Un chapitre sur l’intégration des systèmes biologiques est important comme nous le fait voir le livre de Marie D. Hoff, Sustainable Community Development, car ultimement nous serons amenés à parler, ou du moins à envisager, des moyens pour notre communauté d’atteindre l’autosuffisance dans l’environnement dans lequel il évolue.
Malgré la grande diversité de la vie, il s’y rencontre une impressionant degré d’organisation et d’ordre. Ceci n’a cependant été possible qu’avec une extraordinaire interdépendance fonctionelle et de nombreuses interelations à tous les niveaux du phénomène biologique - de la cellule à l'organisme et de celui-ci à la communauté. Cette tendance à l'organisation et à l'intégration qui caractérise les sytèmes biologiques donne naissance à des mécanismes de régulation, à des actions et à des réactions à toutes les étapes du phénomène "vie" et contribue ainsi à la stabilité de celle-ci sur notre planète. Pour un organisme vivant (considéré seul) une telle stabilité due à l'autorégulation de ses fonctions vitales est l'HOMÉOSTASIE. Celle-ci implique l'existence de boucles de rétroaction qui permettent à un système qui sort de ses conditions de normalité d'y revenir de lui-même.
Chez le mammifère, par exemple, le sucre est un "métabolite" (métabolisant) important. Lorsque le niveau de celui-ci baisse ou s'élève trop pour une raison quelconque, telle par exemple, un jeûne prolongé ou un repas trop copieux1, des hormones - respectivement l'adrénaline et l'insuline - sont déchargées dans le sang pour ramener la concentration du sucre - la glycémie - à des valeurs normales. De tels mécanismes d'autorégulation existent également dans un écosystème dans les diverses communautés de plantes et d'animaux et, à un certain degré, pour les divers paramètres environnementaux, justifiant l'expression populaire d'"équilibre naturel". En effet, c'est en combinant ceux-ci de façon remarquable qu'un écosytème arrive à engendrer et à soutenir un flux d'énergie qui circule de la base au sommet de sa pyramide écologique, en passant à travers les diverses couches de populations qui le composent. Enfin, de l'intégration des divers écosystèmes terrestres dépend la biosphère.
.
.
.
2. Coordination d'un Organisme
Tous les organismes vivants et leurs entourages non vivants sont faits d'éléments chimiques, i.e., d'atomes et de particules sub-atomiques qui peuvent être déplacés par des "quanta d'énergie". La vie, elle-même, un phénomene dont l'origine est à peine comprise par la science actuelle, serait apparue sur terre à la suite d'un nombre indéterminé de combinaisons et de réactions physico-chimiques entre les divers élements rencontrés sur notre planète.
Durant une période de l'évolution de celle-ci qui s'étendit sur des millions d'années, ces combinaisons conduisirent à des composés de plus en plus complexes et de plus en plus organisés et intégrés, par une série de processus naturels, les uns jusqu'à présent inconnus, les autres (et de loin, le plus grand nombre) partiellement compris (et ceci sans aucune intervention divine!). C'est ainsi que s'explique la théorie atomistique qui est à la base de la biologie moderne et qui compare la cellule vivante à une usine de réactions chimiques.
Les unités fonctionelles et les unités structurelles de tout organisme vivant sont intimement intégrées et regularisées en fonction des demandes imposées par l'environnment. Chez les animaux, deux modes majeurs d'intégration sont le système nerveux et le système endocrinien. Le premier réalise cette intégration qui permet une communication rapide rapide entre les diverses parties du corps et entre celui-ci et l'environnement extérieur. Le deuxième est plus lent quoique plus durable dans cette même action. En fait, des investigations scientifiques ont montré qu'il n'y a pas de grande différence entre les deux, et rappelez-vous que l'hypothalamus est, à la fois, une glande endocrinienne et une structure cérébrale. (Les systèmes nerveux et endocrinien ont été vus en classses terminales avec mon texte intitulé L'Homme, ce Primate Supérieur .) Dès lors, je m'attellerai ici sur l'intégration biochimique ou métabolique (pour me rapprocher du cours d'Anglade) mais, avant, je voudrais situer celle-ci dans l'échelle de grandeur des processus naturels qui suit.
Cycles Biochimiques
(ex.: cycle du Carbone ou cycle de Calvin-Benson et cycle de Krebs)
|
Cycles Cellulaires
(ex.: incluant la mitose et l'interphase d'une cellule)
|
Cycles de Vie
(ex.: la reproduction d'un organisme, par ex., celle d'un trématode)
|
Cycles écologiqes
(ex.: cyle de production et de décomposition de la matière organique
incluant les producteurs, les différents stades de consommateurs et les détrivores ou décomposeurs)
|
Cycles Géologiques
(ex: la capture de l'énergie solaire par notre planète,
formée de l'atmosphère, de l'hydrosphère, de la lithosphère et de la biosphère, et son recyclage)
|
Cycles Cosmiques?
(avec la formation et la destruction de mondes: soleils, galaxies, etc.)
|
CYCLES BIOCHIMIQUES ET PROCESSUS CELLULAIRES D'INTEGRATION
La structure de la cellule est essentiellement dépendante de sa fonction qui est d'assurer la vie et qui peut se résumer a deux modalités fonctionelles: le métabolisme et la reproduction. Tous deux réclament de l'énergie pour se réaliser, une énergie qui, en fin de compte, provient du soleil et qui est transformée dans la cellule de la feuille, tout d'abord, en énergie chimique.
Le Métabolisme
Le métabolisme est le processus au cours duquel des molécules de graisse (lipide), de protéine (protide) et de sucre (carbohydrate ou glucide) sont utilisées à des fins énergétiques - le catabolisme - et à celui de la transformation d'une partie de cette énergie pour des activités de synthèse - l'anabolisme. Trois importants processus métaboliques sont l'oxydation l'oxydation (déshydrogénation), la réduction (hydrogénation ou addition d'hydrogène) et la carboxylation (addition de carbone). Trois grandes voies métaboliques sont, par exemple:
- le catabolisme des sucres, la principale source d'energie de l'organisme vivant, dont la première étape est la glycolyse,
- l'anabolisme musculaire qui conduit à la formation des plus grandes molécules (macromolécules) de l'organisme, les protéines musculaires,
- et la photosynthèse qui est la première étape dans la formation des molécules organiques chez la plante et, en fait, leur synthèse de novo à partir des molécules inorganiques.
Le Catabolisme des Sucres
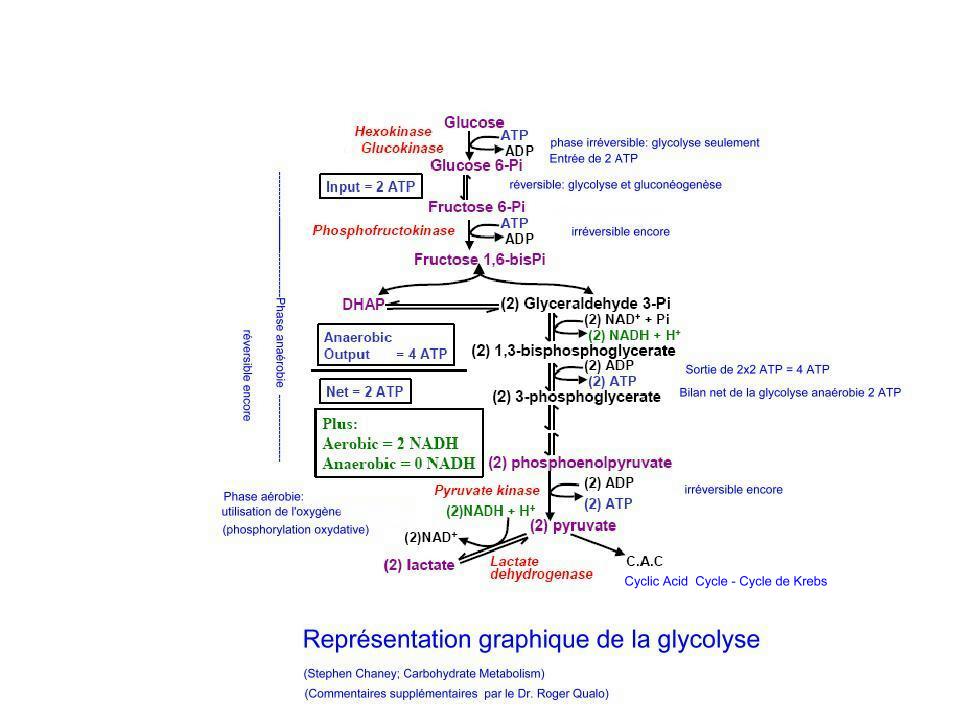
Le catabolisme des sucres peut être divisé en deux étapes: une étape anaérobie ou glycolyse et une étape aérobie ou d'oxydation cellulaire qui inclut le cycle de Krebs et la chaîne des transporteurs des électrons ou chaîne des phosphorylations oxydatives.
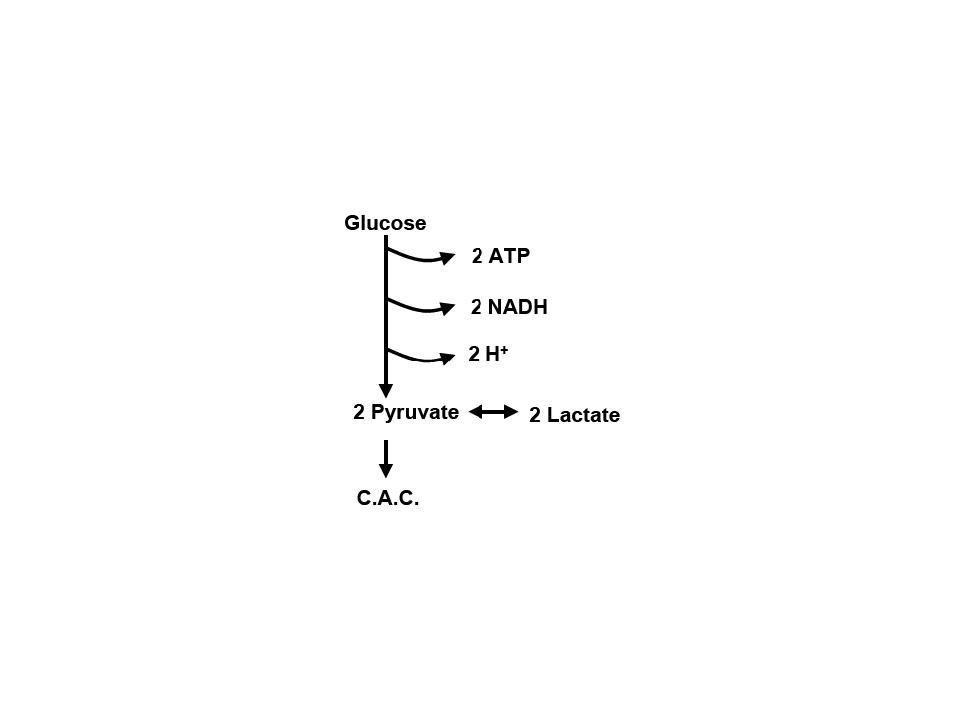 | | Dr. Chancy, G. S.; Carbohydrate metabolism, (966-3586); med.unc.edu |
� LA GLYCOLYSE
Dans la glycolyse, la dégradation du glucose en acide pyruvique ne produit qu'une très faible quantité d'ATP, la forme d'énergie qui est utlisée par la cellule (et qui provient de l'oxydation de ces carburants biologiques que sont les sucres et les graisses de notre organisme). Au cours de la glycolyse, le glucose, un hexose, i.e., un sucre a six atomes de carbone (6C), est progressivement transformé en un sucre à trois atomes de carbone, un triose. Parce que le glucose est phsophorylé au cours de la glycolyse le triose obtenu sera le triose phosphate, conformément à une série de réactions mises en évidence en cliquant sur le schéma ci-contre (et qui demandent une dépense d'énergie initiale). Le bilan de la glycolyse est positif pour la production de l'énergie cellulaire, l'ATP, avec un résultat net de deux ATPs, car 2 ATPs ont été utilisés au cours des trois premières étapes et, au cours des deux dernières etapes, 4 ATP ont été formés. Sur l'un des clichés du catabolisme des sucres, il est mention de gluconéogenèse; celle-ci ne représente pas une voie catabolique mais y est seulement représentée pour la complémentation; la gluconéogenèse (réservé à l'auteur)est le chemin inverse de la glycolyse qui est la voie normale du catabolisme des sucres, (la voie des pentoses phospahtes, une autre voie, y représentant une sorte de dérivation, un "shunt").
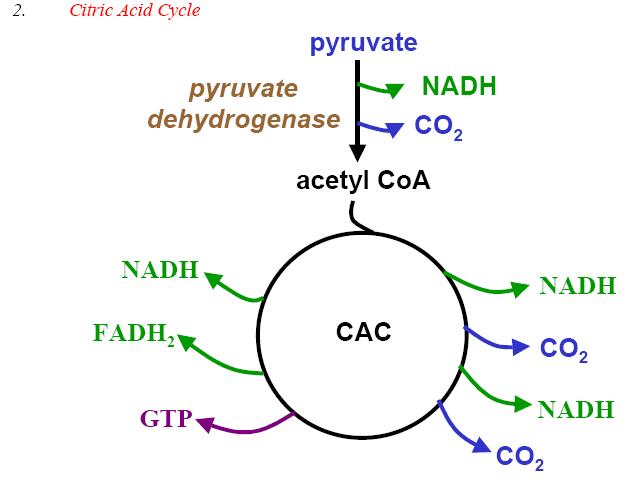 | | Dr. Chancy, G. S.; Carbohydrate metabolism, (966-3586); med.unc.edu |
� Le Cycle de KREBS (cliquez dessus pour voir les détails, puis cliquez à l'apparition de la figure plus large: "view" puis "full screen"; vous pouvez ensuite rouler dessus ("roll over") pour voir davantage de détails).
Le cycle de Krebs a été vu également dans le premier chapitre de Physiologie Végétale.
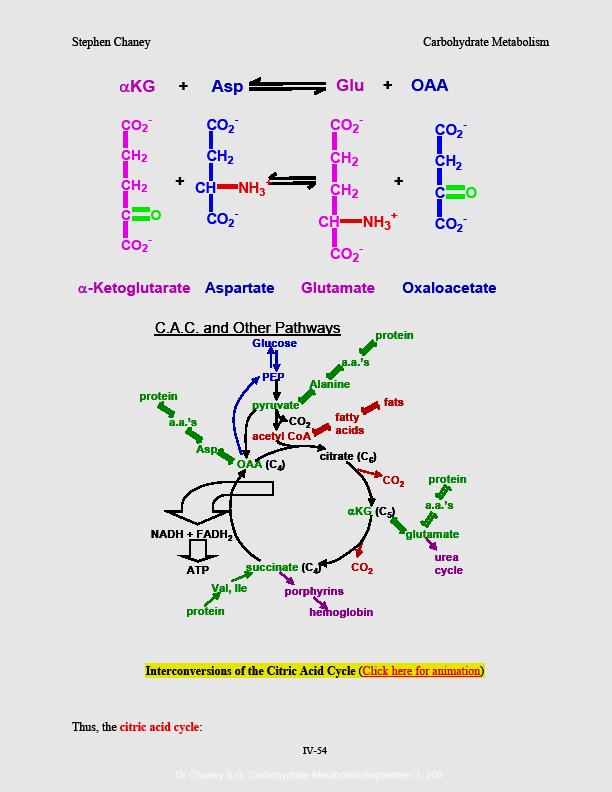
La glycolyse est suivie du cycle de Krebs qui conduit à davantage de production d'ATP (adénosine triphosphate). L'acide pyruvique, obtenu de la glycolyse, est oxydé en acétyl-CoA. C'est celui-ci qui va entrer dans le cycle de Krebs en se combinant à l'oxaloacétate pour former une molécule, le citrate, formée de trois fonctions carboxyles, d'où également la dénomination de cycle d'acide citrique ou de cycle tricarboxylique. L'ensemble nouvellement formé subit ensuite une série de déshydrogénations (équivalentes à des oxydations ) avec production de gaz carbonique (CO2), (d'ou l'appellation de cycle d'oxydation cellulaire ou de cycle de respiration cellulaire). Bien que ces réactions ne mettent pas directement en jeu la présence de l'oxygène moléculaire, elles sont néammnoins aérobies parce qu'elles ne peuvent se dérouler dans un environnement privé d'oxygène et parce qu'elles sont indirectement entraînées par la chaîne des transporteurs des électrons qui met en jeu l'oxygène moléculaire. Les molécules engendrées dans le cycle de Krebs vont subir la phosphorylation oxydative ou l'oxydation phosphorylante et engendrer de l'ATP;
Pour nous résumer, au cours de la glycolyse et du cycle de Krebs, des complexes générateurs d�ATP sont formés. Ces complexes - NADH et FADH - vont engendrer leurs ATPs lorsqu'ils seront capturés par la chaîne des transporteurs des électrons. Le cycle de Krebs situé au niveau de la matrice mitochondriale est donc intimement intégré dans la chaîne des processus qui menent à la phosphorylation oxydative, au niveau des crêtes mitochondriales.
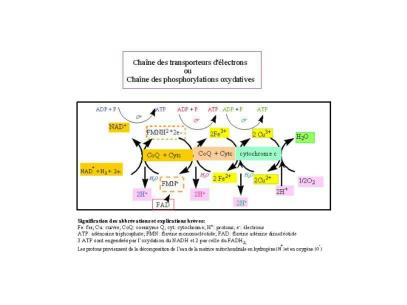 LA CHAÎNE DE TRANSPORT DES ÉLECTRONS ou chaîne des phosphorylations oxydatives. (cliquez sur mon schéma pour en voir les détails.)
(Cliquez ici pour être transporté à la page de M. Therry de l'Université de Connecticut qui en fournit une animation explicative.) (Ce troisième lien est réservé à l'auteur.)
LA CHAÎNE DE TRANSPORT DES ÉLECTRONS ou chaîne des phosphorylations oxydatives. (cliquez sur mon schéma pour en voir les détails.)
(Cliquez ici pour être transporté à la page de M. Therry de l'Université de Connecticut qui en fournit une animation explicative.) (Ce troisième lien est réservé à l'auteur.)
On peut voir comment chaque NADH engendre 3 ATP et comment la FAD n'engendre que 2; le processus est conduit par le gradient de protons (H+) le long de la membrane mitochondriale interne. comme nous l'avons signalé en voyant le cycle de Krebs. En tout 38 ATP peuvent être engendrées lors du catabolisme d'une molécule de glucose, 30 le sont par l'oxydation du NADH, 2 par celle du FADH, 2 le sont par la transformation de 2 molécules de GTP et 2 le sont directement au cours de la phase anaérobie de la glycolyse. En réalité, ceci ne se produit pas et seulement 30 ATP sont généralement engendrées au maximum lors du catabolisme d'une molécule de glucose, puisque la NAD, lui même, a été réutilisée (2 fois) et la FAD (1 fois) dans le cycle de Krebs et la réaction de la GTP couplée à celle de la formation d'ATP [C10H8N4O2NH2(OH)2(PO3H)3H]
est également réversible.
Les complexes enzymatiques faits de cytochrome et/ou deCoQ servent à véhiculer, sans danger, les électrons, à l'intérieur de l'organisme, comme nous le montre le schéma ci-dessus. L'oxygène engendre par la décomposition de l'eau sert à la chaîne de phosphorylations oxydatives (addition de radicaux phosphates) qui débute avec l'oxydation de la NADH et avec celle de la FADH (FMNH), pour se terminer avec la formation de l'eau et d'énergie sous la forme d'ATP, d'ou le terme de chaîne de respiration mitochondriale. Le gradient de protons conduit ou accélère la chaîne, en amenant l'oxydation de valences minérales.
Notes
1. Il est préférable de coupler la formation d'ATP au gradient de protons plutot que de le porter à la fin, (i.e., de se remédier au résultat final) afin de mieux comprendre ce qui se passe et de pouvoir équilibrer les équations des réactions.
2. Le complexe formé du CoQ (coenzyme Q ou ubiquinone) et de la cytochrome c constitue la cytochhrome réductase alors que la cytochrome c seule forme l'oxydase de la cytochrome.
3. En fait 1 molecule de NADH2 a ete reutilisee dans le cycle de Krebs, laissant (5-1=) 4NADH2) pour 2 molecules de pyruvate; 1 molecule de FADH2 et 1 molecule de GTP; ceci amene le total a: (2x4x3=24)+(2x(1-1/2)x2=2)+(2x(1-1/2)=1)+2 amenmant le total approximativement a 29 a 30 ATP
Résumé détaillé et cohérent des processus de la chaîne de phoshorylations oxydatives:
La chaîne débute avec l'oxydation de la NADH et de la FADH, parce que ces molécules organiques sont plus faciles à oxyder ou à céder leurs électrons (e-) que les atomes minéraux de la chaîne. Une fois débutée, la chaîne des processus d'oxydo-réduction continue d'elle-même, étant entrainée par le gradient des protons (H+) ainsi engendrés et par la décomposition subséquente de l'eau ambiante qui s'ionise (H+ et OH-). Tout au cours de la chaîne, des enzymes ou des complexes enzymatiques facilitent le transport des électrons et l'oxydo-réduction subséquente des atomes minéraux - cuivre et fer - de la chaîne. Les atomes d'oxygène, présents dans les radicaux OH-, servent à la formation ou à l'addition de radicaux phosphates (PO4--) à la molécule d'ADP pour la formation d'ATP. La formation d'eau, qui est le processus inverse de la décomposition, arrête le gradient de pression et termine la chaîne. Le bilan de la respiration cellulaire est, sans le cycle de Krebs, la formation de 3 molécules d'ATP par molécule de NADH et de 2 molécules d'ATP par molécule de FADH; 1 a 2 molécules d'eau ont été également utilisées. En fin de compte, les calories des aliments que nous ingérons ont été transférées et emmagasinées dans notre organisme sous la forme d'ATP. |
Conclusion sur le Catabolisme des Sucres
Ces réactions cataboliques ne concernent pas seulement les sucres, car de l'aétyl-CoA est également formé au cours du catabolisme des graisses et des protéines. Cet acétylCoA pénètre, ensuite, dans le cycle de Krebs. Celui-ci a un double rôle. En effet, il est amphibole parce qu'il fonctionne dans les deux sens - celui de l'anabolisme quand l'organisme est replet d'ATP et celui du catabolisme quand celui-ci est déplet d'ATP. En effet, le cycle de Krebs pourvoit l'oxaloacétate et l'α-cétoglutarate qui servent à la formation d'acides aminés ou aminoacidogenèse qui conduit à la protéinogenèse. D'autre part, l'oxaloacétate est transformé en phosphoénol pyruvate qui sert à la formation des sucres ou gluconéogenèse, et l'acétyl-CoA sert à la formation des graisses ou lipogenèse.
Un point important à souligner est que la gluconéogenèse n'est pas exactement le
revers de la glycloyse puisque trois étapes de celle-ci (1), (3) et (9) catalysées respectivement
par les enzymes hexokinase ou glucokinase, phosphofructokinase et pyruvate kinase sont irréversibles. Ces trois étapes correspondent à des réactions différentes dans la gluconéogenèse.
Enfin, des vitamines, comme celles du groupe B, sont importantes au catabolisme des sucres, en servant à la formation de coenzymes:
- la vitamine B1thiamine pyrophosphate ou TPP, qui entre dans la composition de deux enzymes déshydrogénases:
- la pyruvate déshydrogénase, un complexe enzymatique incluant les 3 autres coenzymes,qui catalyse la conversion du pyruvate en acétyl CoA
- et l'α-cétoglutarate déshydrogénase qui intervient dans le cycle de Krebs
.
- la vitamine B3 ou niacine que nous trouvons dans la nicotineamide adénine dinucléotide (NAD)
- la riboflavine ou vitamine B2, sous la forme de FADH
- et l'acide panthoténique que nous trouvons sous la forme de coenzyme A
Notes:
Les vitamines doivent être présentes dans l'alimentation, car elles ne sont pas fabriquées par notre organisme. La carence en vitamine B1, par exemple, cause une maladie sévère, connue sous le nom de Béribéri.
Celui-ci est particulièrement fréquent en extrême-orient où le riz représente le principal aliment et apparaît, surtout, chez ceux qui se nourrisent de riz décortiqué (riz blanchi).
AUTRES CYCLES NATURELS
Deux autres cycles cellulaires importants sont
- le cycle de Calvin Benson qui sert à la photosynthèse (� De la Fougère à l'Arbre) chez les végétaux
- et le cycle de Cori (cliquez dessus pour voir les détails) qui est particulièrement important chez les animaux musclés chez lesquels le malate, produit de la glycolyse musculaire, engendré dans le cycle de Krebs, sert à la gluconéogenèse hépatique.
Le cycle de Calvin Benson a été vu en classe de sixième et dans le cours de Physiologie
Végétale; une représentation graphique en est donnée ici.
Ont été également vus:
- les cycles des molécules organiques de la planète - cycle de l'azote, cycle du carbone (Calvin Benson) et cycle de formation et de décomposition de la matière organique (Calvin Benson et azote intégrés) (fig.) - avec les classes de Sixième et de Cinquième (� De la Fougère à l'Arbre, tome II)
- le cycle de la cellule vivante (fig ) (incluant la mitose des cellules somatiques et la méïose des cellules sexuelles) avec la classe de Rhéto (� L'Homme, ce Primate Supérieur)
- les cycles de vie de quelques organismes vivants (fig.) avec les classes de Sixième et de Cinquième (� De la Fougère à l'Arbre, tome I; De l'Ascaris au Colimaçon, tome II; De l'Éponge au Ver Plat)
- des cycles géologiques - cycle de l'eau; cycle d'énergie de la planète (fig. ) avec les classes de Cinquième et de Quatrième (� De la Fougère à l'Arbre; Géologie et Cosmologie, tome II)
- les réseaux et les chaînes alimentaires des communautés de populations (fig. ) avec la classe de
Cinquième (� De l'Ascaris au Colimaçon, tome II)
- et, dans le cadre du cours des Sciences
Naturelles des cycles cosmiques (Géologie & Cosmologie, tome III), avec les classe de Quatrième et de Troisième
Revoyez les.
Une caractéristique majeure de l'évolution de la vie sur terre est l'interaction constante entre les populations des êtres vivants qui ont y pris naissance et l'environnement dans lequel ceux-ci évoluent. De ce jeu constant et, en grande partie, expliqué par la sélection naturelle, sont nés les phénomènes d'extinction et d'espéciation et sont définies les grandes périodes d'évolution de la biosphère. C'est ceci que nous trouverons résumé dans la tableau suivant:
| FORMES DE VIE | FORMATION DE MONTAGNES
ET AUTRES EVENNEMENTS
MAJEURS |
EON |
ERE | PERIODE | EPOQUE | DE CETTE EPOQUE
A AUJOURD'HUI
(millions d'annees) |
|---|
| Premiers humains | Grande periode glaciaire, recente |
P
H
A
N
E
R
O
Z
O
I
Q
U
E |
C
E
N
O
Z
O
I
Q
U
E
| Quaternaire | Holocene (recente)
Pleistocene | <1 |
| Premiers mammiferes | Sierre Nevada (Amerique du Nord)
Alpes (Europe) | Tertiaire | Pliocene
Miocene
Olgocene
Eocene
Paleocene | 13
25
36
58
63 |
Grands reptiles
Amphibiens | Andes (Amerique du Sud)
Montagnes Rocheuses (Amerique du Nord
Jura (Europe) |
M
E
Z
O
Z
O
I
Q
Q
U
E | Cretace
| Jurassique
| Triassique
|
| | | |
Plantes terrestres:
-->
Charbon de terre
Vie marine:
- 1ers eucaryotes
- 1ers invertebres
- 1ers vertebres
| Appalaches (Amerique du Nord)
Oural (Europe) |
P
A
L
E
O
Z
O
I
Q
U
E |
Permienne |
Carbonifere |
Devonienne |
Silurienne |
Ordovicienne |
Cambrienne |
| | |
| Vie marine:1ere forme de vie | |
P
R
E
C
A
M
B
R
I
E
N |
P
R
O
T
E
O
Z
O
I
Q
U
E |
| | |
4000 - 2700 |
A
R
C
H
E
O
Z
O
I
Q
U
E |
|
| Premiers continents |
|
|
| | |
4300 |
Pour comprendre l'importance des interactions entre les populations et la notion de communauté intégrée, envisageons les répercussions sur la communauté de changements survenus dans une ou plusieurs populations qui la composent. La déprédation spécifique se produit fréquemment et inévitablement dans une communauté à la suite de modifications dans l'utilisation des terres par les organisme vivants. La réduction globale dans la taille des populations, la fragmentation des populations et toute altération dans l'abondance relative d'une ou de plusieurs espèces sont des phénomènes fréquents dans un écosystème qui peuvent avoir des implications majeures pour le reste de la communauté. Plusieurs tentatives d'explication ont été fournies aux effets probables de l'addition et de la soustraction d'espèces dans un écosystème. Chaque espèce, par exemple, peut être considérée comme une pièce importante dans la communauté de telle sorte qu'à sa disparition de l'écosystème s'en suivront des conséquences visibles ou appréciables sur le reste de la communauté. D'un autre côté, du fait du chevauchement fonctionnel qui existe entre les niches contingentes, la disparition d'une espèce de la communauté peut être facilement compensée. Entre ces deux possibiltiés extrêmes, on peut envisager toute une série d'alternatives intermédiaires qui dépendront de la période, de la zone géographique, de la densité de la population et d'un grand nombre d'autres paramètres géologiques et biologiques.
Une espèce-clef est donc celle dont l'impact sur la communauté peut être, ou est en fait, souvent sous-estimée si l'on se réfère uniquement à sa population, i.e., à sa biomasse. Par exemple, la présence ou l'abscence dans notre écosphère d'un virus comme celui du S.I.D.A a beaucoup plus d'importance dans notre communauté qu'elle peut détruire que celle du rhinovirus qui nous fait simplement avoir le "rhume de cerveau", même lorsque la population de celui-ci aurait été plus grande que celle de celui-là. D'un autre côté, un groupe fonctionnel est un ensemble d'espèces ayant des effets similaires ou quasi similaires sur un écosytème.
L'existence d'espèces-clef et de groupes fonctionnels rendent difficilement prévisibles les effets de l'addition et de la soustraction d'espèces sur l'équilibre d'un écosystème et sur l'intégrité de la comunauté qui en fait partie, et cela d'autant plus qu'il peut exister plus d'une espèce-clef et que certaines espèces peuvent appartenir à plusieurs groupes fonctionnels dans la communauté de celui-ci. Par exemple, le Varech, une algue marine, géante est non seulement un producteur pour les consommateurs de son écosystème mais fournit également à ceux-ci l'habitat dont ils ont besoin pour vivre dans cet environnement. D'autres espèces, comme, par exemple, celles qui forment le phytoplancton, peuvent facilement compenser pour la production si ces algues arrivent à manquer mais ne pourront nullement remplacer l'habitat manquant. Le Varech et les espèces qui forment le phytoplancton appartiennent à un même groupe fonctionnel mais le Varech est d'une toute autre importance car il appartient à un autre groupe fonctionnel auquel n'appartient pas le phytoplancton.
Un autre concept important à introduire en parlant de l'intégration communautaire est celui du processus communautaire qui englobe les interactions qui se produisent entre les diverses espèces qui constituent la communauté. Celles-là peuvent être la compétition, la prédation, le commensalisme, la symbiose ou le parasitisme. Par exemple, la disparition d'un prédateur, d'un consommateur secondaire ou tertiaire, d'un écosystème peut conduire à une explosion de la population d'un consommateur primaire qui peut mettre en danger l'intégrité de la communauté de celui-là.
Deux autres concepts importants sont celui de la distribution des populations et celui de la distributivité de l'écosystème. La première est, à la fois spatiale, temporelle et proportionelle. La seconde concerne la répartition des ressources naturelles qui sont les unes des ressources biotiques (les populations) les autres des ressources abiotiques (énergie, eau, ensoleillement, climat etc.). Ces deux aspects peuvent être liés. Par exemple, actuellement, un champignon eurasien, Cronartium ribicola, infecte et tue une espèce de pins (whitebark pines) du Glacier National Park que partargent les U.S.A. et le Canada, et menace, à la fois, d'affamer les ours et les oiseaux qui en vivent et de favoriser à partir des troncs desséchés des incendies de forêt dans cette région (" Natural History Review; Trouble at Timberline; November 1998; vol. 107, No. 9"). Les proportions de ces deux populations en seront affectées (distribution de populations) de même que la répartition des ressources premières (distributivité de l'écosytème) du fait de la modification de l'habitat.
Le principe de conservation et d'équilibre de milieux est non seulement important pour un organisme isolé mais l'est aussi pour des populations entières. Pour un organisme la conservation et l'équilibre de ses milieux intérieurs sont l'homéostasie. Nous avons vu ce que signifie ce terme au début de ce chapitre, et nous en avons eu une notion beaucoup plus approfondie dans l'appendice de L'Homme , ce Primate Supérieur. Nous n'y reviendrons pas. Cependant, je vous invite à revoir ces quelques lignes et quelques pages. Nous serions alors plus libres d'aborder les problèmes épimeux que posent la conservation et l'équilibre de notre environnement extérieur dans notre monde moderne.
Dans l'écosphère actuelle, les problèmes que posent la conservation et l'équilibre de l'environnement demandent de la part du citoyen le passage à l'action et la définition des priorités, pour la mise sur pied des mesures de protection et de restoration des écosystèmes et la protection de la diversité des espèces, des communautés et des génomes. Cependant, avant ceci, voyons les divers mécanismes qui entrent en jeu pour établir et maintenir la diversité d'une communauté. Nous nous arrêterons plus particulièrement à deux modèles: le modèle du réseau écologique et un modèle cybernétique utilisant une matrice écologique (fig.).
A. Le modèle du réseau écologique.
Ce modèle est particulièrement convenable aux petits écosystèmes et devient trop complexe pour un grand écosystème. Les relations entre les divers facteurs et les mécanismes qui contribuent à l'équilibre du milieu sont précises. Par exemple, on peut voir clairement sur la figure correspondante que la prédation à une influence sur le degré de compétition entre les espèces comme l'ont également le nombre de producteurs et des caractéristiques de l'habitat, comme l'hétérogénéité spatiale et la stabilité climatique. Bien que la figure n'indique pas dans quel sens se font ces relations, comme le feraient par exemple une matrice communautaire, (matrice de Levins) il est facile de le deviner, et on peut déjà dire que plus l'environnement est hétérogène moins la compétition interspécifique sera élevée. On peut même quantifier ces divers facteurs qui entrent en jeu dans l'écosystème et leur attribuer des valeurs paramétriques comme cela est pour la niche d'Hutchinson. Enfin, le modèle du réseau écologique nous donne une vue globale de de l'évolution d'un écosystème et nous montre, de façon claire, les divers agents - hétérogénité de l'environnement, productivité du milieu, compétition interspécifique, prédation et même, à un certain point, l'histoire de cet écosystème - de la diversité biospécifique.
B. Le modèle cybernétique utilisant une matrice écologique.
Une matrice, définie mathématiquement est faite de colonnes et de lignes. À l'intersection d'une colonne et d'une ligne se trouve une cellule de la matrice qui représente également une entrée. La matrice qui est représentée ici comporte deux colonnes et un plus grand nombre de lignes. La colonne de gauche caractérise un écosystème jeune ayant une explosion de bioversité et la colonne de droite caractérise un écosystème vieux ou le processus de biodiversification est ralentie. La première ligne a trait à la stabilité de l'environnement et la dernière l'évolution d'un tel écosystème, en particulier, la stabilité communautaire. Entre ces deux lignes, des intermédiaires concernant la productivité de l'environnement, la taille des niches et la grandeur de l'habitat. Un (+) désigne une corrélation positive et un (-) une corrélation négative (fig. ).
En analysant ce modèle, on peut constater qu'avec la progression de l'environnemnt vers la stabilité, la biodiversification s'accroît dans l'écosystème représenté. D'autre part, il y existe une corrélation positive entre l'accroissement de la productivité et le processus de biodiversification et il s'y produit vraisemblablement un chevauchement des niches.
Ce modèle cybernétique montre également que l'instabilité communautaire, comme celle qui est créée par des émigrations, peut accroître la stabilité de l'environnement en ralentissant, par exemple, la déprédation des producteurs.
LA FIXATION DES PRIORITéS
La protection de l'environnement régional demande une stratégie à l'échelle nationale et, sur une plus faible échelle, des actions communautaires. Elle impose des choix pour l'allocation des ressources afin de maximiser les bénéfices à long terme que le pays peut tirer de la biodiversité. Décider, par exemple, quels seraient les habitats et les niches à qui on devrait accorder la priorité dans l'allocation des ressources financières, humaines et institutionelles est une tâche ardue et peut le rester même avec l'avènement du computer. En fait, il n'existe pas de schéma universel pour fixer de telles priorités; tout choix dans ce sens sera aussi un reflet des valeurs humaines et prendra en considération les profits économiques, technologiques, sociaux et culturels que la communauté locale peut tirer d'une politique de protection des ressources de l'environnement. D'autre part, il n'existe pas de système prioritaire qui puisse satisfaire à tous les objectifs d'un plan de conservation.
LES MESURES
L'exploitation des ressources naturelles par l'homme a conduit à l'instabilité de l'environnement et à l'extinction réelle ou probable d'un grand nombre d'espèces. Dès lors, les moyens qui ont été mis en oeuvre pour la protection de l'environnement ont souvent mis l'accent sur l'éloignement des centres d'activités humaines des écosystèmes à préserver. Ceci ne peut néammoins pas profiter à long terme et sur une grande échelle aux communautés humaines. En effet, les hommes occupent ou exploitent 70% environ des écosystèmes des régions tempérées et tropicales pour produire 98% de leurs biens économiques. Dès lors, la plupart des environnements à préserver et la plupart des espèces en danger d'extinction se trouvent dans des zones géographiques occupées par des populations humaines et dans des mers visitées et exploitées par les hommes.
C'est pourquoi toute politique de conservation de l'environnement devra avoir de préférence pour tâche de minimiser les impacts des diverses activités humaines sur les écosystèmes et de s'efforcer de maintenir la biodiversité dans les sphères géographiques des activités humaines que sont, par exemple, l'agriculture, la foresterie, la pêche, le développement urbain, le tourisme et le transport.
|
{kind=link}
{kind=link}