
Threonine |
Asparagine |
Serine |
C |
|
Isoleucine |
Threonine |
Lysine |
Arginine |
A |
|
Methionine |
Threonine |
Lysine |
Arginine |
G |
|
3 et 4 |
8 |
13 et 14 |
6 et 19 |
|
|
|
| 1° |
2° |
2° |
2° |
2° |
3° |
|
T ou U |
C |
A |
G |
|
|
Valine |
Alanine |
Aspartate |
Glycine |
U |
| G |
Valine |
Alanine |
Aspartate |
Glycine |
C |
|
valine |
Alanine |
Glutamate |
Glycine |
A |
|
valine |
Alanine |
Glutamate |
Glycine |
G |
|
5 |
9 |
15 et 16 |
20 |
|
|
|
|
|
Fig. 5.19- Le code génétique. Les
codons sont représentés par des lettres majuscules, et
les acides aminés par leurs noms et des chiffres. Les X
représentent les messages terminaison de la chaîne
d’acides aminés; celle-là est commandée par
des codons dont le code ne peut être traduit (codons
non-sens).
Table 18 |
Dans la nature, il existe un grand nombre d’acides
aminés, environ 300, dont la combinaison peut donner un grand
nombre de protéines. Toutefois, les savants ont constaté
que sont présents, en séquence variable dans une
protéine, pas plus de 20 acides aminés différents
et ceux-ci ont été toujours les mêmes depuis plus
de deux billions d’années d’évolution de la
planète. Quelques protéines contiennent moins de dix de
ces acides amninés, d’autres en contiennent davantage.
Comment le Gène Réalise-t’il la
Synthèse d’une Protéine?
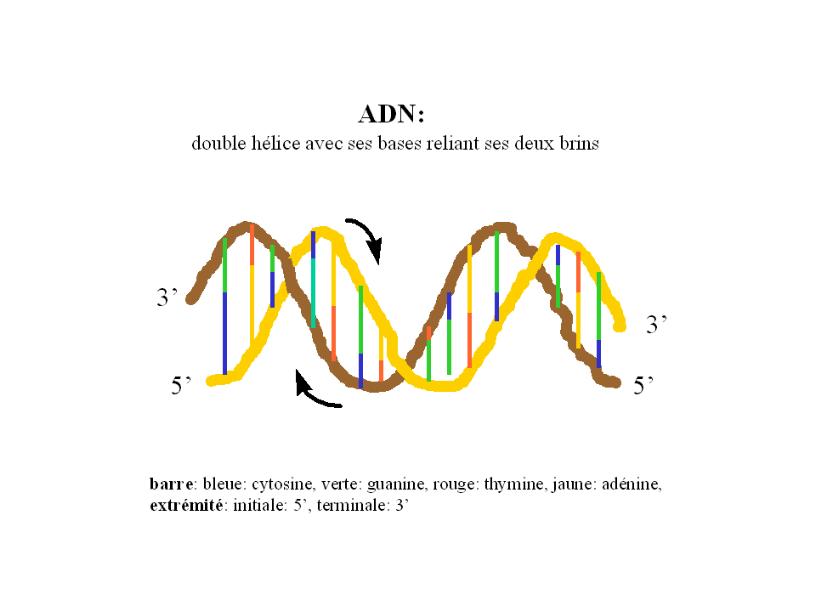
Le mode d’action des gènes met en en action la
molécule d’ADN, celles d’ARN, deux mécanismes
physio-cellulaires - la transcription et la traduction - et le
ribosome, une structure extranucléaire.
- Parce que les deux maillons de l’ADN bicaténaire peuvent se
séparer en deux molécules et que les bases purines et
pyrimidines sont complémentaires, l’ADN monocaténaire, ainsi
formé, peut servir de “template”, de MODÈLE, ou de MOULE,
à la formation de nouvelles chaînes de nucléotides.
C’est ainsi, par exemple, que la molécule d’ARN -
acide ribonucléique - se forme mais avec cette exception que,
dans la molécule d’ARN, l’uracil remplace la thymine
présente dans l’ADN. Un tel mécanismne est la
TRANSCRIPTION du code génétique; les nouveaux
codons, ainsi formés, forment l’ARNm (ARN messager)
- Le code génétique, ainsi passé à
l’ARNm, sera de nouveau transféré à une
nouvelle molécule d’ARN - l’ARNt (ARN de transfert), dont les
triplets de nucléotides complémentaires sont des
ANTICODONS. Ceux-ci, se fixant à un RIBOSOME qui est une unité
structurelle faite d’ARN - l’ARNr - et de protéine,
permet la traduction du code en le phénotype
désiré. Ce mécanisme est la TRANSLATION ou la
TRADUCTION du code génétique (fig. ). Les deux
molécules d’ADN monocaténaire se recouplent ensuite
pour reformer la molécule d’ADN bicaténaire.
Le gène qui code la formation du
phénotype est le gène de structure. Parce que les
gènes de structure sont situés en série sur
la molécule d’ADN, leur mutation est très
susceptible de conduire à une mutation par
changement de cadre (ang., "frameshift mutation”) et,
dès lors, à un changement radical du phénotype ;
en effet, une telle mutation, serait-ce, par exemple, une
délétion, produirait, une séquence de codons
complètement différents de la séquence initiale et
pourrait donc conduire à des variations
phylogénétiques qui se situeraient au dessus du niveau
des espèces, du point de vue de la cladistique.
D’autres gènes, plus isolés, ne
font que régulariser la fonction des gènes de structure
et leurs mutations soint moins susceptibles de conduire à des
changements marqués de phénotypes.
Sur une molécule d’ADN se trouvent, donc, des
séquences bien définies de gènes; la localisation
exacte d'un gène dans la séquence ou sur la
molécule d'ADN est appelée son locus. Dans une séquence, les
gènes qui sont responsables immédiatemnent de la
structure du phénotype sont les gènes de structure.
Fig.5.10 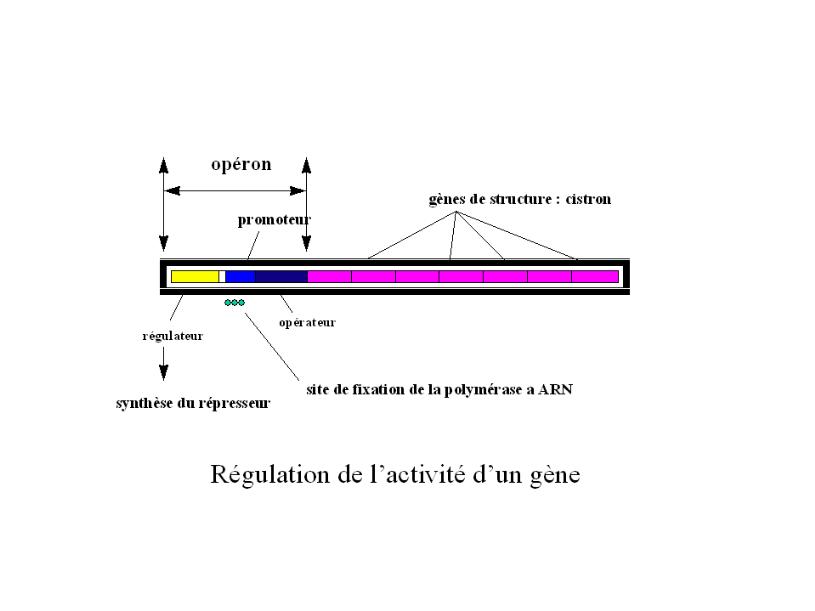
Chez les bactéries, au moins,
à proximité des gènes de structure dont la
séquence est dénommée le cistron, des
gènes constituant l’opérateur commandent
l’expression, ou la transcription, des gènes de structure.
L’ensemble, ainsi formé du cistron et de
l’opérateur, est l’opéron (fig. ). Sur celui-ci, d’autres
gènes - le régulateur
et le promoteur - exercent un
contrôle (régulation et facilitation) et ont,donc,
également, une action sur l’expression du phénotype
(voir pénétrance ci-dessus); leur fonctionnement peut
dépendre des conditions de l’environnement (par exemple,
le substrat) qui peut être plus ou moins approprié.
Au niveau de celui-ci, le régulateur,
réprime, sous l’effet d’un environnemnent
inappropié représentant le répresseur,
l’expression du cistron, i.e., le phénotype, en y bloquant
le promoteur, qui active le fonctionnement de l’opéron.
Dans un environnement approprié, ou inducteur, le blocage
du promoteur est levé et l’opéron produit le
phénotype désiré.
C’est ainsi que s’explique, chez
l’E. coli, le fonctionnement du Lac opéron,
i.e., de l’opéron qui est responsable du catabolisme
(digestion et assimilation) du lactose sous l’action des enzymes
sécrétés par l’opéron (par son
cistron) en présence du lactose. De tels mécanismes
pourraient aussi permettre de comprendre, a côté du
contrôle polygénique, comment surviennent des variations
intraspécifiques ou intragénériques qui sont, du
point de vue de la cladistique, des transformations
phylogéniques moins importantes que celle qui conduisent
d’une famille à une autre, d’un ordre à un
autre, d’une classe à une autre, d’un embranchement
à un autre ou d’un règne à un autre.
Losqu’une mutation affecte
l’opéron qui vient à échapper à
l’action du régulateur, le gène s’exprime de
façon dominante. Lorsque la mutation affecte seulement le
régulateur, le gène déréprimé qui
s’ensuit est un gène récessif (s’exprime de
façon récessive). Une mutation qui affecte
l’opérateur peut éliminer la transcription
entiére de l’opéron.
Il existe egalement de longues régions non
codantes de l'ADN, dénommées introns, intercalées entre les
séquences codantes ou exons.
Les mutations ont plus de chance de survenir au niveau des introns
qui ont, de ce fait, un rôle protecteur qu'au niveau des
régions codantes. Les introns facilitent, également les
arrangements des régions codantes en empêchant leur
interférence réciproque et permettent qu'un petit nombre
de gènes donnent naissance a un grand nombre de
protéines; il existe, donc, une double organisation des
genes, sur deux échelles de grandeur differente: la
1ère échelle est celle du code génétique
primordial des bases puriques et pyrimidiques pour la synthèse
des acides aminés, la 2ième échellle est celle des
loci aidés des introns pour la synthèse des
protéines.
Apparemment, on n'a pas pu mettre en evidence, jusqu'a nos jours, d'operons dans les organismes superieurs, mais il est tres probable que chaque gene soit controle par un promoteur et un represseur, comme dans l'operon bacterien, et l'existence de sequences repetees d'introns (regions non codantes de l'ADN) et d'exons (regions codantes) est indiscutable.
|
Enfin, l’équilibre ponctué, un
sous-produit de la sélection naturelle, pourrait
s’expliquer aussi par un changement drastique de
l’environnement qui profiterait davantage aux phénotypes
provenant des mutations par changement de cadre, i.e. , des
phénotypes très différents du phénotype
initial, qu’à ceux dont les mutations n’auraient
produit que des modulations du phénotype initial.
Comment le gène est-il Conservé pour Établir
l’Hérédité? -- La RÉPLICATIQN
Parce que la propriété
essentielle qui distingue un être vivant d’un objet
inanimé est la capacité de celui-ci à se
reproduire et parce que certains cristaux comme l’argile, par exemple, et certaines
molécules, comme celle d’ADN
ont également cette propriété, il est difficile de
dire quand a commencé la vie. Il est néammoins clair que
la première cellule a surgi des substances précellulaires
de l’environnement.
Comment se fait la réplication de
l’ADN. Au moment de sa réplication, l’ADN bicaténaire s’ouvre à
une extrémité et chacune des deux
branches de la fourchette de réplication ainsi
formée sert de modèle à la formation d’une
nouvelle molécule d’ADN. La molécule
d’ADN porte deux extrémités, 5’ et
3’. Parce que les deux brins complémentaires de
l’ADN bicaténaire sont orientés en sens inverse, un
peu comme votre main gauche paraît être votre main droite
dans un miroir, la synthèse des deux nouveaux brins qui commence
toujours à l’extrémité 5’ est
orientée vers l’intérieur de la fourchette de
réplication, pour l’un de ceux-ci, et vers
l’extérieur de celle-la pour l’autre. La réplication - de
l’extrémité 5’ vers
l’extrémité 3’- des nouveaux brins ayant
été complétée, les deux brins de
l’ADN parental se recouplent et la fourchette de
réplication se ferme, alors que les deux brins nouvellement
synthétisés vont se lier pour former un ADN bicaténaire filial, exactement pareil
à l’ADN bicaténaire parental et contenant,
dès lors, les mêmes
gènes.
Le Gène, une Unité Fonctionelle
Le gène a été défini comme l’unité
héréditaire structurelle. Le gène est aussi
représenté par des
allèles, i.e., des formes
d’un même gène qui existent en nombre pair au
même locus (à la même
position) sur des chromosomes homologues.
cependant, lorsque deux mutations surviennent sur un même
gène, l’effet sera différent selon qu’elles
surviennent au même locus du gène - loci en position cis - ou sur des loci
différents - locus A de l’allèle m et locus B de
l’allèle n de ce gène - loci
en position trans. C’est l’effet cis-trans (de là,
l’appellation de cistron donnée au gène de
structure), la contiguïté des loci
définissant l’unité fonctionelle du
gène. Dès lors, le vieux concept que le
gène est une unité structurelle et non plastique est
périmé. Le gène est
défini maintenant fonctionellement par l’action de son
cistron (un gène --> une protéine), i.e., par
l’action des nucléotides contigüs, et peut être
représenté schématiquement de plus d’une
façon (voir schéma ci-dessous).
Une application de ce nouveau concept du gène
est de rechercher si deux phénotypes mutants qui se ressemblent
renvoient à des allèles d’un même gène
ou à des gènes différents. Ceci peut être
fait par le test cis-trans; celui-ci consiste à croiser
entre eux les individus de la F1 afin
d’en obtenir des hybrides et à rechercher dans la
F1 des individus qui présentent
la complémentation (d’où son appellation aussi de
test de complémentation) parce que celle-ci peut se
présenter seulement avec des formes non alléliques, i.e.,
avec des gènes différents qui agissent sur le même
phénotype. L’expérience suivante de classe de
laboratoire vous éclaircira les idées.
Exercice:
Votre professeur amène en salle de classe des bocaux
contenant, chacun, une femelle drosophile, différant par la
couleur de leurs yeux, un trait pour lequel le gène est
défini par le cistron. Trouvez si toutes les drosophiles
similaires, i.e., présentant un phénotype similaire, ont
un génotype similaire, en relation avec cette
caractéristique.
La Base Moléculaire de la Mutation
Puisque le nucléotide qui est l’unité de
base du codon est formé d’une molécule de base -
purine ou pyrimidine, d’une molécule de
sucre - ribose et désoxyribose, et d’une
molécule de phosphate, celui-là représente
le centre moléculaire de la mutation. En fait, toute modification d’une purine ou d’une
pyrimidine du codon produit une mutation. Ces altérations
peuvent intéresser un ou plusieurs nucléotides (une ou
plusieurs bases) suivant la longueur du segment d’ADN
concerné,
|
|
ALLÈLES NORMAUX
|
ALLÈLES MUTANTS
(délétion sur la 1ère pair et
addition sur la 2ième) |
A
---|---------------- --------------------
---|---------------- -------------------- (1)
B
cistron fait d’unités alléliques en
position cis |
A
; B
---|----------------
---|----------------
-------------------- ------------------- (2)
cistron ou les unités alléliques en position
cis ne sont pas des alleles |
|
|
|
|
|
|
|
|
|
|
ALLÈLES RECOMBINANTS
(recombinaison apres enjambement ou "crossing-over") |
(recombinaison a l’intérieur du cistron
normal)
A
---|---------------- --------------------
----------------|--- -------------------- (3)
; B
cistron fait d’unités alléliques en
position trans |
(recombinaison a l’intérieur du cistron
mutant)
A
---|---------------- --------------------
-------------------- ---|---------------- (4)
; &nbs
p; B
cistron où les unités alléliques en
position trans ne sont pas des allèles |
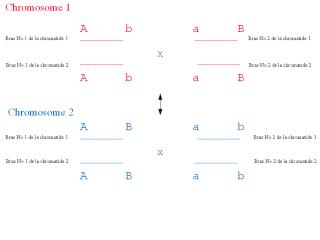
| Fig. 5.20- Mutation
par délétion et addition simples d’un gene (1 et 2)
et mutation par "crossing-over" (3 et 4) |
| * Au cours de
l’enjambement ("crossing-over"), un lien se forme entre les
chromosomes homologues (dans ce cas, un du côté paternel
et un du côté maternel) de la pair, entre les bras des
chromosomes homologues ou, en général, entre leurs
segments d’ADN, au cours de la synapse de la prophase et la
formation de la plaque équatoriale de la métaphase,
durant la méïose. S’ensuit un échange de
segments entre leurs chromatides non soeurs qui conduit à la
translocation des gènes, i.e., à la formation de
recombinants. L’enjambement explique le linkage partiel qui est
une exception aux lois de la ségrégation
indépendante de Mendel. Il est une importante source de
variabilité génétique. |
Quand la mutation n’intéresse
qu’un nucléotide, elle peut être:
- la substitution d’une base par une autre base -
la transition: remplacement d’une purine par une
autre purine ou celui d’une pyrimidine par une pyrimidine ou la
transversion: remplacement d’une purine par une
pyrimidine ou vice versa
- l’addition ou la délétion
d’une, de deux ou plus rarement des trois bases conduisant
à la mutation par changement de cadre qui à
partir de l’altération d’un nucléotide peut
amener celle de plusieurs nucléotides et même de plusieurs
gènes.
Les altérations nucléotidiques
intéressant de larges segments de l’ADN sont
improprement appelées des mutations; elles seraient plutôt du domaine des aberrations
chromosomiques. En effet, les chromosomes sont des zones de condensation ou de surenroulement de l’ADN
qui sont liées à des protéines.
Ces altérations peuvent être:
| |
des changements quantitatifs qui sont:
- la monosomie qui est, chez un organisme diploide, la
délétion d’un chromosome
- la polysomie (alloploidie) qui est l’addition
d’un ou de plusieurs chromosomes
- la polyploidie (allopolyploidie) qui est la
duplication de tout l’ensemble de chromosomes
- des changements de segments de chromosomes:
délétion ou addition d’un bras, par
exemple
|
| |
des changements qualitatifs qui sont dûs à des
réarrangements de l’ADN:
- l’inversion de la séquence des
nucléotides, due à un mode d’enroulement d’un
maillon d’ADN au niveau d’un chromosome
- la translocation de nucléotides ou de
séquences de nucléotides.
|
La majorité des mutations sont
léthales chez les organismes supérieurs ou conduisent
à l’apparition d’individus malades. Cependant, une
fois en passant, une ou quelques mutations peuvent être
bénéfiques. De celles-ci, les duplications paraissent avoir
favorisé l’évolution des organismes vivants. En
effet, des gènes ayant une fonction apparentés sont,
souvent également structurellement proches les uns des autres.
La probabilité que ces gènes soient issus d’un
ancêtre commun, par le processus de duplication, est alors
grande. La raison en est que lorsqu’un gène acquiert une
nouvelle fonction, il le fait le plus souvent au risque de perdre une
ancienne fonction qui était adaptée à son
environnement. Cependant, lorsque
l’organisme "produit" une ou plusieurs copies de ce gène,
elle peut garder l’ancienne fonction et en développer une
nouvelle.
Ainsi, par exemple, peut-on expliquer
1’évolution des organismes haploïdiques et
procaryotiques aux organismes diploïdiques et eucaryotiques.
C’est ainsi, par exemple, que s’explique aussi l’évolution du maïz, une
importante ressource alimentaire des hommes. Chez le maïz moderne,
l’endosperme nourrissant
l’embryon (une fonction importante) est triploïde et les
gènes qui y sont présents sont responsables de la couleur
du fruit. Ordinairement, et comme ceci se produit chez la plupart des
organismes, un seul allèle s’exprime et représente
le gène dominant, tandis que les autres allèles restent
à l’état récessif. Cependant, de temps
à autre (et ceci a été démontré par
Barbara McClintock) un des trois allèles vient à
s’exprimer et devient l’allèle dominant tandis que
les deux autres passent à un état de
récessivité. Le maïz accomplit ceci grâce
à un opérateur activateur (Ac) qui, par
l’intermédiaire d’un promoteur dissociateur
(Ds), active un cistron (ici
réduit à une allèle) et inhibe, en même
temps, les deux autres cistrons, faisant ainsi varier la couleur du
fruit.
Toutefois, chez les organismes supérieurs,
les mutations qui frappent les gènes des cellules autosomes ne
sont pas transmissibles et ne peuvent, dès lors, pas conduire
à l’espéciation. Seules les mutations qui affectent
les gènes des cellules germinales le sont. C’est pourquoi,
chez le maïz, ces gènes qui affectent la couleur de
l’endosperme, mais non l’embyon luimême,
paraissent sautiller (apparaître et disparaître
soudainement) d’une génération à une autre
(d’où leur appelation de gènes sautillants ou "jumping genes,"). Chez les organismes
unicellulaires et chez quelques organismes pluricellulaires qui
n’ont pas la différenciation cellulaire, toutes les
mutations sont transmissibles. Chez la bactérie,
néammoins, lorsque la mutation affecte le plasmide, elle
n’est pas nécessairement transmissible d’un clone
cellulaire à un autre, Cependant, grâce au
mécanisme de conjugaison bactérienne, ceci est possible.
Exercice 10
Soit C, C’ et C’’ les allèles dominants,
présents dans l’endosperme triploïde, qui determine
la variabilité de la couleur, et c, c’ et c’’
les allèles correspondants. Si chacun des trois allèles
est activé préférentiellement et à tour de
rôle, construisez les échiquiers de croisement des
hétérozygotes correspondants et trouvez les rapports de
distribution des phénotypes dans chacun des cas.
LECTURE:
vous avez probablement entendu parler de
l’ingénierie (génie)
génétique. Celle-ci, encore appelée la
technologie d’ADN recombinant, est une méthode
de manipulation du génome des organismes vivants à des
fins médicales (diagnostic et traitement), légales
et agro-économiques. Les outils de l’ingénieur
en génétique sont:
- les ciseaux moléculaires
que sont des enzymes appelés les endonucléases de
restriction, ils permettent de couper l’ADN donneur afin
d’en extraire des gènes
- des colles moléculaires
que sont ces enzymes appelés les ligases d’ADN. Ils
permettent de rattacher des fragments d’ADN coupés par les
endonucléases à d’autres ADNs
- des vecteurs moléculaires
que sont ces ADNs circulaires bactériens appelés
plasmides; le filament d’ADN donneur est introduit dan un
plasmide qui a été ouvert au préalable par
l’endonucléase de restriction; la ligase colle ensuite le
filament d’ADN donneur au plasmide qui se referme; le nouveau
plasmide représente alors un ADN recombinant (d’où
l’appellation de cette technologie de "d’ADN
recombinant").
- des chercheurs
moléculaires que sont des molécules
d’ARN; ils permettent de rechercher le fragment
d’ADN désiré afin de le couper par
l’endonucléase, de le prélever et de
l’exporter ensuite par l’intermédiaire d’un
plasmide; ce processus est celui de l’hybridation de
l’ADN à l’ARN.
Évolution Moléculaire du Prion
Le prion est
la protéine qui code pour la maladie de la vache laitière
ou "mad cow’s disease" qui est responsable de
l’encéphalite spongiforme ches les bovidés ou la
maladie de Creutzfeldt-Jakob, chez les humains. Le prion existe,
normalement, chez les mammifères et protège le cerveau
contre la dégénerescence. La "maladie de la vache
laitière folle" est causée par un handicap de cette
fonction par une forme altérée ou mutante du prion du
bovidé qui vient à s’inclure insidieusement au
poste du prion des primates qui en est très distant, du point de
vue héréditaire, d’où l’appellation en
anglais de "rogue protein", littéralment "protéine
vagabonde et malicieuse", donnée à cette forme mutante
(pour expliquer le mécanisme postraductionel). Le lien au site
ci-dessus vous introduira à l’évolution de cette
molécule. (Réservé à l’auteur).
La figure qui s’y trouve illustre un cladogramme montrant la
proximité et le lien qui existe entre différentes
espèces ("Chart showing prion evolution tree"), basé sur
l’analyse moléculaire. La proximité ou la
parenté est illustrée, a la fois, par la couleur
(même couleur) et l’appartenance à la même
branche du cladogramme. On peut y voir, par exemple, que le gorille
(Gorilla gorilla) et l’homme (Homo sapiens) ont des
prions très apparentés alors qu’eux et un petit singe (Macaca mulattta ou singe
rhezus) ont des prions tres éloignés, étant
distants de 10 souches (branches) parentales. Le prion de Cebus
apella, un singe du nouveau monde, s’est rapproché des
prions des grands singes [tel le gorille, le chimpanzé (Pan
troglodyte) et l’homme], plus proches des singes de
l’Ancien Monde, par un mécanisme, probablement complexe de
mutation, ayant été originellement distant de 7 branches
parentales. Le prion d’un bovidé (Bos taurus) est
également distant de 7 souches de ceux-ci, mais sans
rapprochement mutationel, consécutif. Le prion d’un rongeur (Zitter rat) est beaucoup
plus primitif, n’ayant pas subi de grands changements
évolutifs (espéciation) à partir de la
molécule souche (seulement 2), et est éloigné du
prion des hommes de 8 branches parentales, mais
l’éloignement qualitatif (grande différence entre
les souches parentales) y est encore plus grand..
Le processus de l’ADN recombinant a permis d’introduire
et de cultiver les gènes de
l’insuline humaine chez les bactéries qui servent
dès lors d’usine à production de l’insuline
humaine; celle-ci ne provoque pas de réaction immunitaire
lorsqu’elle est injectée à un receveur
diabétique.
D’autre part, en utilisant des chercheurs
moléculaires, ordinairement des molécules d’ARNm
auxquels ont été attachées un isotope radioactif,
les laboratoires médicaux peuvent maintenant faire un diagnostic médical de précision et
dire, par exemple, si une diarrhée est causée par le
salmonella de la typhoïde, par toute autre salmonella ou par le
staphylocoque doré qui cause l’empoisonnement
alimentaire.
Une autre technique de la technologie de l’ADN
recombinant est employée en criminologie. La plus petite parcelle de tissus
(sang, brin de cheveux ou encore salive) est prélevée de
la scène du crime et de l’ADN, sera comparé aux
ADNs prélevés des individus suspects. Les ADNs
monocaténaires en sont séparés, ensuite, en
utilisant l’électrophorèse sur gel et leur position
sur le gel est comparée à la position de l’ADN
prélevé de la scène du crime, d’où le
nom d’empreinte d’ADN.
La technologie d’ADN recombinant permet de
travailler à l’amélioration
les espèces de plantes et d’animaux qui ont une
importance économique et alimentaire pour les hommes afin
d’en produire de meilleures espèces. Pour ce travail, une
bactérie du sol, l’Agrobacter tumefaciens qui
parasite les plantes et y transfère son plasmide, est
utilisée. Le plasmide de cette bactérie est d’abord
ingénié en utilisant les outils sus-cités et le
gène désiré lui est introduit, Dans un second
temps, la bactérie est cultivée avec une cellule autosome
de la plante et lui transfère son plasmide
ingénié; cette cellule autosome sera à la base
d’un clone de cellules portant le gène
ingénié. Le clone de cellules produites pourra, selon
besoin, être cultivé en une plante adulte qui
présente les caractéristiques désirées:
vigueur et résistance aux maladies des plantes, production
d’une plus grande quantite d’azote ou de protéines,
croissance plus rapide, multiplication plus nombreuse, etc.. Parce que
les plantes se reproduisent aussi asexuellement (pensez à la
greffe, au bouturage et au marcotage qui sont des modes artificiels de
reproduction asexuée, et au grand nombre d’angiospermes
ont la reproduction asexuee, tout en qardant leur capacité de se
reproduire sexuellernent) il est désormais possible
d’ingénier un grand nombre de plantes commerciales et
alimentaires en utilisant la technologie de l’ADN recombinant.
Des plantes telles que celles de l’orange, du maïz, du
café, du tabac, de la tomate, du soya, et de la laitue ont
été déjà ingéniées de cette
façon et beaucoup de céréales, comnme le riz, be
blé ou l’orge, sont maintenant à
l’étude. Pour les animaux, dont le bétail,
l’ingénierie génétique n’est possible
qu’avec bes cellules sexuelles, en particulier les oeufs.
(Rappelez-vous qu’un chercheur suisse a pu cloner des
moutons.)
Enfin, par suite du développemnent de la
technologie de l’ADN recombinant, une nouvelle branche de la
génétique, la génétique à
revers - a pris naissance. Alors que la génétique
classique part du phénotype pour étudier le
génotype, la nouvelle branche de la génétique part
du génotype pour arriver à connaître le
phénotype. Le gène suspect ou en question est
isolé ou préfabriqué et les effete de sa
présence, de son absence ou de sa mutation sur le
phénotype sont étudiés. Les chercheurs nourrissent
maintenant l’espoir que la génétique à
revers sera utilisée pour traiter des maladies
héréditaires, comme la fibrose kystique du
pancréas, la chorée d’Hutington et tant
d’autres.
Parce que l’expérimentation génétique
n’est pas possible chez les humains, les chercheurs ont recours
à des méthodes d’étude indirecte des
gènes humains en vue de soigner et de prévenir les
maladies héréditaires. Ces méthodes indirectes
sont:
- la construction et l’étude de l’arbre
généalogíque à partir des phénotypes
familiaux
- l’étude du caryotype du sujet
- la construction et l’étude de la carte
génétique de l’individu
- l’analyse biochimique
La Construction et l’Étude de l’Arbre
Généa1ogique à partir des Phénotypes
Familiaux
L’arbre généalogique est
la représentation formelle de la présence et de
l’absence d’un trait pour une lignée
familIale. Plusieurs générations sont
représentées sur un arbre généalogique pour
analyser le mode d’héritage du trait - mendélien ou
non mendélien, dominant ou récessif, lié au sexe
ou non - et le sujet chez qui le trait a été tout
d’abord identifié est pointé, d’où son
appellation de propositus ou de cas indice.
Ordinairement, un trait dominant est
présent dans toutes les générations, mais ceci
dépend du nombre d’enfants et du génotype des
parents - l’un pouvant être hétérozygote pour
le trait etl’autre pouvant être normal ou les deux pouvant
être des hétérozygotes (comme nous allons le voir
avec les quelques exernples d’arbres généalogiques,
accompagnés parfois d’échiquiers de croisement, qui
vont suivre). . S’il n’y a qu’un ou deux enfants dans
une génératlon, le trait dominant peut ne pas être
présent dans cette génération, surtout si
l’un des parents est hétérozygote et l’autre
parent normal.
Un trait récessif saute ordinairement les
générations, car, étant donné
qu’il ne se manifeste qu’à l’état
homozygote, les deux allèles qui déterminent le trait
doivent être présents chez le sujet. Il est par
conséquent p1us rare.
Un trait lié au sexe ne se manifeste,
souvent, que pour le sexe masculin, quand il est dominant, car,
pour le sexe féminin, il se produit, ordinairement, un processus
d’inactivation réalisé par l’autre chromosome
normal - l’autre chromosome X - de la pair. Parce
qu’aussi le gène dominant amène des
conséquences plus graves pour le sujet ou des manifestations
plus marquées, il ne se rencontre ordinairement
qu’à l’état hétérozygote
où- un allèle, seulement, détermine le trait,
l’autre allèle étant normal. Un trait
récessif et lié au sexe peut se manifester chez la femme
homozygote, mais il y est beaucoup moins fréquent que chez
l’homme.
Le gène qui détermine le trait peut
être présent chez un individu sans que celui-ci
présente les manifestations de ce trait. Un tel sujet est un
porteur sain. Un trait peut se
manifester, également, à des degrés divers et le
gène est dit avoir une pénétrance complète ou
une pénétrance
incomplète, selon le cas. Ordinairement, un
allèle dominant peut manifester de la pénétrance
incomplète lorsqu’il est présent à
l’état hétérozygote et s’exprimer
complètement à l’état homozygote; un
alléle récessif, par contre, ne peut s’ exprimer,
complètement ou incomplètement, qu’à
l’état homozygote. Enfin, la présence d’un
trait pathogène, chez le foetus, peut conduire à un
avortement ou à un mort-né et rendre plus difficile la
construction de l’arbre généalogique.
Quelques symboles utilisés dans un arbre
généalogique, à titre indicatif, sont:
- pour suivre la transmission des tares
héréditaires sur un diagramme:
- le carré vide pour le mâle qui est normal(§ fig.)
- le cercle vide pour la femelle normale
- le losange pour le sujet de sexe non déterminé
- le carré ou le cercle semi-pleins, suivant le
sexe, pour l’hétérozygote: dans le cas de la
transmission:
- récessive et autosome
- récessive et liée au sexe
(au chromosome X): femelle affectée, seulement
- le carré ou le cercle pleins (coloré en
noir), suivant le sexe, pour le sujet affecté (le degré
d’affectation etant indiqué en pourcentage en dessous du
symbole) dans le cas de la transmission:
- dominante et autosome
- dominante et liée au sexe
- récessive et liée au sexe:
mâle affecté et femelle homozygote,
affectée
- le cercle centré d’un point pour le porteur sain, ordinairement,
une femelle par suite de l’inanactivation du gène par
l’autre chromosome X normal, dans le cas de
l’hérédité liée au sexe
- le carré ou le cercle pleins suivi de la lettre
P, pour le propositus ou
le cas-indice
- pour indiquer les types de liaisons en cause:
- le carré et le cercle reliés d’un
trait simple dans le cas de l’union d’un mâle et d’une femelle non
apparentés
- le carré et le cercle reliés d’un
trait double dans le cas de l’union consanguine
- le carré ou/et le cerle reliés par une
fourchette dans le cas de jumeaux
- pour d’autres raisons:
- le petit cercle pour l’avorté ou le mort-né
- le carré ou le cercle tracés d’une
diagonale pour le sujet
décédé après la naissance
- le point d’interrogation pour indiquer
qu’on doute de la véracité des données
Les arbres généalogiques que nous allons
étudier sont ceux:
- d’un patient souffrant d’achondroplasie
- d’un patient souffrant d’hémophilie
- et d’un patient souffrant d’anémie
falciforme
L’achondropolasie est une
maladie héréditaire, autosome dominante, qui frappe, en
paiticulier, la croissance en longueur des os qui sont, alors,
normalement développés en largeur; ceci etablit la
différencie entre le nain achodronplase et le nain
hypopituitaire chez lequel la croissance du squelette est
diminuée dans toutes les dimensions. Dans les deùx cas,
l’intelligence n’y est pas affectée et le
développement affectif n’est pas affecté par la
maladie (contrairement au nain hypothyroïdien).
L’hémophilie est une maladie
héréditaire grave, liée au sexe, qui est due
à l’absence d’une protéine sanguine qui est
nécessaire à la coagulation normale du sang, le
facteur de coagulation. Un hémophile peut saigner
à mort à la plus petite blessure, mais il existe des
degrés divers de gravité.
L’anémie
falciforme est une maladie à héritage autosome
récessif qui frappe les globules rouges du sang qui, au lieu
d’être ronds, sont en forme de faux, d’où le
nom. Elle peut être plus ou moins grave, d’apres la
pénétrance du gène. . Elle est souvent confondue
avec la thalassémie qui est une anémie beaucoup
plus grave et qui est due au défaut d’une synthèse
normale de l’hémoglobine du sanq. (La personne qui souffre
de la thalassémie a besoin de recevoir périodiquement du
sang, mais habituellement ne vit pas longtemps.) Dans les pays
où la malaria est endémique, l’allêle causant
l’anémie falciforme est fréquent car il
confère une certaine résistance à la malaria.
Exercice:
Soit les arbres généalogiques suivants: (1), (2) et
(3). Analysez les, trouvez le mode d’héritage et dites
à laquelle des trois maladies sus-citées chacun
d’eux correspond. Pouquoí? Construisez les
échiquiers de croisement de la première
génération? Existe-t’il une corrélation
entre l’échiquier de croisement et l’arbre
généalogique? Expliquez.
(1)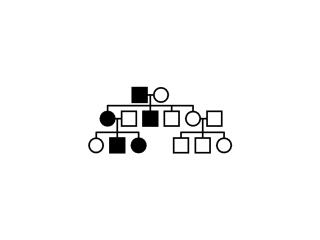 (2) (2)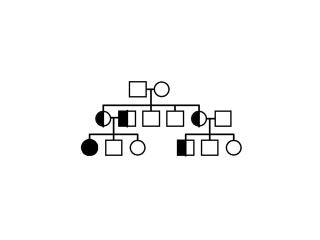 (3) (3)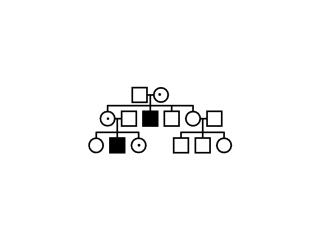
Fig. 5.11, 5.12 et .5.13
Le Caryotype
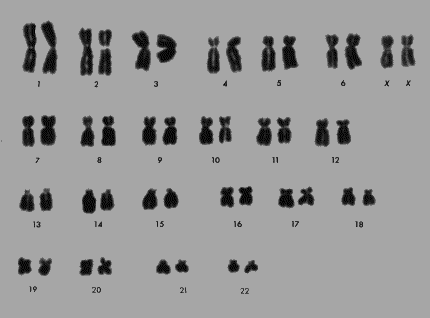 Fig. 5.14 Fig. 5.14
Le caryotype est
l’étude cytologique du génome (i.e. de
l’ensemble des gènes d’un organisme), plus
particulièrement des chromosomes qui le
portent. Les chromosomes sont colorés durant la
métaphase de la mitose, photographiés à un fort
grossissement, puis les clichés sont coupés et
groupés d’après des critères de forme et de
taille. Il existe normalement, chez les humains, vingt-deux pairs de
chromosomes autosomes et une pair de chromosomes sexuels. Les
vingt-trois pairs de chromosomes homologues (ou semblables) sont
rangées par ordre de grandeur décroissante tout
d’abord, puis d’après la position de leur
centromère. Les plus grands chromosomes sont ceux des pairs de
la première rangée du caryotype - 1, 2, 3, 4, 5, et 6 -
pour les autosomes, et XX ou XY, suivant le sexe du sujet, pour la pair
de chromosomes sexuels. Les plus petits des chromosomes sont ceux des
pairs 19, 20, 21 et 22 de la dernière rangée (fig. ). Par
l’étude des caryotypes, les généticiens ont
pu déterminer les causes de quelques maladies
héréditaires, comme la trisomie 21 (où il existe
trois chromosomes 21 au lieu de 2), le syndrome de Turner (où le
génotype sexuel est X0 au lieu d’être xx ou XY), ou
encore le syndrome de Klinfelter (dû à la présence
de deux chromosomes Y à côté du chromosome X
donnant le genotype XXY) . La trisomie 21 (mongolisme) et le syndrome
de Kiinfelter sont assez communs et se rencontrent avec unene
fréquence de 1 sur 500 naissances.
Dans la trisomie 21, le principal facteur est
l’âge de la mère et la maladie est plus
fréquente chez les enfants des mères âgées
(ordinairement ayant en moyenne plus de 35 ou plus de 40 ans).
L’enfant est mal formé, mais sa plus grande
débilité vient de l’idiotie qui accompagne la
maladie. Les enfants atteints de mongolisme sont en
général peu éducables et leur instruction scolaire
est très difficile.
Dans la maladie de Kiinfelter, tout comme dans la
maladie de Turner, un majeur facteur débilitant est la
stérilité des sujets qui en sont atteints. Enfin, ces
maladies sont dues à la non disjonction des chromosomes au cours
de la méiose.
Exercice 12
Faites une photocopie du caryotype présent dans votre livre
et découpez en les chromosomes . Amener les chromosomes ainsi
découpé en salle de classe où des groupes
d’élèves s’amuseront, après leur
repartition au hazard dans une boîte, à les reclasser
d’après leur forme et d’après leur taille.
(Les numéros en dessous des chromosomes seront
enlevés.)
La Carte Génétique
La carte génétique est
la représentation linéaire (sur une ligne) des
différents loci d’un chromome ou d’un segment de
chromosome. (En effet grâce aux travaux de Sturtevant, un
coéquipier du laboratoire de Morgan, il a été
découvert que les gènes peuvent etre representes selon
une disposition linéaire dans le chromosome.) Sur la carte
génétique, cependant, les distances entre les loci ne
sont pas souvent proportionelles aux distances que ceux-ci occupent sur
le chromosome, quoiqu’elles y sont toujours dans le même
ordre.
Sur la carte cytogénétique, les
proportions des distances entre les loci sont conservées sur une
représentation graphique du chromosome. Lorsqu’il
s’agit d’un chromosome sexuel - X ou Y ces loci
représentent des traits liés au sexe.
L’Analyse Biochimique
Un grand nombre d’anomalies héréditaires
peuvent être diagnostiquées par des analyses biochimiques.
De celles-là seront seulement citées la
phénylcétonurie, l’hypothyroïdisme
(crétinisme), la maladie de Tay-Sachs et l’alcaptonurie.
Les enfants souffrant de la
phénylcétonurie ont un retard mental sévère
et restent ordinairement à l’âge mental de deux ans,
leur tonus musculaire est bas et les mouvements corporels sont
désordonnés, leur épiderme et leur chevelure sont
pâles et leurs yeux sont souvent bleux. (Ils peuvent passer pour
de s enfants blonds lorsqu’ils sont de la race blanche et pour
des albinos lorsqu’ils sont de race noire.)
Dans le crétinisme, le niveau des hormones
thyroïdes est très faible, le tonus musculaire est faible,
la croissance est ralentie et le sujet souffle de retard mental.
La maladie de Tay Sachs est fréquente chez
les juifs, probablement par suite de la fréquence des mariages
entre cousins. Elle est due à la carence d’un enzyme qui
est nécessaire à la dégradation d’un lipide
complexe qui s’accumule, alors, dans le tissu nerveux causant le
retard mental, la paralysie, la cécité et une
augmentation de la mortalité infantile.
La reproduction active est la caractéristique de tous les
êtres vivants. Il existe une reproduction asexuée et une
reproduction sexuée.
Le mécanisme de la biologie cellulaire
à la base de la reproduction est la mitose, la division
"augmentationelle" durant laquelle l’ADN nucléaire
se réplique (se duplique) pour donner deux ADN identiques et,
donc, un double assortiment de gènes similaires qui donneront
deux noyaux similaires puis deux cellules similaires et, si possible,
deux organismes similaires.
La reproduction
asexuée est celle des cellules autosomes de notre corps
et, en général, des cellules
autosomes des eucaryotes pluricellulaires. Ordinairement , la
reproduction asexuée est absente chez les eucaryotes
supérieurs. Cependant, la propagation
végétative est une forme de reproduction
asexuée qui est fréquente chez les végétaux
supérieurs et qui est très exploitée en
horticulture (jardinage), étant à la base de certaines
techniques de culture comme le bouturage, le marcotage et le greffage
(§ De la Fougère à l’Arbre du Dr. Roger
Qualo) . En effet, à la différence des animaux où
l’organogenèse n’a lieu que durant le stade
embryonnaire, les plantes conservent, toute leur vie, des centres
d’organogenèse - les méristènes -
formés de tissu embryonnaire. Chez là bactérie, la
reproduction asexuée a un aspect particulier, car elle se fait
sans la mitose caractéristique des eucaryotes, en particulier,
sans la formation du fuseau mitotique et sans la migration des
chromosomes à deux pôles opposés de la cellule;
c’est la fission binaire.
Le mécanisme additionel de biologie cellulaire
qui rend possible la reproduction
sexuée est la méïose, ou la division réductionelle, qui permet la
formation des gamètes et rend posible leur fusion
consécutive (la fusion de leurs noyaux) pour donner un ou
plusieurs organismes portant un assortiment complémentaire de
gènes. Pour beaucoup de gens, la reproduction sexuée est
le synonyme de la copulation, tel que cela ce voit chez les
orqanismes supérieurs, en particulier, chez les plantes et chez
les animaux. Pourtant, la conjugaison bactérienne peut
être considérée comme une forme de reproduction
sexuée durant laquelle une bactérie échange
activement du matériel nucléaire avec une autre
bactérie par l’intermédiaire d’un
prolongement sexuel provisoire - le pilus sexuel - codé
par un gène — le facteur F - situé sur un ADN
bactérién, le plasmide - qui est localisé
en dehors du chromosome unique bactérien (ou de l’ADN
principal bactérien). Deux autres mécanismes de la
reproduction bactérienne qui ressemblent, mais beaucoup moins
que la conjugaison, à la reproduction sexuée sont:
- la transduction
virale où un virus - le bactériophage - sert
d’intermédiaire entre les deux bactéries, la
bactérie donneuse et la bactérie receveuse,
- et la transformation
bactérienne (un mécanisme unique à la
bactérie) où du matériel nucléaire libre
dans l’environnement est adsorbé par la bactérie et
vient à s’incorporer à son matériel
nucléaire. Ces caractéristiques de l’ADN
bactérien sont à la base de la Technologie de l’ADN
recombinant, une des techniques du génie biologique
(bioingénieurie).
Le génie génétique emploie la Technologie de l’ADN Recombinant où un
gène est isolé d’un organisme par l’action
d’un enzyme coupeur, l’endonucléase de
restriction, puis est introduit dans une bactérie où
il sera purifié des gènes avoisinants par la technique
du clonage, puis identifié par la technique
d’hybridization entre l’ARN et l’ADN (grâce
à des marqueurs à ARN complémentaires et
radioactifs) , enfin est “cultivé” en y introduisant l’ADN
de l’organe hôte pour pouvoir reproduire en masse le trait
désiré.
En bioingéneurie (quasi
équivalente, de nos jours, au génie
génétique) aussi, la technique du clonage, que nous avons
déjà rencontrée en étudiant les
bactéries, permet actuellement de maintenir les
caractéristiques avantageuses d’une population
d’organismes, en évitant le brassage
génétique dû à la reproduction
sexuée. Le clone est une
lignée de cellules ou d’organismes qui sont la copie
exacte d’une cellule parentale ou d’un organisme parental.
(Chez nous, par exemple, des clones de lymphocytes du sang ou des
lymphes qui répondent à des antigènes
spécifiques accroissent la capacité de défense
notre organisme (conformément à la Théorie de
la Sélection Clonale.)
Ainsi, pour la canne à sucre et pour
l’ananas, il est possible désormais d’induire
aisément la formation de meilleures plantes (et dès lors
d’augmenter la production) par la technique du bouturage, en
utilisant des échantillons de boutures de plantes clonées
pour leurs meilleures caractéristiques, et en employant des
hormones de croissance bioingéniées. Il a
été possible également d’induire la
reproduction de pommiers “supérieurs”.
Cependant, le clonage réalise le travail inverse de la
biodiversification et présente des risques
appréciables de la propagation rapide des
épidémies dans la population clonée et ceux de la
possibilité d’extinction de la population rendue
homogène (explicables par ce mécanisme de
l’evolution qu’est la sélection naturelle)
L’exemple suivant illustre un tel risque. En Australie,
par exemple, le virus du myxome causant une maladie très
léthale, chez les lapins, a été introduit afin de
réduire la la population de lapins retourna à sa
densité initiale, du fait de la multiplication de souches de
lapins résistantes au virus, par suite du jeu de la
sélection naturelle et de la biodiversification naturelle
à l’intérieur de la population de lapins (due en
partie à la variation génétique aléato:ire
et au choix sélectif du partenaire.) Etait-ce une population de
lapins clonés, ou, du moins, homogènes, le virus du
myxome l’aurait conduite à son extinction.
Du Gamète à la
Progéniture: ontogenèse
Le but de la méiose est de réduire de
moitié le nombre de chromosomes qui seront présents dans
les gamètes, afin de permettre la formation d’un nouvel
individu formé d’un égal nombre de chromosomes
paternels et de chromosomes d’origine maternelle. La
méiose rend la cellule eucaryote haploïde et la
fécondation, i.e., l’union des gamètes, qui
suit, la rend à nouveau diploïde. Le bagage
génétique de chacun des parents est ainsi à
moitié conservé chez sa progéniture. En
même temps, il se produit plus de variétés dans la
population où existe la reproduction sexuée que chez
celle où elle n’existe pas, par suite du brassage de
gènes qui se produit à 1a fécondation.
Supposez, en effet, que votre
arrière-grand-père ("great-grand-father")est A. Votre
grand-père B aura reçu la moitié des gènes
de A; votre père C aura reçu la moitié des
gènes de B et de ceux-ci aura été le quart de
gènes de A; vous, le descendant D,
l’arrière-petit-fils aura reçu la
moitié des qènes de C et de ceux-ci seront
comptés le quart des gènes de B et le
huitième de A. Et ceci est également valable pour
les gènes reçus du côté maternel, i . e., le
nouvel individu aura les gènes A’, B’ et C’
reçus du côté maternel. Quelle grande
diversité! En effet, plus on remonte dans l’arbre
généalogique plus on constate combien la reproduction sexuée introduit de
variété dans la lignée familiale et plus
généralement dans 1’ espèce.
si, par contre, la population se reproduit
asexuellement, (et chez les bactéries sans les
phénomènes de conjuqaison, de transformation ou de
transduction), les individus de toutes les générations
auront leurs gènes en tout point identiques à ceux de
l’arrière-grand-parent A, si l’on exclut les
mutations.
Enfin, chez l’organisme pluricellulaire qui se
reproduit sexuellement, I’ oeuf fécondé de
1’organisme commencera à se subdiviser pour devenir un
amas de cellules qui connaîtront la différenciation
cellulaire, la spécialisation de différents groupes
de cellules et l’histogenèse. Un tel amas constitue
l’embryon formé de trois feuillets de cellules
différenciées: l’entoderme, le
mésoderme et l’ectoderme. A la fin de
l’embryogenèse,
débute l’organogenèse ou la formation des
organes bien distincts tels que le foie, le coeur, les reins, le
pancréas, etc. Néammoins, la reproduction sexuée
n’est pas le seul chemin qui mène à l’évolution. D’autres
mécanismes, que nous avons vus, au chapitre
précédent et au cours de ce chapitre, y conduisent
également, i.e., à la variabilité et à la
différenciation des organismes vivants. "L’ontogenèse récapitule" ou, du
moins, rappelle, d’une certaine manière, "la
phylogenèse", l’évolution des êtres
vivants, rappelons-nous en.
-----------------------
1 Table des coefficients binomiaux (n):
(a + b)n/k
n ; k = 1, 2, 3, 4, 5, 6, 7, 8, etc.
||
1
1 2 1
1 3 3 1
1 4 6 4 1
1 5 10 10 5 1
1 6 15 20 15 6 1
1 7 21 35 35 21 7 1
1 8 28 56 70 56 28 8 1 |

 ADN
ADN