Évolution: Ce Que C’Est et Historique
Sélection NAturelle: Son Rôle dans l’Évolution
Néo-Darwinisme: Ses Rapports avec la Macroévolution & l’Extinction
Évolution & Classification des Êtres Vivants
Preuves Écrasantes de l’Évolution
Évolution de l’Espèce Humaine
L’Évolution & le Déplacement des Continents
-------------------------------------
Sélection NAturelle: Son Rôle dans l’Évolution
Néo-Darwinisme: Ses Rapports avec la Macroévolution & l’Extinction
Évolution & Classification des Êtres Vivants
Preuves Écrasantes de l’Évolution
Évolution de l’Espèce Humaine
L’Évolution & le Déplacement des Continents
-------------------------------------
Évolution: Ce Que C’Est et HistoriqueL’Évolution: Principe Unificateur en BiologieLa théorie de l’évolution, tel qu’elle a été proposée par Charles Darwin, est essentiellement une théorie écologique, avec des évidences tirées directement de la nature, sans aucune référence à la génétique ou à la biologie moléculaire. À l’époque où cette théorie a pris naissance, les travaux de Mendel n’avaient pas encore été découverts et la génétique (qui la confirme) n’avait pas l’essor qu’elle a connu récemment. En effet, Darwin avança sa théorie dans deux bouquins, The Origin of Species et The Descent of Man qui furent publiés, respectivement, en 1849 en 1851. Mendel ne rapporta les résultats de ses expériences avec les pois qu’en 1866, mais ses travaux restèrent inconnus pendant 35 ans et ne furent redécouverts qu’en 1900, soit 50 ans après la publication des textes de Darwin. Par conséquent, pour expliquer l’espéciation et la biodiversité qui en découle, il ne sera fait que peu d’allusion à la génétique, du moins au début de ce cours. La théorie de l’évolution unifie les diverses branches de la biologie qui inclut, entre autres: la biologie de la cellule ou la cytologie, la biologie de l’oeuf ou l’embryologie, la biologie du fonctionnement de l’organisme ou la physiologie, la biologie structurelle ou l’anatomie, la biologie des tissus et des organes ou l’histologie, la biologie du comportement des espèces ou l’éthologie, la biologie de l’environnement ou l’écologie, la biologie des microorganismes qui est la microbiologie, enfin la biologie de l’avènement de la vie dans l’univers qui est du domaine de la biologie moléculaire. Chacune de ces branches a des sous-branches et des domaines apparentés, et la division est, parfois, peu claire entre ceux-ci. La raison est que l’évolution d’un organisme considère et est intimement liée à ces divers aspects de sa biologie.Qu’est l’évolution?La théorie de l’évolution nous dit que les êtres vivants ont eu une origine unique à partir d’un ancêtre commun qui fut la première cellule vivante. Par la suite, au cours de billions d’années de changements dans la structure et dans la physiologie des organismes vivants, les descendants successifs de ce premier être vivant se sont abondamment multipliés, diversifiés et sont devenus de plus en plus complexes pour donner les différents organismes vivants que nous connaissons actuellement et aboutir finalement à l’homme. La théorie de l’évolution s’est opposée au fixisme qui avait nié toute possibilité d’évolution et au créationisme selon lequel tous les êtres vivants auraient été créés par un être supérieur, Dieu, et notre planète serait seulement vieux de quelques milliers d’années. Les preuves de l’évolution sont néammoins abondantes et omniprésentes dans la nature. Il en existe, par exemple, des preuves tirées des prospections géologiques, comme les fossiles, des preuves d’anatomie comparée comme celles qui ont été apportées par les naturalistes, dont Charles Darwin, des preuves embryologiques comme celles de l’anatomie comparative des embryons des différents animaux, et maintenant des preuves génétiques tirées de l’analyse comparative des ADNs. La Théorie de l’évolution avant DarwinLe concept d’évolution suivit relativement de près les débuts de la biologie moderne qui remontent aux environs du 16ième et 17ième siècles. Cependant, à cette époque, la majorité des gens instruits croyaient encore à l’invariabilité des traits hérités des ancêtres et, par conséquent, en l’impossibilité d’un organisme vivant d’avoir des descendants assez différents; c’était l’âge du "fixisme". Pourtant, les 17ième et 18ième siècles furent marqués par une révolution dans la façon dont l’homme concevait la Nature. Dans ces "siècles de la raison", la "révélation biblique" ne représentait plus l’autorité finale à laquelle on se référait pour expliquer les phénomènes naturels, et le théisme avait été largement remplacé par le déisme1a, et même par l’athéisme, dans les cercles intellectuels: le Dieu des interventions miraculeuses et du déterminisme du monde avait été, alors, remplacé par le concept du dieu Créateur des lois générales de l’univers et de l’Origine du monde. Cette interprétation était consistante avec la découverte des grandes lois physiques qui expliquaient le mouvement des planètes sans l’intervention divine. Le philosophe Kant et le mathématicien Laplace qui, entre autres, avaient des idées "rationalistes"1b sur la marche de l’univers avaient été acceptés. A la fin du 18 siècle, il devint clair qu’un problème majeur demandait une explication, celui de l’origine et de la diversité d’un si grand nombre d’espèces d’organismes vivants. Pour un biologiste moderne, le temps qui s’est écoulé entre les premières suggestions provocatrices de l’évolution par Leibniz, dans son Protogaea (1694), et l’hypothèse claire proposée par Lamarck (1800), peut paraître très long. Cependant, Buffon (1707-1788) a été à deux doigts de l’évolutionisme durant toute sa vie et plusieurs autres penseurs ont fait allusion à une échelle de temps dans l’apparition des espèces. Néammoins, aucun d’eux ne s’était, un moment, imaginé, que toutes les espèces aient pu avoir une origine commune. Lamarck (1744-1829), qui commenca ses études de Sciences Naturelles à 49 ans et qui, par la suite, se mit à enseigner celles-ci, pensait jusqu’à 54 ans que les espèces avaient été créées par Dieu et étaient immuables, comme en témoigne son Discours d’Ouverture à l’Académie Française. Cependant, Lamarck avait été influencé aussi par la physique de Newton qui révélait un univers obéissant à des lois naturelles et par les idées de Leibniz. D’autre part, Lamarck, qui était devenu par la suite le directeur de la collection des mollusques du Musée de Paris, avait pu constater que les fossiles de ceux-ci avaient des analogues vivants et montraient un continuum morphologique qui, dans certains cas, remontaient jusqu’à la période tertiaire (comme le révélaient l’examen des couches géologiques) . Lamarck émit alors l’hypothèse qu’avec des transformations graduelles de leur anatomie, les individus d’une population se transformaient, après une longue succession de générations, en des individus assez différents de leurs ancêtres pour être classés dans une nouvelle espèce. L’ensemble de telles espèces, ainsi formées à partir d’une même population initiale et homogène, constitue la lignée phylogénique, le phylum ou l’embranchement. Cuvier, qui, à la même période, avait étudié les fossiles d’animaux terrestres (en particulier des mammifères) qui ne se conservent pas aussi bien que les fossiles marins, n’en avait pas rencontré d’analogues vivants et un continuum morphologique n’était pas tellement évident pour eux. Pour lui, les espèces intermédiaires s’étaient éteintes, i.e., elles avaient disparu à tout jamais de la planète. Enfin, la découverte du fait que les espèces subissaient l’extinction souleva les appréhensions de Lamarck sur l’origine de la vie et donna plus de poids et d’assurance à son hypothèse phylogénique. Cependant, Lamarck n’avait pas l’intention d’écrire une théorie de l’origine des espèces. D’autre part, et ce qui a été plus grave, il se trompa sur le mécanisme de l’adaption qu’il ne sut pas expliquer convenablement. La Sélection Naturelle: Son Rôle dans l’Évolution:La Theorie de l’Évolution de Charles Darwin ou de Darwin-Wallace:La Sélection Naturelle:Dans The Growth of Biological Thought (Le Développement de la Pensée Biologique), l’auteur, vraisemblablement un Lamarckiste, s’est certainement trompé pour avoir écrit que Lamarck a rejetté l’action inductrice directe de l’environnement sur l’organisme comme étant le processus responsable de l’apparition d’une nouvelle espèce. Bien au contraire, l’explication attribuée à Lamarck de l’apparition de la giraffe, cet animal au long cou, n’est que cela. En effet, pour Lamarck, le processus inducteur de l’évolution est environnemental. En d’autres termes ce sont des changements de l’environnement qui provoquent l’évolution d’un organisme à partir d’un organisme préexistant. Ceci est également vrai pour Darwin (1809-1882)(not for internet user), mais alors que Darwin (faute d’une bonne connaissance de la génétique à cette époque) ne chercha pas à expliquer comment ces changements surviennent chez les descendants de l’organisme parental, Lamarck l’expliqua par l’utilisation de l’organe (une explication physiologique). Parce que les transformations morphologiques ou physiologiques amenées par l’usage, ou par le non usage, chez un organisme vivant, ne sont pas héritables, le Lamarckisme a été tourné en dérision. D’autre faits qui ont échappé à l’auteur, Ernst Mayr, du livre sus-cité sont le vrai sens - de l’hypothèse saltatoire ou de l’équilibre ponctué de l’évolution, i.e., de l’évolution par cladogenèse - qui a, principalement, une explication écologique et paléontologique, - et de la différenciation aléatoire (ang., "random differenciation"), i.e., de celle qui survient au hazard, laquelle est, principalement, due aux mutations et a, donc, une explication génétique ou moléculaire de l’évolution par anagenèse2. Ces deux hypothèses ne s’excluent pas d’ailleurs mais se complètent pour expliquer l’évolution des êtres vivants. Avant que nous nous arrêtons sur cette synthèse moderne de la théorie de lévolution (nous y sommes, maintenant, en plein) ou le Néo-Darwinisme, voyons la productivité de Darwin au cours de son voyage, en Amérique, sur le voilier The Beagle. Pour expliquer la diversification des organismes vivants, Darwin admit (sans avoir pu le constater) que des changements surviennent chez les organismes vivants, mais il ne chercha pas à expliquer ceux-là. D’après Darwin, parce que la nature ou l’environnement est éminemment divers et favorise telle ou telle modification sur telle ou telle autre - la sélection naturelle - certains changements sont particulièrement convenables à tel ou tel environnement, et dès lors les espèces qui en héritent y sont particulièrement adaptées; c’est l’adaptation spécifique ou l’espéciation (ang., " speciation"). D’autres modifications, chez les descendants de l’organisme parental, sont moins heureuses, dans tel ou tel environnement, et y sont suivies de l’extinction ou de la disparition du milieu de ces organismes qui en héritent. Darwin, qui avait lu Malthus, traduisit en des termes expressifs - la lutte pour l’existence, i.e., la survie du plus apte - ce mécanisme qui explique la biodiversification (différenciation biologique) survenue ainsi naturellement (par la sélection naturelle) et qui est, à ce stade, directionnelle ou non aléatoire. 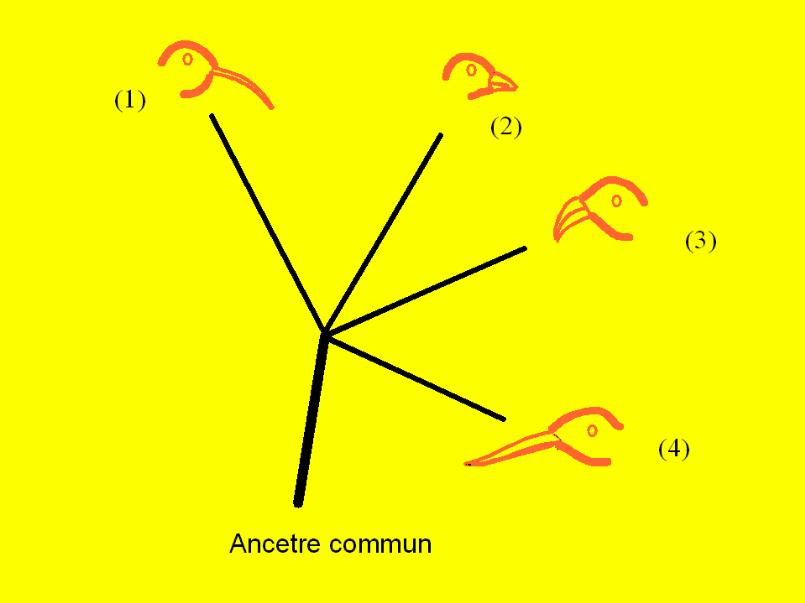 Vous avez vraisemblablement entendu parler des chardonnerets (pinsons) de Darwin (ang., "Darwin’s finches"). Au cours de son voyage de cinq ans aux îles Galapagos, situées dans le Pacifique, face à la cote équatorienne, Darwin eut l’occasion d’observer de tels oiseaux. De ces observations prit naissance sa théorie de l’évolution. En effet, les îles Galapagos sont un archipel de petites îles volcaniques d’apparition relativement récente - près d’un million d’années d’aujourd’hui (comparativement aux 3,5 billions d’années de la planète) - et assez éloignées de la côte continentale, la côte équatorienne, pour empêcher sa colonisation par plusieurs espèces d’animaux et de plantes. Pourtant des 26 espèces d’oiseaux qui dominent la faune aviaire de ces îles, 13 espèces, classées d’après la dimension de leur bec et de leurs habiletés alimentaires, sont des chardonnerets, des oiseaux qui auraient de la difficulté à traverser ces grandes étendues d’eaux. Vous avez vraisemblablement entendu parler des chardonnerets (pinsons) de Darwin (ang., "Darwin’s finches"). Au cours de son voyage de cinq ans aux îles Galapagos, situées dans le Pacifique, face à la cote équatorienne, Darwin eut l’occasion d’observer de tels oiseaux. De ces observations prit naissance sa théorie de l’évolution. En effet, les îles Galapagos sont un archipel de petites îles volcaniques d’apparition relativement récente - près d’un million d’années d’aujourd’hui (comparativement aux 3,5 billions d’années de la planète) - et assez éloignées de la côte continentale, la côte équatorienne, pour empêcher sa colonisation par plusieurs espèces d’animaux et de plantes. Pourtant des 26 espèces d’oiseaux qui dominent la faune aviaire de ces îles, 13 espèces, classées d’après la dimension de leur bec et de leurs habiletés alimentaires, sont des chardonnerets, des oiseaux qui auraient de la difficulté à traverser ces grandes étendues d’eaux. Pour Darwin, un tel phénomène ne peut être expliqué que par la différenciation d’une ou de quelques espèces de chardonnerets qui ont colonisé, les premiers, ces îles; c’est l’évolution horizontale* ou l’ADAPTATION RADIATIVE qui explique le polymorphisme des phénotypes à l’intérieur d’une population et qui est ultimement responsable de l’ESPÉCIATION. À l’opposé de celle-ci, il existe une évolution verticale ou phylogénique3 qui se révèle surtout par l’étude des fossiles- la paléontologie, que nous verrons avec le NÉO-DARWINISME et la MACRO-ÉVOLUTION. D’autre part, l’évolution verticale peut se combiner ou non à l’évolution horizontale pour produire les trois modes majeurs d’évolution: CONVERGENTE, DIVERGENTE et PARALLELE (ďż˝ fig. Arbre Evolutif). L’évolution qu’a défendue Darwin après son voyage aux îles Galapagos est l’évolution divergente ou évolution en rayons de roue appelée encore adaptation radiative (ang., "adaptative radiation". Darwin s’est opposé à l’idée de l’évolution convergente (Darwin Ch.; Origin of Species, p.165, The Modern Library, ed. 1993), parce qu’il défendait son rationalisme qui expliquait l’évolution divergente et l’espéciation dans sa forme la plus simple, i.e, lďż˝apparition naturelle d'une espďż˝ce (d'espèces) dans un environnement donne, à partir de légères altérations dans le dessin du corps d'un ancêtre (commun).. Darwin avait-il raison ou non? Je ne saurais le dire. L��volution convergente n'est pas lďż˝espďż˝ciation à partir d'un ancetre commun. Cependant, l��volution convergente est la cause du "rapprochement" des trait superficiels et communs aux diffďż˝rentes espďż˝ces qui sont prďż˝sentes dans un environnement donnďż˝. L'évolution convergente serait l'espéciation à partir d'ancêtres différents, mais serait toujours le résultat de l'adaptation (à un environnement donné), (comme l'a pensé Darwin pour l'évolution divergente). Une chose est, néammoins, certaine: l'évolution convergente est une logique qui a été plus difficile à entrer dans la première formulation de la théorie de l'évolution qui a été un premier pas vers la compréhension de la phylogenèse (évolution verticale) . (Dialectique du Dr. Roger Qualo). (fig.)... Pour plus de précision sur la nomenclature, concernant cette question référez vous au glossaire Quand des caractéristiques morphologiques semblables, chez des espèces différentes, sont héritées d’un ancêtre commun, appartenant, par exemple, à la même classe, il se crée une homologie. Quand de telles caractéristiques sont héritées d’ancêtres très différents, il se crée une analogie. C’est ainsi que la ressemblance apparente, qui existe entre la forme fuselée de la baleine, de l’Icthyosaure et du requin est un exemple d’analogie du à l’évolution convergente, mais leurs nageoires pectorales antérieures représentent une homologie, leurs lignes divergentes utilisant le même programme génétique, celui qui concerne la formation des membres et du schéma général du corps, ceux-là l’ayant hérité d’un ancêtre commun reptiliforme. D’un autre côté, il existe une dissemblance apparente entre ces deux structures homologues que sont le membre antérieur d’un primate et celui d’une baleine, tous deux des mammifères, produits d’une évolution divergente, alors qu’il existe une ressemblance apparente entre ces deux structures très différentes que sont l’aile d’un oiseau et celle d’un insecte qui est due à une évolution convergente ayant produit cette analogie (fig.) Il a été vu précédemment que l’hétérogénéité spatiale d’un écosystème permet à celui-ci de soutenir plus d’espèces qu’un environnement homogène, en y limitant le processus d’exclusion par la compétition. L’explication du phénomène de biodiversification observé par Darwin est l’hétérogenéité adatative ou la TERRITORIALITÉ. Celle-ci vient du fait que chacune des nouvelles espèces s’adapte et s’attache à des habitats différents qui représentent autant d’aspects singuliers de cet écosystème. :Puisque les îles Galapagos forment un archipel de petites îles relativement isolées par des BARRIÈRES GÉOGRAPHIQUES - des étendues marines, des pressions sélectives distinctes sur ces îles différentes, représentant autant de microhabitats, ont pu conduire, par le mécanisme de la sélection naturelle, à de telles adaptations - adaptations en rayons de roue - et à une ÉVOLUTION DIVERGENTE3 du chardonneret. Tel fût le pressentiment de Darwin après son retour du voyage de cinq années sur le Beagle. Commentaires sur la Théorie de l’Évolution de DarwinLa théorie apportée par Darwin est avant tout celle de la sélection naturelle. En biologie, la SÉLECTION NATURELLE est un fait tout comme l’est, par exemple, l’héritage mendélien des caractères parentaux. Tandis qu’il n’existe pas d’absolu en science, sauf en mathématiques où les postulats sont considérés comme donnés, d’énormes quantités d’informations, supportant la théorie de la sélection naturelle, ont été collectées au cours de ces deux derniers siècles et font de celle-ci un fait indéniable. La sélection naturelle n’est pas l’évolution, et le contraire en est souvent une misconception. L’évolution, telle qu’elle est définie aujourd’hui, est toute modification, dans le temps, des organismes vivants (que ces changements soient ou ne soient pas le résultat de la sélection naturelle) causant:
La sélection naturelle n’est qu’un des mécanismes de l’évolution qui en comporte plusieurs - entre autres (cf. Chapitre de Génétique):
Somme toute, les mécanismes de l’évolution sont à la fois écologiques et génétiques, à la fois environnementaux et moléculaires. Le côté moléculaire s’explique par la capacité de l’ADN (et, en général, de la molécule organique) à former de longs polymères et à donner toute une "infinité" d’intermédiaires, de l’ADN le plus simple (ou le plus petit) des organismes unicellulaires à l’ADN le plus complexe (ou le plus grand) des angiospermes et des mammifères; d’autre part, pour chaque molécule d’ADN il existe de multiples possibilités d’arrangements des unités codantes, ou codons, et donc un grand nombre de gènes possibles et donc de génotypes distincts Le côté environnemental s’explique par la régulation exercée par l’environnement, due à la prédation ou à d’autres épreuves rencontrées dans cet environnement, qui ne permet que la survie des plus aptes et un nombre limité de ces créatures - les phénotypes - de l’ADN et de la chimie organique, et ceci est la SÉLECTION NATURELLE. Darwin a été le pionnier de l’explication environnementale, et Mendel, indirectement, celui de l’explication moléculaire*. Comment se fait cette régulation? Trop souvent, la SÉLECTION NATURELLE est appréciée en termes de la différence du taux de mortalité entre et à l’intérieur de populations différentes, et les individus les plus forts physiquement - les plus capables "athlétiquement" - sont considérés avoir un avantage sur les individus les moins pourvus physiquement. Tel n’est pas souvent le cas, et la sélection naturelle se mesure davantage en termes de capacité de reproduction ou, mieux, par le nombre de gènes laissés dans la population - le puits de gènes - qu’en termes "de capacités athlétiques". Cependant, ce n’est pas tout. En effet, les populations qui peuvent se distribuer dans le plus de niches écologiques possibles - i.e., ayant le plus de capacité d’adaptation - ont également un avantage prodigieux, et, lorsqu’il n’existe pas de barrières reproductives, comme ce l’est souvent à l’intérieur des espèces, l’effet de la sélection naturelle est considérablement amoindrie. (Toutefois, les amérindiens sont presqu’éteints et les néanderthalais - une race d’hommes qui pouvaient sans nul doute se reproduire avec l’Homo sapiens - ont disparu). D’autre part, parce que l’environnement est changeant, les phénotypes qui se distribuent autour de la moyenne (le plus grand nombre d’individus), et qui sont les plus représentatifs de la population, peuvent ne pas pouvoir s’adapter aux nouvelles conditions, alors qu'un phénotype extrême, situés à l’un ou l’autre bout d'une distribution plus large de population, de caracteristiques phenotypiques - i.e., ayant rapport a sa capacité de diversification - pourra, plus facilement, le faire. Cependant, l'inverse est aussi vrai et le tout depend de la direction de ces changements de l'environnement et de leur importance. L’ADAPTATION peut être donc définie comme le "degré de conformité qui existe entre un organisme et son environnement". En d’autres termes, l’adaptation est "l’ensemble des traits, des phénotypes morphologiques, physiologiques ou/et comportementaux, présents chez un individu ou dans une population, qui lui permettent de mieux répondre aux conditions de l’environnement et d’éviter l’exclusion par la compétition". Ces deux définitions précédentes, où l’adaptation est définie par ses résultats, sont néammoins statiques quoique facilement vérifiables. Définie de façon dynamique, l’adaptation est la série de transformations phénotypiques, graduelles qui surviennent dans une population afin de lui permettre de mieux répondre aux conditions nouvelles de l’environnement (ou d’un nouveau environnement) et d’éviter l’extinction ou l’exclusion compétitive. Contrairement à ce qu’avançait Lamarck, ces transformations ne sont pas dues à l’usage puisqu’elles sont héritables, mais elles sont dues à un mécanisme inconnu du temps de Darwin et maintenant connu sous l’appellation de mutations et plus généralement sous celle de transformations génétiques. Ces transformations sont profitables à la population ou aux individus qui en héritent puisqu’elles leur permettent de mieux réussir et de mieux prospérer dans cet environnement et, en fin de compte, d’éliminer ceux qui ne portent pas ces caractéristiques acquises. C’est, donc, ainsi, que l’adaptation a pu conduire aussi à l’espéciation, mais pour cela d’autres conditions ont été necessaires, telles l’isolement géographique et/ou la territorialité qui ont conduit à l’isolement reproductif. Une des preuves de l’adaptation est la CLINE, i.e., la variation graduelle de la fréquence d’un trait intraspécifique dans la population originelle d’une région géographique donnée ou le changement graduel et, donc, intragénérique des phénotypes dans une limite géographique donnée. De tels changements sont caractéristiques chez les indigènes amérindiens de l’Amérique Latine et, en général, de l"Amérique où les individus les plus grands et les plus élancés habitent les plaines et les individus les plus courts, avec la partie supérieure du corps très développée (pour l’adaptation aux altitudes élevées) habitent la cordillère des Andes. Tous seraient vraisemblablement originaires de la même population des premiers hommes qui auraient colonisé les Amériques, en passant par le détroit de Bering, et qui auraient conquis, d’abord, ses plaines et ses régions côtières. L’adaptation à des conditions imprévisibles de l’environnement (tel, par exemple, les effets d’un impact d’astéroide ou de comète) est toujours plus difficile que d’ordinaire. L’adaptation à des conditions très irrégulières, très erratiques de l’environnement peut paraître impossible. Cependant, plusieurs organismes vivants ont développé des phases cycliques et dormantes de vie qui leur permettent de survivre au cours des périodes qui leur sont défavorables. Les modifications de l’environnement qui réduisent l’adaptation de toutes les populations et de de tous les individus qui y vivent sont communément appelées la détérioration de l’environnement. De nos jours, les effets de la pollution crée par les hommes ont conduit à une détérioration marquée de l’environnement qui conduit à l’extinction de toutes les autres espèces. Voilà aussi pourquoi l’espéciation et l’évolution dans la Nature seront de plus en plus difficiles à se faire et à observer. La Nouvelle Synthèse: le Néodarwinisme et la Macro-ÉvolutionDarwin n’a pas lui-même créé le darwinisme. Ses défenseurs et suiveurs - Thomas Huxley et Ersnt Haeckel - en sont les fondateurs. Le darwinisme a assimilé l’évolution au progrès et a contribué ainsi à rendre la théorie de l’évolution plus attrayante face à une opposition forte émanant surtout des croyances religieuses et mythologiques. Cependant, Darwin n’avait jamais assimilé la RADIATION ADAPTATIVE à la MACROÉVOLUTION et à l’ÉVOLUTION VERTICALE. Dans la nouvelle synthèse de la Théorie de l’évolution, celle-ci n’est plus synonyme au progrès et lorsque celui-ci survient au cours de celle-là, ceci n’est que fortuit. En effet, au cours de l’évolution des êtres vivants, des espèces très spécialisées et très avancées, n’ayant pu s’adapter à un changement de l’environnement, ont disparu, ont subi l’EXTINCTION, laissant des espèces moins avancées. Ce n’est que par la suite et ce ne serait que par pur ’hazard’ que de celles-ci est reparti un nouveau processus évolutif. Le Néodarwinisme et la Macro-ÉvolutionLa macro-évolution est tout changement phénotypique d’ordre ’élevé’ qui survient dans une population d’organismes vivants. De tels changements importants dans l’organization d’un organisme se présentent sous la forme d’altérations dans le dessin initial du corps, dans sa structure de base, comme, par exemple:
Que nous apprend à ce propos la paléontologie? Que nous apportent les archives et les collections de fossiles? Pour certains animaux tels, par exemple, le cheval, un genre, et les mammifères, une classe, la lignée de descente est bien conservée, et il n’y a pas eu de difficulté à la retracer. L’évolution d’une forme à une autre de l’animal fut évidente. Le premier cheval, par exemple, était du genre Hyracotherium, un cheval très petit, de la taille d’un chien berger allemand et possédait plusieurs orteils sur un pied (i.e., n’avait pas de sabot). Puis durant 50 millions d’années, une série de changements progressifs ont conduit de celui-ci au Mésohippus, puis à ces chevaux relativement modernes que sont l’Hipparion et l’Hippidion qui ne possédaient qu’un seul orteil par pied, enfin à l’Equus, le cheval moderne. D’un autre côté, les archives paléontologiques montrent, par exemple, une transition graduelle de la forme du reptile à celui du mammifère, avec des animaux qui sont relativement bien placés entre les deux classes et ne peuvent pas être classés dans l’une ou l’autre classe. Cependant, pour un grand nombre d’organismes, la lignée évolutive n’est réellement pas claire. Alors qu’il existe, par exemple, des représentants fossilifères pour tous les groupes majeurs de la classe des échinodermes, tel le concombre de mer, l’étoile de mer, l’oursin de mer (chadron), etc., il n’a pas été possible jusqu’à présent de comprendre la progression de plusieurs espèces. Il existe plusieurs explications à la présence de dossiers vides dans les registres des fossiles:
L’Évolution & l’Extinction:Ce qui donne à la théorie de l’évolution une teinte émotionelle est non seulement ses démêlées avec le créationisme religieux mais aussi le phénomène de l’extinction. Nous pouvons facilement comprendre combien il nous est triste de savoir que notre espèce disparaîtra, un jour, sur terre, sans pouvoir hériter du "paradis céleste" ou de son miroir sur terre--le "paradis terrestre"! Pourtant, le principal artisan de l’extinction est maintenant l’homme dans ses activités quotidiennes: pollution de l’environnement, déboisement, appauvrissement des terres, chasse et pêche intensives. Un exemple bien connu est celui du cheval. Il y a de cela dix milles ans, par exemple, de grandes hordes de chevaux parcouraient les plaines de l"Amérique du Nord. Puis brutalement, ils y disparurent, probablement chassés jusqu’à l’extinction, car le cheval est un animal relativement docile qui se laisse approcher par l’homme qui a pu, par la suite, le dompter. Heureusement, le cheval a pu survivre en Europe où il a pu être domestiqué par les hommes et fut réintroduit, par les colons européens, en Amérique. De nos jours, seuls les animaux qui ont été domestiqués par les hommes et les animaux nuisibles qui suivent ceux-ci ne sont pas menacés d’extinction. Un certain nombre d’espèces d’animaux ont pu, grâce à l’action des environnementalistes et des naturalistes, être enlevés de la liste des animaux en danger d’extinction, mais ceci n’est que provisoire. La croissance même de la population humaine pose un danger à un grand nombre d’autres espèces. Qu’est l’extinction? L’extinction est la perte définitive, dans le patrimoine des organismes vivants, d’une espèce ou de tout groupe phylogénique placé plus haut dans la classification hiérarchique des êtres vivants. Elle est aussi la perte d’un patrimoine génétique. L’extinction n’est pas un phénomène rare. Elle est survenue, plusieurs fois, au cours de l’évolution de la vie sur terre et a frappé des organismes après des millions d’années d’évolution, et elle a touché, à la fois, des animaux et des plantes. Aucun être vivant n’en est exempt. Actuellement cinq grandes périodes d’extinction massive ont été unaniment reconnues par les paléontologistes qui étudient les fossiles:
Les vraies causes de ces extinctions massives ne sont pas connues. L’explication la plus probable est la survenue de changements climatiques globaux dus à la dérive des continents qui a provoqué des modifications des courants marins ou à d’autres phénomènes naturels en rapport avec la géologie de la planète. D’autres causes moins probables ou moins vraisemblables de ces modifications climatiques sont les changements d’inclinaison de l’axe de la planète ou encore, ce qui est actuellement moins probable, quelques évennements plus rares au niveau du système solaire, tel par exemple un impact d’astéroide sur terre. Une extinction qui a fait parler beaucoup d’elle et qui représente un témoignage phénoménal du fait évolutif est celle de ces reptiles géants que furent les dinosaures. Les premiers dinosaures firent leur apparition sur terre, il y a de cela 200 millions d’années auparavant, à une période géologique appelée la Triassique et donnèrent plusieurs groupes d’animaux qui vécurent sur tous les continents durant les millions d’années de cette période consécutive appelée la Jurassique (§ Dr. Qualo, R.; Géologie et Cosmologie, vol. I). À la fin de la période suivante, la Crétacée, soit 130 millions d’années après, les dinosaures étaient amplement diversifiés et représentaient le règne animal dominant parmi les vertébrés. Leurs fossiles furent, en effet, découverts un peu partout, en France au pied du Jura (une chaîne de montagnes qui parcourt l’Europe) en Mongolie (un plateau de l’Asie) en Amérique du Sud et en Amérique du Nord ou fût découvert le fossile du robuste carnivore, le Tyranosaure. Pourtant, les dinosaures s’évanouirent de la terre durant les quelques 10 millions d’années de la fin de la Crétacée - une période assez courte relativement à leur évolution. (Et des reptiles géants, seuls les crocodiles sont encore présents.) Nul ne sait la cause de cette extinction massive. Cependant, ce qui est clair est que leur disparition de la scène terrestre, il y a de cela 65 millions d’années, laissa celle-ci relativement vacante pour le développement d’une nouvelle classe, celle des mammifères, qui était déjà en compétition avec celle des dinosaures. Ceux-là se développèrent rapidement pour devenir la classe dominante des vertébrés sur tous les continents. Une conclusion à en tirer est que l’évolution telle qu’elle est expliquée par Darwin, c. à d., la radiation adaptative, ne se réduit pas à l’exclusion compétitive, mais peut survenir à la suite de crises de l’environnement qui rendent brusquement inaptes des espèces qui étaient auparavant très adaptées à leur environnement. (À bon entendeur, salut!) L’Équilibre Ponctuel. - Quid? La théorie de l’évolution proposée par Darwin, un naturaliste, est gradualiste, i.e., elle explique le processus d’espéciation par des changements phénotypiques graduels dans les populations d’une même espèce. De tels changements aboutiront éventuellement à l’isolement reproductif et à l’espéciation.
Pour nous résumer, disons que l’évolution est comme une grande stratégie de jeu ("a huge game strategy") ayant plusieurs matrices profitables comportant plusieurs entrées variables, *(this page can only be accessed by the author for convenience; the reader can access it by clicking matrices above) tirées de l’hérédité et de l’environnement; l’extinction ou l’espéciation et l’exclusion ou la domination par la compétition peuvent être envisagées comme des sorties après un pari sur telle ou telle entrée. . . . . Les Preuves Écrasantes de l’Évolution.. Les Preuves Tireés de l’Anatomie ComparéeAu début, les phylogénistes utilisaient essentiellement, comme méthodes d’étude des fossiles, l’anatomie comparée et la taxonomie de Linnae. Cependant, depuis peu d’années, l’évolution de la science de la génétique et celle de la biologie moléculaire ont créé une véritable révolution dans la méthode phylogénique: la révolution cladistique4 . Celle-ci est une méthode de classification des êtres vivants qui fait passer au second rang la méthode classique d’anatomie comparée (ou phénétique), connue depuis des années, et qui se base uniquement sur la généalogie (pedigree ou linéage) établie par des méthodes plus précises. Ces nouvelles méthodes - le datage radioactif, la séquenciation protéinique ou nucléique, la distanciation immunologique, le clonage de gènes et l’hybridation de l’ADN- encore appelées la génétique inversée ("reverse genetics"), permettent de déterminer, avec plus de précision, les relations entre les espèces et d’éliminer ou de réduire les erreurs du passé. C’est ainsi qu’en 1970, lorsque des paléontologistes soutenaient que le primate Ramapithèque était un hominidé, un ancêtre direct de l’espèce humaine, l’anthropologue et immunologue Zarich disait qu’il n’en était rien. Se basant sur la séquenciation protéinique et sur la distanciation immunologique, Zarich avait trouvé que la composition des protéines de l’homme et celle de ce singe du Miocène étaient trop différentes pour qu’ils soient considérés comme apparentés, i.e., et que ce singe soit un hominidé . Des découvertes paléontologiques postérieures ont prouvé que l’immunologiste avait raison. Un ensemble de découvertes faites en Chine, durant les années 1980, ont révélé que ce pongidé est un hominoïdé primitif dont le clade (la branche) s’est bifurqué du tronc principal des anthropoïdés a une époque beaucoup plus antérieure que ne l’ont fait les hommes et les grands singes actuels, en d’autres termes que cet hominoïdé est beaucoup plus éloigné des hommes que ne le sont les grands singes, en particulier le gorille et le chimpanzé . Les Preuves Tirées de la Biologie Moléculaire Les dernières preuves de l’évolution ont apportées avec force par la biologie moléculaire, en particulier par la technologie de la greffe du matériel génétique. Il est maintenant, en effet, possible d induire expérimentalement le passage d’une espèce une autre. L’ADN hybride utilisé dans ce cas est appelé ADN recombinant. La technologie en cause est celle de l’ADN recombinant, encore appelée le génie génétique, une spécialité du génie biologique. Les outils de l’ingénieur généticien sont: Les dernières preuves de l’évolution ont apportées avec force par la biologie moléculaire, en particulier par la technologie de la greffe du matériel génétique. Il est maintenant, en effet, possible d induire expérimentalement le passage d’une espèce une autre. L’ADN hybride utilisé dans ce cas est appelé ADN recombinant. La technologie en cause est celle de l’ADN recombinant, encore appelée le génie génétique, une spécialité du génie biologique. Les outils de l’ingénieur généticien sont:
Les implications de la technologie d’ADN sont nombreuses, extraordinaires et sérieurses quoique les succès soient actuellement très limités. Par exemple, des espèces peuvent être améliorées et amenées à s’adapter à des conditions auxquelles elles ne s’adaptaient pas auparavant. Les Preuves Paléontologiques de l’ÉvolutionLa paléontologie ressemble à l’archéologie et dès lors à l’histoire. Dans certaines universités, elle est même étudiée comme une branche de celle-là; l’archéologie nous permet de connaître les civilisations passées. Dans le dictionnaire anglais, le Unabridged Grand Webster International, l’archéologie est définie comme étant l’"étude scientifique des civilisations éteintes ("extinct") ou celle des phases antérieures de la culture d’un peuple à travers les fossiles laissés et les objets créés par celui-ci dans le passé, tels les squelettes, pour les premiers, les monuments, les outils, les artefacts, l’écriture, et les inscriptions, pour les seconds." La paléontologie est en rapport avec la géologie; elle est cette science qui étudie les fossiles des organismes vivants et apporte des données sur la chronologie des diverses phases de l’évolution de la vie sur Terre et sur ces rapports avec l’évolution du globe. Dès lors, la paléontologie a été d’une grande importance à la nouvelle théorie scientifique, celle de l’évolution. L’étude des fossiles d’espèces disparues a permis aux hommes de se rendre compte que des espèces d’être vivants primitives et éteintes étaient dominantes sur terre dans le passé lointain, alors qu’un grand nombre d’espèces actuelles ne sont apparues sur Terre que dans des périodes relativement récentes. Souvent, des formes intermédiaires en ont été trouvées, ce qui a été une preuve formidable en faveur de l’évolution. Présentement, les biologistes et les paléontologistes, dans leurs tentatives de reconstruire l’histoire de l’évolution de la vie sur terre, peuvent le faire dans des intervalles raisonnables de précision, en s’aidant des méthodes utilisées par les sciences expérimentales. Une de ces méthodes, et non des moins importantes, est le datage radioactif (qui nous vient des Sciences Physiques). Le principe du datage radioactif est l’utilisation des isotopes radioactifs comme des horloges radioactives. Trois points importants y sont à retenirr :
Parce que le temps de désintégration de C14 est relativement court, la quantité qui en reste dans un fossile après environ 40.000 ans devient trop faible pour être détectable par le compteur, et il s’avère nécessaire d’utiliser des isotopes qui se dégradent plus lentement tels l’uranium 235 et le plomb 207 (U235/Pb207) (celui-là donnant celui-ci) dont le t1/2 est 713 millions d’années,le potassium et l’argon 40 (K40/Ar40) dont le t1/2 est 1300 millions d’années, l’uranium 238 dont le t1/2 est 4510 millions d’années et le rubidium 87 dont le t1/2 est 47000 millions d’années. Une fois que les âges d’un groupe de fossiles aient été établis pour un organisme, il devient possible d’en construire une phylogénie, i.e., une description de la ligne de descente, du linéage, de cet organisme en passant d’une époque, d’une période ou d’une ère géologique à une autre. Enfin, en s’aidant du datage radiaoctif, les paléontologistes pensent avoir établi l’âge de quelques fossiles avec une précision de 98%; ce qui est plus que raisonnable! Les Preuves Tirées de l’Embryologie et de l’OntogénieL’embryologie a fourni à Darwin l’un des meilleurs arguments contre le Créationisme scientifique. Si, en effet, les espèces avaient été créées, pourquoi les embryons de divers ordres d’animaux d’une même classe présentent-ils un stade antérieur durant lequel ils se ressemblent tous? Pourquoi, par exemple, des branchies sont-elles présentes chez tous les embryons des vertébrés? Pourquoi à, un stade ultérieur, ces embryons diffèrent-ils? C’est précisément qu’au cours de la phylogenèse des variations se sont introduites à partir d’ancêtres communs pour produire les différences observées, et c’est ce que nous apprend l’ontogenèse. Cependant, les preuves embryogéniques et ontogéniques5 restèrent longtemps controversées, par suite de la dialectique ombrageuse, niaise et stérile qui opposa les récapitulationistes (école d’Haeckel et de Muller) et les fixistes (école de Von Baer) (réservé à l'auteur). Ceux-là affirmaient que l’ontogenèse récapitule la phylogenèse et que l’embryon des animaux récents (en particulier celui de l’Homme) passe par différents stades morphologiques qui rappellent les formes adultes des espèces inférieures, i.e., des espèces qui leur ont précédé dans la phylogenèse. Le contre-argument apporté par Von Baer est que des structures primitives, telles que la notochorde et les poches branchiales rencontrées au cours de l’embryogenèse des vertébrés (cf. Dr. Qualo R., dans De l’Étoile de Mer à la Lamproie & Du Poisson au Mammifère), sont remarquablement absentes chez les adultes de la plupart des ordres antérieurs, et que les ordres successifs (ayant apparu par la suite) ne peuvent pas dès lors avoir évolué ou divergé à partir d’un ancêtre commun; ce qui s’est produit, d’après lui, n’est qu’une différenciation du général au particulier, chez tous ces animaux, et ceci ne laisse en rien penser qu’il y a eu une évolution de la morphogenèse, une évolution des formes simples vers des formes plus complexes, laquelle serait ensuite héritée et se retrouverait condensée au cours de l’embryogenèse des organismes supérieurs. La découverte des lois de MENDEL mit fin aux allégences des récapitulationistes (ainsi qu’à celle des Lamarckistes), qui avaient avancé la possibilité d’une évolution, par l’addition terminale des variations phénotypiques et par leur condensation subséquente, laquelle se manifesterait au cours de l’embryogenèse des formes supérieures. En effet, ces lois montrent que:
Les lois, formulées plus tard par MORGAN révélèrent que:
Ceci mit fin aux allégences des fixistes et prouva que l’évolution est possible à partir de formes antérieures, la sélection naturelle faisant le triage des phénotypes ainsi produits. Il est maintenant admis par tous les protagonistes de l’évolution que le parallélisme entre l’ontogenèse et la phylogenèse n’est valable que pour les embryons des divers groupes (classes, ordres, familles, genres et espèces) d’un cladogramme mais est inapplicable à la comparaison entre les formes embryonnaires et les formes adultes de celui-ci. L’Évolution HumaineLes hommes sont des hominoïdés. Cette superfamille comprend les hommes, les grands singes et les gibbons. Les gibbons sont de la famille des hylobatidés. Les grands singes dont les plus modernes sont le gorille, l’orang-outan et le chimpanzé sont de la famille des pongidés (ancienne classification). Les hommes actuels et les hommes préhistoriques sont des hominidés. Les hominoïdés forment avec les petits singes le sous-ordre des anthropoidés, lequel constitue avec celui des prosimiens - lémur,tarsier et loris - l’ordre des primates. Le premier HOMINOÏDÉ est apparu sur terre il y a de cela 29 à 15 millions d’années auparavant avec l’Aegyptopithèque, vraisemblablement descendant d’un ancêtre commun avec le gibbon. De ceux-ci évolua le Dryopithèque dont le représentant africain, le Proconsul africanus, est généralement désigné comme l’ancêtre commun à l’hominidé et au pongidé (grands singes). Du Proconsul prit naissance le Ramapithèque/Sivapitèque qui donna l’orang-outan et l’Africopithèque. Ce dernier donna deux branches, celui des grands singes et celui de l’Australopithèque, il y a de cela 3.0 à 2.0 millions d’années auparavant. L’Australopithèque dont le nom veut dire non pas un grand singe australien , mais un grand singe du sud de l’Afrique représente réellement le premier HOMINIDE. En effet, bien que le volume du cerveau de l’Australopithèque africanus ait été pareil à celui des grands singes - 450 à 550cm3, le quotient d’encéphalization, qui est le rapport du volume du cerveau à celui du corps, de l’Australopithèque est deux fois plus grand que celui des grands singes et est beaucoup plus proche de celui des hommes. Un trait important de la dentition de l’Australopithèque qui le fait classer aussi comme un hominidé est que la partie extra-gingivale de la caninne n’est pas plus longue que celle des autres dents comme ceci est chez les singes. Dans la famille des hominidés, deux genres sont representés,comme nous le révèle l’étude des fossiles: le genre Australopithèque et le genre Homo, comportant, chacun, plusieurs espèces. Pour le genre Australopithèque, celles-ci furent: A. afarensis, A. africanus et A. robustus (A. boisei). Pour le genre Homo, les différentes espèces, tel qu’il en découle de l’ étude des fossiles sont: H. habilis, H. erectus (encore appelé Homme de Pékin, Homme de Java ou Pithécantropithèque) et H. sapiens. De ce dernier prit naisance trois sous-espèces: H. sapiens primitif, H. sapiens néanderthalensis (Homme de Néanderthale), et H. sapiens sapiens, cette dernière sous-espèce, représentant l’homme moderne, étant encore la seule vivante. D’autre part, une forme de transition - l’Homme de Cromagnon - a été trouvée entre l’Homme de Néanderthale et l’homme moderne. Enfin, un nouveau cladogramme qi se base sur les évidences apportées par la biologie moléculaire place le chimpanzé et le gorille dans la famille des hominidés parcequ’il s’est révélé que les grands singes africains sont plus proches génétiquement des hommes qu’ils ne le sont de l’orang-outan (le grand singe asiatique de Bornéo et de Sumatra!!!). La table et le schéma qui suivent résument et rendent plus claire ce que nous venons de dire: Classification des primates
 -------------------------------------------------------------------------------> Sens de l’évolution Les grandes caractéristiques du genre Homo et, en général, des primates supérieurs sont:
Certaines de ces caractéristiques se retrouvent également chez d’autres primates et ne sont pas, dès lors, l’exclusivité du genre, encore moins de l’espèce. L’Évolution & le Déplacement des ContinentsUne question clef au sujet de la macro-évolution est de se demander si une variation génétique radicale est responsable de ces changements phénotypiques prononcés qui existent entre les majeurs groupes taxonomiques, tel, par exemple, entre les classes et entre les embranchements. En effet, dans un environnement stable, de telles modifications majeures du phénotype peuvent ne pas conduire à des individus viables par l’action de la sélection naturelle. Cependant, lorsque celles-ci coïncident "par chance" (de façon aléatoire) avec un changement "catastrophique" ou majeur de l’environnement, de tels phénotypes deviennent des "monstres désirables", et leur survenue représente une "ponctuation", une "singularité" dans l’histoire de la vie sur terre. Cependant, les mêmes mécanismes qui sont à la base de la microévolution expliquent aussi bien la macroévolution. Ceux-ci sont:
Immédiatement après la découverte des travaux de Mendel, les théories qui expliquent les deux modes d’évolution - la microévolution et la macroévolution - étaient incompatibles, et le Mendélisme s’opposait au Darwinisme. Cependant, grâce, et surtout, aux travaux de Morgan, la génétique fut amenée à reconnaître que la plupart des caractéristiques phénotypiques sont contrôlées par plus d’un gène et que l’expression de celui-ci peut être plus ou moins régularisé. Dès lors, il est devenu plus facile d’admettre que ces mêmes mécanismes qui sont à la base de la microévolution peuvent déboucher, dans d’autres cas, sur la macroévolution. Que certains le veuillent ou non, la grande réalité est que les dinosaures ont existé. Bien que plusieurs hypothèses circulent sur la cause probable de leur extinction, il est fort probable que des changements écologiques majeurs, produits par un phénomène ordinaire, comme, par exemple, le déplacement des continents--une hypothèse alternative à celle d’Alvares ou de celle de l’impact d’un grand astéroide, puissent l’avoir causée. À la période dévonienne, par exemple (il y a de celà 380 à 400 millions d’années) les continents étaient très éloignés les uns des autres et dans une position excentrique sur le globe terrestre. Le climat était probablement glacial sur les continents d’antan, comme en témoignent les empreintes laissées par les glaciers qui furent au nombre de quatre (4) durant l’ère précambrienne ou protézoique(2500 millions d’années auparavant) de l’éon cryptozoique, de deux(2) durant la période ordovicienne (500 millions d’années auparavant) et d’une (1) durant la période carbonifère (300millions d’années auparavant) de l’ère paléozoique de l’éon phanérozoique (dans laquelle nous nous trouvons toujours). Puis est venue la période permienne (280 millions d’années auparavant) qui s’est caracterisée par la fusion des continents en une masse centrale et en grande partie équatorienne- Pangea. Dans les régions intérieures et éloignées des poles de ce supercontinent, il a du faire très chaud du fait de leur inaccessibilité relative à l’effet adoucissant des courants marins et de la brise océanique, bref un climat propice aux animaux à sang froid, tel que les reptiles par exemple. Les dinosaures16 ont pu se développer au cours de cette période; ils étaient les bienvenus car le climat leur était propice. Par la suite, la dérive des continents dans un sens opposé amena l’éloignement de ceux-ci et explique le fait que des fossiles de dinosaures soient trouvés dans des régions aussi éloignées et séparées par les océans que sont l’Europe de l’Ouest, au pied du Jura, la Mongolie, en Asie, les Amériques. Avec ce déplacement des continents vers les régions polaires, (fig. ) les climats continentaux se sont considérablement adoucis et refroidis, créant un temps propice au développement de ces animaux à sang chaud que sont les mammifères et à l’extinction des grands reptiles incapables d’engendrer la chaleur corporelle nécessaire à leur survie (à l’eclosion de leurs oeufs fort probablement). Quid du volcanisme?Les dinosaures vivaient-ils à une période d’intense volcanisme continental? Fort probablement! Cependant, l’effet d’écran à la lumière solaire, produit par les cendres volcaniques, n’a pas été suffisant pour provoquer un refroidissement significatif du globe; il faudrait aussi compter avec l’effet de la maison de serre ("Greenhouse effect"), produit par le gaz carbonique volcanique, allant à l’encontre d’un refroidissement continental. La température du globe aurait même augmenté de 0o à 20oC durant l’ère mézozoique, dans la période immédiate après la fracture du Pangea, créant un climat propice à l’eclosion des reptiles mais pour atteindre des niveaux insupportables aux grands animaux, à la fin de la Crétacée, puis chuter brutalement avec l’élargissement de l’Atlantique et la formation de nouveaux courants marins, (tenant compte des décalages entre les phénomènes géologiques et les variations climatiques). (§ Cesar E.; Planet Earth: Cosmology, Geology, and the Evolution of Life and the Environment).ConclusionPour clore, répétons que l’évolution est comme une grande stratégie de jeu ayant plusieurs matrices profitables comportant plusieurs entrées variables (tirées de l’hérédité et de l’environnement); l’extinction, l’espéciation et l’exclusion ou la domination par la compétition peuvent être envisagées comme des sorties après un "pari", un jeu sur telle ou telle entrée. Cependant, même cette définition paraît un peu trop simple, car l’évolution a une directionalité et ne fait pas marche arrière. En effet, les organismes eucaryotes sont plus évolués que les procaryotes, les pluricellulaires le sont davantage que les unicellulaires, les vertébrés le sont encore plus que les invertébrés et les mammifères le sont davantage que les reptiles; et leur apparition au cours de l histoire de la terre s’est faite dans un ordre chronologique. Peut-être que la science actuelle aidée de la nouvelle mathématique du CHAOS et de l’étude des boucles d’itérations pourrait nous aider un jour à mieux la saisir, car l’évolution porte sur des systèmes dynamiques en perpétuel changement.
| ||||||||