De l’emplacement de l’ancien trou noir céleste, les "ingrédients" d’un nouveau monde ont émergé; un ancien monde y a fini. Ses nuages, ou nébuleuses de gaz, provenant de la fin brutale de ses grandes étoiles ont parsemé l’univers. D’eux sont nés de nouveaux soleils ou de nouvelles étoiles qui, à leur tour, ont formé d’autres mondes. Des planètes s’y sont sont formées et des planètes y sont mortes. Leurs continents ont pu se déplacer et leurs climats ont pu créer la diversité. Leurs atomes ont pu former des molécules et celles-ci ont pu s’assembler pour donner des cellules vivantes. Ainsi a pris naissance l’organisme vivant qui, subissant la sélection naturelle, a évolué, s’est diversifié, a proliféré et a disparu.
Dans une antichambre de ce scénario gigantesque et, ici, représenté d’une façon poétique et simplifiée, la terre ainsi que toutes les planètes de notre système solaire sont ainsi apparues, il y a de celà, 4.55 billions d’années. Au début, la surface de la terre était un océan de laves qui entourait un centre plus dur. Plus tard, il y a de celà 4.50 billions d’années, la terre se refroidit et ses premières roches se formerent. Plus tard encore, il y a de celà 4.2 billions d’années, les premières molécules organiques s’y formèrent, mais les premières cellules vivantes n’y apparurent qu’il y a de celà 3.5 billions d’années; la vie est apparue sur terre.
LA CELLULE, BASE DE LA VIE
La cellule est l’unité fonctionelle et structurelle de tout organisme vivant. Comme les blocs qui constitent un édifice, l’ensemble des cellules forme l’organisme vivant. Les cellules sont de taille et d’apparence diverses. Cependant, c’est leur nombre et non leur volume individuel qui détermine la grandeur de la taille d’un organisme. Les cellules d’une fourmi, celles d’un homme et celles d’une baleine sont, toutes, à peu près du même volume. Néammoins, la baleine est la plus grande de ceux-ci, parce que ses gènes ordonnent qu’un plus grand nombre de cellules y soient formées que chez la fourmi ou que chez l’homme.
Une cellule mesure, en moyenne, dix microns (micromètres), i.e., dix millionièmes de mètres. Elle est donc si petite qu’elle ne peut être décelée à l’oeil nu, et, à l’exception des cellules les plus grandes, ni même à la loupe. La taille d’une cellule animale varie, en moyenne, de 10 à 20 microns, c. à d., elle est cinq fois plus faible que la taille du plus petit objet qui peut être vu à l’oeil nu. Celle d’une cellule végétale varie, en général, de 1O à 100 microns.
Pour étudier la cellule, un instrument de grossissement des images - le microscope - est utilisé. Le microscope optique permet de voir des détails jusqu’à la grosseur de 2/10 micromètres de diamètre, i.e.., d’environ de 10-6m. Le microscope électronique permet de voir des détails plus petits de la grosseur d’une molécule ou de quelques angstromes, i.e., d’environ 10-11m. Des méthodes de contraste et la coloration des spécimens permettent d’augmenter encore la résolution de l’image observée au microscope. Cependant, toute étude de cellules exige des tranches de section très fines de tissus, faites avec un microton, une machine possédant des lames très fines de razoir qui permettent de faire des tranches de section aussi fines que 10 à 1 micron de diamètre.
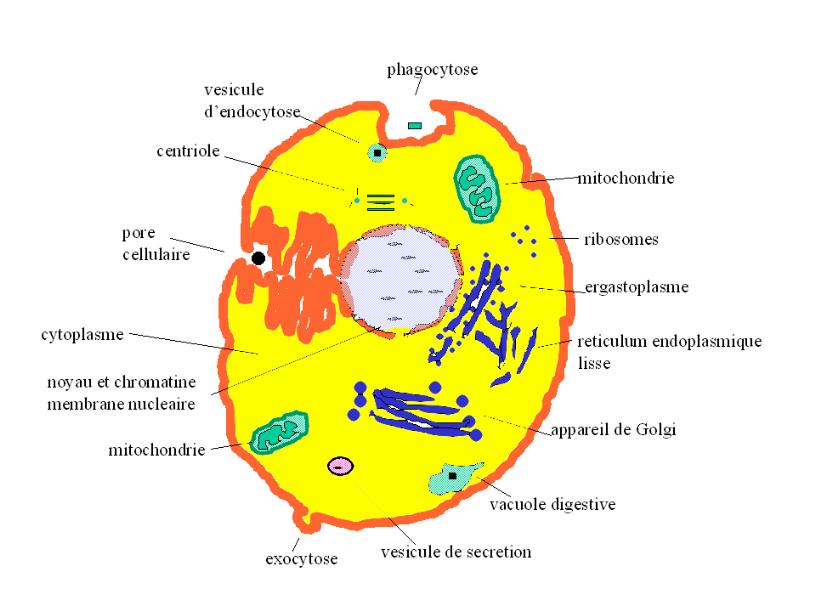
La cellule de la plante, tout comme celle de l’animal, est bordée d’une structure délicate-- la membrane cellulaire. Celle-ci a plusieurs rôles:
|
Les propriétés de la membrane cellulaire déterminent celles de la cellule:
- La cellule protège. La membrane cytoplasmìque de la cellule isole les organelles intérieures de l’environnement extérieur. Cette membrane permet la survie des structures délicates et internes de la cellule dans un environnement hostile et assure la stabilité du milieu intérieur de la cellule. Cette stabilité est ausi assurée par des mécanismes régulateurs qui se situent au niveau de la membrane cellulaire: pores, systèmes de transport actif et passif, gradients de pression et de concentration (ceux-ci sont souvent la conséquence de ces sytèmes de transport). Chez les eucaryotes, la membrane nucléaire isole et protège le matériel génétique de la cellule de la dégradation des enzymes cytoplasmiques.
- La cellule conserve. La membrane cytoplasmique empêche aux éléments intracellulaires, particulièrement les organelles qui représentent des structures fonctionelles intracellulaires de s’éparpiller à l’extérieur. La membrane nucléaire a un rôle similaire chez les eucaryotes pour protéger l’ADN nucléaire. Lorsque la cellule se divise, ces éléments intracytoplastniques et intranucléaires se divisent également, indépendamment ou non, de l’ADN nucléaire et se répartissent dans les nouvelles cellules sans être perdus à l’extérieur. La mitochondrie et le chloroplaste sont ces organelles qui se divisent indépendamment de l’ADN nucléaire et sont considérés par le biologiste Lynn Margulis comme des SYMBIONTS , i.e., des organismes vivant en symbiose avec le reste de la cellule eucaryote. Cependant, cette symbiose est définitive et héritable et ces organelles ne peuvent s’échapper de la cellule et y vivre en dehors.
Les éléments figurés du protoplasme sont, au niveau du noyau, les nucléoles, qui sont des amas de molécules d’acide nucléique et de protéine, et les chromosomes, qui apparaissent au cours de la mitose et qui sont formés de la condensation régionale de la molécule d’acide nucléique du nucléoplasme, additionnée de molécules de protéine provenant de la membrane nucléaire. Au niveau du cytoplasme, les éléments figurés du protoplasme sont les organelles: réticulums endoplasmiques lisse et granuleux, appareil de Golgi, mitochondries, plastes (entre autres les chloroplastes), lysosomes, centrioles. Les six premiers éléments figurés du protoplasme ont une fonction métabolique, le dernier a une fonction de reproduction (§ L’Homme, Ce Primate Supérieur, ch.I: La Cellule; Dr Roger Qualo; New York, 1998)( Physiologie Végétale; Reproduction Partielle, Critiques & CommentaIres du Cours des F.I.C.; Dr Roger Qualo; New York, 1999). [Revoyons les rapidement]:
Le réticulum endoplasmique est un système membranaire qui est impliqué dans la synthèse des molécules organiques, comme les protéines de l’organisme, celles-ci etant synthétisées au niveau des lysosomes de l’ergastoplasme (réticulum endoplasmique granuleux).
L’appareil de Golgi est un système membranaire qui est impliqué dans la synthèse des sucres complexes de l’organisme, telle la cellulose et dans l’excrétion de ces molécules complexes que sont les glycolipides et les glycoprotéines, formés respectivement de l’addition des sucres et de graisses ou de sucres et de protides.
La mitochondrie, ou chondriososme, est un organelle cellulaire au niveau duquel se fait le catabolisme (dégradation) des nutriments de la cellule avec production d’énergie sous la forme d’ATP. Les cycles d’oxydation mitochondriale sont le cycle de Krebs, ou cycle de l’acide citrique ou cycle tricarboxylique, et le cycle de phosphorylation oxydative cellulaire, celui-ci utilisant comme substrats les produits - NADH et FADH2 - de celui-là et brûlant, directement, l’oxygène moléculaire entraînant la production davantage d’ATP. Les substrats alimentaires sont transportés activement à travers la membrane mitochondriale, externe dans la mitochondrie (1). Là, ils sont attaqués par les systèmes enzymatiques de la matrice qui les transforment en un dérivé acétylé (2). Celui-ci sera attaqué par les enzymes qui se trouvent sur les crêtes mitochondriales (qui baignent dans la matrice) où il sera transformé en ces transporteurs d’électrons que sont le NADH et le FADH2 (3). Au niveau de la zone la plus périphérique de la membrane mitochondriale interne, ces transporteurs d’électrons se combinent directement à l’oxygène moléculaire, engendrant de l’énergie sous la forme d’ATP (4). Parce que ce processus implique, sous l’effet d’un gradient de pression, un transport osmotique de protons (H+) qui se combinent aux électrons et des processus chimiques, cataboliques, il a été dénommé "processus chimio-osmotique" ou "processus électro-chimioosmotique" (fig.1’-1)
Le plaste est un organelle qui est responsable de l’anabolisme cellulaire et au niveau duquel se fait la synthèse de novo des molécules organiques de la cellule végétale. Ce processus est, comme le processus catbolique qui se déroule au niveau de la mitochondrie, un mécanisme chimio-osmotique et met en jeu un gradient de pression et des transporteurs d’électrons. Ceux-ci sont localisés au niveau des membranes thylakoides qui sont des membranes empilées à l’intérieur du plastide.(fig. 1’-2)
L’ORGANISME DÉFINI, ICI, COMME UN ENSEMBLE DE CELLULES COORDONNEÉS
Quelle est l’importance historique des coacervats? Comment l’évolution s’est-elle faite de la cellule à l’organisme? S’est-elle faite par un assemblage de différentes cellules? Le premier organisme a-t-il été produit à partir d’une même lignée de cellules qui ne se sont pas séparées après leur mitose? Pour comprendre le fonctionnement d’un organisme, nous verrons donc, tout d’abord, ces trois thèmes - coacervat, colonie et reproduction - qui font la liaison entre les notions de molécule, de cellule et d’organisme.
Les Coacervats: La Vie Serait-elle Née d’un Assemblage de Molécules?
D’après les expériences de Sidney Fox et d’Oparin, des structures prébiotiques peuvent avoir fonctionné d’une manière similaire à celle des cellules primitives. Dans des conditions expérimentales qui rappellent l’atmosphère primitive, des molécules organiques, ordinairement des macromolécules, s’assemblent naturellement pour former des microsphères, encore appelées coacervats. Ces macromolécules sont des protéinoides (Fox) et, entre autres, des molécules d’ADN et d’ARN qui sont les molécules responsables de la repoduction cellulaire et de la synthèse des molécules organiques de la cellule.L’hypothèse que la vie serait née d’un assemblage disparate de molécules organiques a été fortifiée récemment avec la découverte de “mutations” spontanées dans la molécule d’ARN par épissage2 (“RNA’s splicing”) spontané et randomisé de cette molécule et par la découverte de la transcription à revers au cours de laquelle une molécule d’ADN peut être synthétisée à partir de la molécule d’ARN. D’autres données suggèrent que les cristaux d’argile peuvent avoir joué le rôle initial de moules et de facilitateurs des réactions métaboliques dans ces organismes prébiotiques. Si une cellule peut naître d’un assemblage de molécules, a fortiori, un organisme peut naître d’un assemblage de cellules de la même espèce.
Le Premier Organisme Multicellulaire Aurait-il Évolué d’une Colonie comme celle de Volvox?
Il est probable que les premiers organismes multicellulaire aient été formés d’un assemblage de cellules comme en témoignent la colonie de Volvox et, surtout, cet organisme si différent de nous qu’est l’Eponge (§ Dr. Roger Qualo; De l’Eponge au Ver Plat). Celle-ci est néammoins formée de plusieurs cellules disparates à la différence de la colonie de Volvox. Serait-ce le résultat d’un assemblage ou de la différenciation cellulaire à partir d’une même cellule produisant différentes lignées.La Reproduction Cellulaire Aurait-elle Été Responsable de la formation du Premier Organisme?
Il est aussi possible que la formation du premier organisme multicellulaire ait été dû à la division d’une cellule primordiale en plusieurs cellules qui sont restées accolées après la division cellulaire. Pour comprendre un tel processus, revoyons un peu le mécanisme de reproduction.Il existe deux modes majeurs de reproduction de la cellule: la reproduction asexuée et la reproduction sexuée. La reproduction asexuée est le clivage ou la division en deux nouvelles cellules de la cellule initiale; elle est précédée de la division du noyau de cette cellule. Celle-là, selon qu’elle s’accompagne ou non de la réduction du bagage initial de chromosomes, prend la dénomination de méiose ou de mitose. La méiose survient ordinairement au niveau des organes sexuels des organismes multicellulaires et s’accompagne de la formation de gamètes qui sont des cellules au nombre de chromosomes réduit de moitié par rapport aux autres cellules de l’organisme-mère. La mitose survient chez toutes les cellules qui se reproduisent.
Enfin, la reproduction sexuée s’accompagne de la fusion de deux cellules au bagage chromosomique réduit - les gamètes - pour former une cellule unique - l’oeuf - dont le bagage chromosomique normal de l’espèce est rétabli.
Qu’a t-il pu advenir ensuite? Chez les organismes multicellulaires, les cellules consécutives qui naissent de la division de l’oeuf (la cellule-mère) ont pu rester 1iées entre elles pour donner un nouvel organisme pluricellulaire. Il est cependant plus probable qu’au cours de l’évolution ces deux modes de production - par assemblage ou par absence de séparation des premières cellules - d’un organisme multicellulaire ont tous deux joué. En effet, l’analyse du caryotype de la mitochondrie et du chloroplaste, chez les cellules eucaryotes, fait penser que celles-ci seraient les produits d’une association symbiotique. La division de l’oeuf d’un eucaryote pluricellulaire fait toutefois penser au second mode.
Petit Rappel sur la Division Cellulaire
Au cours de la reproduction asexuée, la division du cytoplasme fait suite à celle du noyau et prend, suivant le cas, la dénomination de bourgeonnernent, de scissiparité (fission binaire) ou de cytodiérèse. Le bourgeonnement se fait par l’émission d’une projection ou d’un prolongement de la cellule et de son noyau qui se sépare ensuite du reste de la cellule, comme cela se voit chez le Saccharomyces pombe, la levure de la bière africaine. La scissiparité est la division de la cellule par scission, sans être précédée de phases distinctes de la division du noyau, comme cela se voit chez le Saccharomyces cerevissae, la levure utilisée dans nos boulangeries et dans nos brasseries. La cytodiérèse est la division normale du cytoplasme des cellules eucaryotes qui sont plus évoluées que les cellules procaryotes. Elle s’accompagne ou est précédée, chez les eucaryotes, de phases distinctes de la division nucléaire qui ont été partagées en prophase, métaphase, anaphase et télophase.
La méiose* (division réductionelle) et la mitose qui, chez les eucaryotes, sont les deux modes de la division du noyau connaissent, chacune, ces trois phases; la grande différence est que la mitose est précédée d'un dédoublement du matériel nucléaire, alors que la seconde mitose de la méïose** qui en connaît deux, n'est pas précédée de dédoublement du matériel nucléaire et conduit à la réduction (de moitié) du bagage nucléaire chez les cellules-filles, les gamètes. Chez le procaryote, la division nuclêaire procède différemment et est appelée, parfois, l’amitose.
(Voir aussi ces termes dans le glossaire pour éviter la confusion)
L’Organisme Pluricellulaire
Quelque temps après que les premières cellules qui fussent apparues sur terre eurent constitué des organismes pluri-cellulaires, la reproduction sexuée devînt le mode courant de reproduction. Encore appelée la fécondation, elle se produit par l’union de deux cellules sexuelles ordinairement différentes - un gamète male et un gamète femelle. Chez les organismes supérieurs, elle réclame l’appariement d’organes sexuels anatomiquement différents. Chez les animaux, en général, les cellules sexuelles sont le spermatozoide, chez le mâle, et l’ovule, chez la femelle. Chez les animaux supérieurs les organes qui hébergent les gamètes sont les gonades, qui sont, chez eux, les testicules chez le mâle et les ovaires chez la femelle.
Le processus d’union des cellules sexuelles, au cours de la reproduction sexuée, peut être externe (comme chez les poissons) ou interne. Dans la fécondation externe, le gamète mâle est déposé au voisinage du gamète femelle, et la pénétration de celui-ci par celui-là s’ensuit naturellement et est favorisée par l’environnement. (Dans un environnement aqueux, par exemple, les gamètes nagent.) Dans la fécondation interne, le rendement de l’accouplement sexuel est grand, car la fécondation est plus directe, contournant les caprices de l’environnement.
D’ autres modes de reproduction d’un organisme multicellulaire sont: la sporulation, la parthénogenèse et la régénération d’organes.
Dans la sporulation, les nouvelles cellules qui se forment sont recouvertes d’un revêtement épais qui les protège et les isole des conditions adverses de l’extérieur, permettant ainsi à la spore de memer une vie ralentie.
La parthénogenèse est la maturation d’un oeuf, ou ovule, pour donner un embryon, sans avoir éte fécondé au préalable; elle se rencontre chez quelques vertébrés, particulièrement chez les lézards et les crapauds (chez qui la parthénogenèse peut être induite expérimentalement en piquant, legèrement, l’oeuf avec une épingle). Elle se rencontre fréquemment chez un invertébré, l’oursin de mer.
Enfin, la régénération est un mécanisme commun de la réparation de quelques tissus chez les vertébrés et de tout l’organisme chez des invertébrés, comme l’hydre marine. Chez celle-ci, bien qu’il existe une coordination cellulaire qui fait fonctionner l’organisme comme un tout, chaque cellule est indépendante l’une de l’autre et l’animal peut se régénérer aisément.
Une autre preuve que l’organisme multicellulaire provient de l’évolution des premières cellules vivantes est donnée par l’embryologie. “L’embryogenèse récapitule la phylogenèse”.
Bien que cette phrase ne reflète pas, exactement, la réalité, un organisme multicellulaire a été défini comme un clone ordonné de cellules, toutes contenant le même génome mais n’ayant pas, toutes, la même fonction (Bruce Alberts et al.; Molecular Biology Of The Cell). Cependant, parce qu’un clone est défini comme un groupe de cellules génétiquement identiques, i.e., ayant le même génotype, et provenant d’une même cellule-mère par la division asexuée, cette définition pose quelques problèmes. En effet, chez l’organisme multi-cellulaire, le ZYGOTE qui donne naissance à celui-ci, bien qu’il le fasse par la division asexuée, est lui-même formé de l’union de deux cellules parentales, différentes - les gamètes - et non d’une seule, ce qui tendrait a faire écarter l’évolution comme cause de la biodiversité chez celui-ci. Néammoins, parce qu’on peut maintenant cloner expérimentalement des organismes multicellulaires, une telle distinction n’y tient plus.
L’argument con est qu’une différenciation cellulaire se produit inexorablement, chez la quasi-totalité des organismes multicellulaires, de la phase du zygote à la phase adulte, ce que l’on ne voit pas pour le clone de cellules. Les différentes phases de l’embryogenèse qui vont amener cette différenciation sont: la morulation, la blastulation, la gastrulation, la neurulation et l’organogenèse.
Enfin, l’argument pro est bien que ce soit le noyau qui détermine ultimement l’évolution de l’oeuf, cette différenciation est parfaitement compréhensible dans la thèse de l’évolution linéaire ou clonale parce qu’elle est promue par le cytoplasme dont le clivage créerait des particularités et, d’une manière générale, par l’environnement de l’oeuf, son(ses) micro-environnement(s), tout comme au cours de la phylogenèse, la diversité est promue par les macro-environnements de la planète.
Rappel Bref des Différents Stades de l’Embryogenèse
L'embryogenèse débute rapidement, seulement 30 heures après la fécondation. Au cours de son premier stade, la morulation, l’oeuf se divise rapidement, mais un de ses pôles se divise plus rapidement que l’autre et aura un sort différent. Ces deux pôles sont le pôle animal et le pôle végétal. La division de l’oeuf ne conduit pas non plus à des cellules de taille égale à celle de l’oeuf, mais à des cellules plus petites, d’où le nom de clivage, donné à ce processus et les cellules ainsi formées sont les blastomères.
A la morulation fait suite la blastulation au cours de laquelle l’embryon se creuse d’une cavité - le blastocèle - entourée d’une multitude de cellules. Il est maintenant une blastula.
A la blastulation fait suite la gastrulation. Au cours de celle-ci, se creuse, au niveau du pole végétal, une pochette - le bastopore - qui s’invagine et s’incurve ensuite pour encercler ce pôle. Le résultat ultime est un embryon à trois feuillets tissulaires qui sont de 1’intérieur vers l’extérieur: l’endoderme, le mésoderme et l’ectoderme.
A la gastrulation fait suite l'organogenèse qui débute 3 semaines après la fécondation, par la neurulation. Les différents feuillets formés au cours de la gastrulation auront, chacun, un destin différent. L’endoderme se creusera en un tube qui donnera le sytème digestif comprenant: le tube digestif, formé du pharynx, de l’oesophage, de l’estomac et des intestins, et les glandes qui luí sont associées- glandes salivaires, foie et pancréas. Cependant, alors que l’endoderme formera l’épithélium de ces structures, le tissu fibreux et musculaire de support viendra du mésoderme. Celui-ci donnera, en général, la colonne vertébrale et les os, en partie, le système génito-urinaire formé des reins et des organes de reproduction, enfin,le système cardio-vasculaire et le tissu sanguin. L’ectoderme formera la peau et le système nerveux au cours de la neurulation.
Cas Particuliers de 1’Embryogenèse
GROSSESSE MULTIPLE OU JUMEAUX
Comment peut- on expliquer la naissance de jumeaux?
La multiparité (les naissances multiples) quoiqu’elle ne soit pas rare chez les animaux est plutôt une exception dans l’espèce humaine. Chez les hommes, ordinairement , une grossesse gémellaire a deux enfants survient pour 86 grossesses uniques, une grossesse aux triplets pour 7.400 grossesses uniques, une aux quadruplets pour 635.000 et une aux quintuplets pour 55 millions. L’incidence de grossesses multiples est, néarnmoins, plus élevée chez les femmes plus âgées que 35 ans et chez celles qui prennent les médicaments pour combattre l’infertilité que dans la population des femmes, en général. Dans la plupart des cas, un ou plusieurs des enfants, nés au cours d’une grosesse multiple, meurent après l’accouchement.
Les jumeaux peuvent etre non identiques ou fraternels, encore appelés jumeaux dizygotiques. Ceux-ci sont formés quand l’ovulation simultanée de plusieurs oeufs survient et est suivie de leur fécondation simultanée. Chaque jumeau fraternel a, dans cette grossesse, son propre placenta, son propre cordon ombilical, son propre chorion et son propre amnion.
Une autre catégorie sont les jumeaux identiques ou monozygotiqueB. Ceux-ci proviennent de la fécondation d’un oeuf unique qui se divise, ensuite, pour donner deux oeufs complètement indépendants mais très rapprochés. Dès lors, ces jumeaux sont du même sexe, sont génotypiquement pareils et ont un placenta et un chorion communs. Cependant, les cordons ombilicaux sont séparés et les amnions sont aussi individualisés, sauf dans de rares cas.
Les jumeaux conjoints, encore appelés jumeaux siamois, sont des jumeaux identiques dont les disques embryonnaires ne se sont pas complètement séparés. Parfois les jumeaux siamois ne partargent qu’un morceau de peau, ils peuvent dès lors être séparés aisément. Plus souvent, les frères siamiois partargent un ou plusieurs organes et ne sont pas viables à plus ou moins long terme.
HERMAPHRODISME ET CHANGEMENT DE SEXE
L’hermaphroditisme est un état où l’individu a un sexe ambigü. il peut être d’origine hormonale ou d’origine génétique.
Dans 1’hermaphrodisme hormonal, ou pseudo-hermaphrodisme, la structure des organes génitaux - testicules et ovaires- sont normaux, mais les appendices génitaux externes, qui sont également les organes de l’accouplement, sont anormaux; le clitoris, par exemple peut être hypertrophié chez la femme sous l’action d’une hypersécrétion des hormones androgènes. Quelques cas d’hermaphrodisme hormonal surviennent lorsque les parents prennent des stéroïdes pour des raisons théerapeutiques ou pour tricher dans le sport. Le caryotype, qui est la photographie des différents chromosomes de l’individu, classés ou groupés selon une nomenclature sur une feuille de paipier ou sur l’écran de votre computer, y est normal (fig. 1’-3), (Le caryotype humain normal est de 22 paires de chromosomes autosomes, représentés et rangés sur une feuille de papier du chromosome 1 au chromosome 22, ce qui donne 44 chromosomes autosomes) et d’une pair de chromosomes sexuels, le tout totalisant 46 chromosomes. Pour l’homme, l’écriture du caryotype normal est 46,XY et pour la femme 46,XX).
Dans l’hermaphrodisme de nature génétique, ou hermaphrodisme vrai, l’individu a, à la fois, des ovaires et des testicules; le caryotype est souvent 46,XX, mais cecí n’est pas constant, et quelques individus ont un caryotype clairement anormal. Le généticien Polani présenta, en 1970, 108 cas d’hermaphrodisme vrai parmi lesquels 59 avaient le caryotype 46,XX, 21 avaient 46,XY et 28 avaient un mosaicisme, dont un certain nombre étaient 46,XX/XY.
Pour chacune de ces catégories, des procédés de correction chirurgicale sont possibles et amènent le changement désirable de sexe. Le procédé chirurgical est toujours individualisé et adapté au cas.
DÉTERMINISME SEXUEL
Chez les eucaryotes supérieurs, il existe deux types de progéniteurs - les monoécieux dont l’individu produit, à la fois, des gamètes mâles, et des gamètes femelles, i.e., dont l’espèce présente un monomorphisme sexuel, et les dioécieux chez lesquels il existe deux catégories d’individus, les uns produisant des gamètes mâles, les autres des gamètes femelles, i.e. où il existe deux sexes séparés et distincts et où il se présente un dimorphisme sexuel.
Chez plusieurs catégories d’insectes qui sont des dioécoeux, en particulier chez la sauterelle - un organisme diploïde - il a été trouvé que les cellules somatiques des femelles contenaient un extra-chromosome qui fut appelé le chromosome X. Plus tard, avec la mouche drosophile - également un organisme diploïde (ayant des pairs de chromosomes identiques) - il a été déterminé que les sécrétions gonadiques des femelles ne contenaient que des chromosomes X, tandis que celles des mâles contenaient, à la fois, des X et un autre chromosome en proportions égales, avec le même nombre de chromosomes pour chaque sexe. La conclusion a été que les femelles avaient deux chromosomes sexuels identiques - XX- alors que les mâles avaient deux chromosomes sexuels assimilaires - un X et un autre qui fut dénommé Y. Cette distribution se rencontre également chez les mammifères et chez certaines catégories de plantes, telles des angiospermes.
Cependant, le sexe n’est pas déterminé par des chromosomes (au niveau chromosomique) - théorie syngamique - chez tous les organismes qui présentent un dimorphisme sexuel. Chez l’asperge, par exemple, (une plante) il est clair que le sexe est déterminé (au niveau allélique) par un allèle (un gène d’une pair de gènes qui détermine le phénotype) dominant, le sexe mâle étant le caractère dominant, le sexe femelle le caractère recessif. En effet, chez l’asperge, qui présente un dimorphisme sexuel, quelques plantes ne portent que des étamines (des fleurs mâles) et sont donc des plantes mâles, d’autres que des pistils (des fleurs femelles) et sont donc des plantes femelles. D’autre part, des pistils rudimentaires se rencontrent sur quelques plantes mâles, et des étamines rudimentaires sur des plantes femelles. Ces organes sexuels rudimentaires sont ordinairement afonctionnels et abortifs. Il arrive que parfois quelques pistils se trouvant sur des plantes mâles sont fécondés par le pollen des étamines de celles-ci et produisent des fruits, mais l’inverse ne se produit pas. Ce "comportement est caractéristique de la dominance partielle, ou dominance incomplète (§ tableau de croisement des caractères):
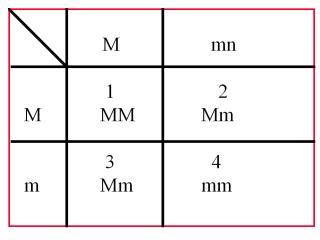
Fig.1: Tableau de croisement des caractères
Sur ce tableau M est le caractère dominant et m le caractre récessif; ensemble ils représentent le génotype (pour le trait sexuel), i.e., l’ensemble des gènes qui fixent le caractère sexuel. Dans la première case, la progéniture (F) est male, du fait de la présence des deux allèles dominants (individu homozygote) qui déterminent le sexe mâle. Dans le deuxième case, la fleur possède un allèle dominant et un allèle récessif et est donc hétérozygote, mais le phénotype (représenté par les traits extérieurs) est male, du fait de la dominance de l’allèle qui détermine le sexe mâle. Dans la troisième case, on peut dire que la plante est encore mâle en majorité, mais que, du fait de la dominance incomplète, l’allèle récessif se manifeste timidement et donne quelques pistils (organes femelles) fonctionnels. Dans la quatrième case, la plante est femelle et homozygote, du fait de la présence des deux allèles récessifs qui déterminent le sexe femelle. (Ainsi a été prouvé que le sexe est aussi déterminé génétiquement.)
LA POPULATION: UN ASSEMBLAGE D’INDIVIDUS (OU D’ORGANISMES) DE LA MEME ESPÈCE
Notre étude des populations débute avec le mimétisme, car celui-ci a quelqu’importance dans l’étude des interactions entre les populations, en particulier en ce qui concerne la prédation et, dès lors, dans l’équilibre écologique.
Qu’est le Mimétisme?
Le mimétisme est la ressemblance temporaire adoptee d’un animal à un animal d’une espèce différente, à une plante ou à un objet inanimé de l’environnement dans le but de permettre à celui-là d’échapper à un prédateur. Le mimétisme est déterminé immédiatement par des sécrétions hormonales, mais toutes les espèces d’animaux n’exhibent pas de mimétisme et, dès lors, le mimétisme est déterminé ultimement, i.e., initialement, par des gènes.
Autres Interactions entre Poplulations
Deux ou plusieurs populations peuvent interagir ou peuvent ne pas interagir entre elles. Si elles entrent en interaction, l’influence de l’une sur l’autre peut être bénéfique ou adverse. Dans l’un ou l’autre cas, cette relation peut être mutuellement bénéfique, mutuellement maléfique ou totalement bénéfique d’un côté et totalement adverse de l’autre. Quelques exemples de ces interactions sont la compétition, le neutralisme, le mutualisme, la coopération, la prédation, le parasitisme, le commensalisme et l’amensalisme. L’importance de ces interactions peut être déterminée quantitativement et un chiffre peut lui être attribué afin de permettre 1’ interprétation mathématique.
La relation de compétition survient quand des populations d’un environnement donné se battent pour la même ressource qui est en quantité limitée dans cet environnement et qui, dès lors, engendre une interaction adverse entre celles-là; une telle relation est d’ordinaire symbolisée (-,-). La relation de neutralisme survient, quand il n’existe aucune interaction favorable ou défavorable entre ces populations; elle est symbolisée (0,0). La relation de mutualisme existe quand il existe une interaction favorable entre ces populations et que leur association est nécessaire à leur survie; elle est symbolisée (+,+). La relation de coopération (ou de protocopération) existe quand l’interaction favorable entre ces populations n’est nullement nécessaire à leur survie; elle peut être également symbolisée matriciellement par (+,+) [dans le cas de ces matrices simples] . La relation de prédation existe quand l’une (les unes) de ces populations affecte(nt) l’autre (les autres) de façon adverse, mais, en général, les deux en bénéficient; elle est symbolisée (+,-) . Le parasitisme est similaire à la prédation, avec cette exception que l’hôte ou la proie (un membre de la population qui est adversement affectée) n’est pas tué sur le champ, mais est exploité pendant quelque temps avant l’issue finale; cette relation est habituellement symbolisée (+, -). Dans le commensalisme, une des populations bénéficie de l’interaction, tandis que l’autre n’en est pas affectée ou en est peu affectée; cette relation est symbolisée (+,0). L’amensalisme est une interaction dans laquelle une des populations est adversement affectée par une autre qui n’en est pas du tout affectée; elle est symbolisée (-,0).
De toutes ces relations, les trois plus importantes à retenir en Écologie Évolutionaire sont la COMPÉTITION, la PRÉDATION et la SYMBIOSE. Quand la relation de prédation existe, le mimétisme peut protéger la proie contre le prédateur. La symbiose est toute relation intime qui existe entre deux individus sans que l’un ou l’autre en soit adversement. affecté. Dès lors, la symbiose inclut, à quelque degré, le neutralisme, la coopération, le mutualisme et le commensalisme et peut être représentée par la matrice suivante [(0,0) (+,+,a) (+,+, b) (+,0)] ou a et b représentent respectivement la relation obligatoire et la relation non obligatoire. On peut simplifier les calculs matriciels en considérant, pour chaque colonne de la matrice, seulement une espèce affectée au cours d’une interaction entre deux espèces. Un exemple d’une telle matrice communautaire est comme suit pour une communauté naturelle de cinq espèces;
espèce affectée: individus ou populations
|
A, B, C, D et E: populations non affectées
Fig.2: Matrice communautaire
Ainsi, par exemple, au cours de l’interaction entre la population A et la population B, la matrice nous indique que la population A est affectée positivement (1ère colonne) et la population B est affectée négativement (deuxième colonne)->(+,-). Au cours de l’interaction entre les populations A et D, la population A n’en est pas affectée (première colonne) et la population D n’en est pas affectée (0,0)
La Population Mendélienne
Le brassage des caractères génétiques intervient pour chaque génération d’organismes qui se reproduisent sexuellement. Cependant, les traits hérités à la génération suivante ne sont pas dûs à un malaxage, mais sont hérités comme des entités distinctes ou indépendantes qui se présentent en pairs; ces entités sont les gènes. Mendel qui a découvert un tel mode d’héritage des caractères parentaux est connu pour être le père de la génétique. Souvent d’ailleurs, ce n’est qu’un un seul caractère de la pair qui s’exprime; c’est le caractère dominant. Celui qui ne s’exprime pas est le caractère récessif. Par exemple, si, après le croisement de deux fleurs, l’une rose et l’autre blanche, il arrive que tous les individus de la première génération sont roses et 3/4 de ceux de la deuxième génération sont roses et 1/4 en sont blancs, on dira que le trait dominant est rose (R), et que le trait récessif est (r), chaque trait étant hérité indépendemment.
P ou génération parentale: RR x rr |
Pour chaque génération d’organismes qui se reproduisent sexuellement, le brassage des gènes parentaux survient donc pour produire une nouvelle distribution des génomes. Dans une population donnée, une telle distribution, constitue le puits de gènes, et tous les organismes qui y participent forment une population Mendélienne. Les individus d’une telle population se reproduisent davantage entre eux qu’avec les individus d’une autre population, de telle sorte que les populations mendéliennes sont des groupes où il existe un échange substantiel de gènes. De telles populations sont encore appelées dèmes et l’étude de leurs statistiques vitales, telles que le taux de natalité, le taux de mortalité, le taux de fécondité, etc., est la démographie. Un exemple d’étude démographique nous est donné par l’histogramme suivant de la distribution de fréquence des cas de mortalité à des âges différents dans une population donnée (l’intervalle d’âges étant de 10 ans) :
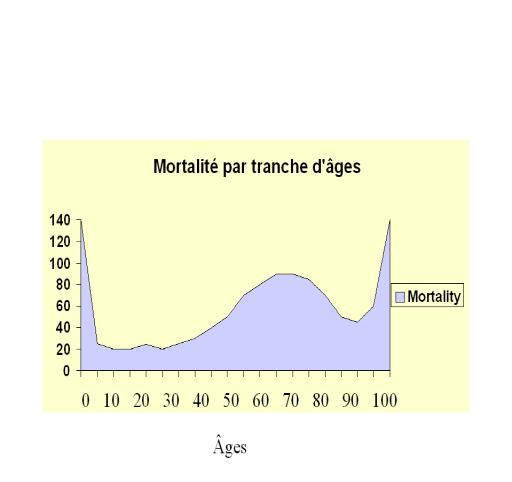
Fig.4: Taux de mortalité (nombre de décès par 1000 têtes de la population concernée) pour différentes tranches d’âges (intervalles de 10 ans)
Dynamique de Population (“Population Dynamics”)
La dynamique de population est ce chapitre de la démographie qui étudie la croissance et l’équilibre des populations, leurs cycles ou leurs oscillations, enfin leur densité. La croissance d’une population est, en définitive, appréciée par un paramètre: la moyenne ou le taux moyen. Cest ainsi qu’est défini un taux moyen de mortalité et un taux moyen de natalité, souvent calculés par tranches d’âges ou pour une cohorte d’individus, i.e., un ensemble d’individus du même âge et suivis depuis une même période, par exemple depuis leur naissance. Un autre paramètre qui permet d’apprécier les aptitudes d’une population est la variance qui permet d’apprécier la distribution de certains traits, par exemple.
Aucune population ne peut croître indéfiniment. Tôt ou tard, une population arrive à rencontrer des conditions difficiles d’existence dans son environnement - des conditions qu’on peut alors qualifier de Malthusiennes - et peut dès lors décroître jusqu’à son extinction. Des perturbations de l’environnement périodiques ou non, tels des incendies d’habitats naturels (ou de forêts), des inondations, des ouragans, des glaciers, des périodes de sécheresse, des éruptions volcaniques, des impacts d’astéroides ou de comètes, des secousses sismiques, enfin, tout cataclysme naturel peut drastiquement réduire la densité d’une population en deçà de sa capacité de croissance ou d’équilibre et dès lors conduire à son extinction. Plus fréquemment, la densité d’une population peut connaître des bas et des hauts et une évolution en dents de scie. La population des insectes et celle des petites plantes annuelles croissent rapidement, par exemple, durant l’été mais très peu durant l’hiver. De telles populations sont qualifiées d’opportunistes.
Il existe donc des cycles ou des oscillations de populations. Ceux-ci peuvent dépendre d’un certain nombre de facteurs, tels le taux de prédation, les ressources alimentaires, en bref, toute cause de sélection naturelle. Un type particulier de sélection qui opèie au niveau de l’individu semble déterminer la sélection naturelle des groupes. C’est celle qui détermine la lignée familiale (“kinship”). Pourquoi, en effet une abeille ouvrière ne se reproduit-elle pas, laissant ce souci à la reine de la ruche, et dès lors sacrifiant, ses propres gènes qui ne subiront pas la sélection naturelle? Cette ouvrière est aussi prête à sacrifier sa vie pour la défense de la ruche. Il est néammoins clair que le comportement altruiste ne procure aucun avantage à une population en fait d’évolution par la sélection naturelle. Pour expliquer cette apparente antithèse, Hamilton nous dit. que la sélection naturelle opère au niveau de la lignée. C’est parce que les individus qui vivent en groupes ont davantage de représentants de leurs gènes dans le puits de gènes de leur population, véhiculés par leur reine, et qu’ils sont, dès lors,plus apparentés que s’ils vivaient isolément, comme des hermites. En d’autres termes, la reine des abeilles qui est aussi la seule femelle à se reproduire dans la ruche porte les mêmes gènes que les différents individus de son peuple qui auront, dès lors, cet avantage énorme d’avoir plus de chance de voir sélectionner leurs propres gènes ou des gènes qui leur sont identiques et de se perpétuer à travers l’espèce.
---------------------------------------
1 L’épissage de l’ARN est une coupure dans la molécule d’ARN, au niveau des introns (parties non codantes pour une protéine), suivie du raccollement des extrons (parties codantes).
** Réservé à l’auteur