

|
The Ascent of Man (continued) With quadrupedalism and the slow rate of food acquisition, an equilibrium would have resulted balancing off the benefits of inter-group competition and intra-group scramble competition. In other words, the evolutionary trend would slide into a log jam where groups would fare better with more members, but they would not have ready access to the resources to support any additional members. 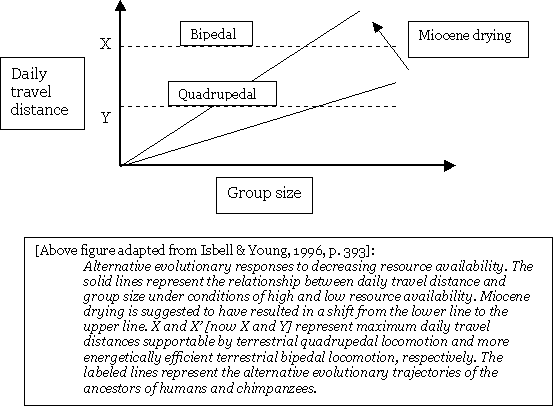 In order to blast their way out of this log jam, our hominoid ancestors resorted to two basic strategies. Firstly, they could maintain quadrupedalism and evolve a fission-fusion system, or alternatively, they could evolve bipedalism and also larger group size. The former route was taken by the ancestors of chimpanzees. The latter option was selected by our ancestors. The tendency to aggregate into larger groups under certain circumstances has been observed among other primates as well, albeit resulting from a trait other than bipedalism. For example, a population of vervet monkeys developed larger group sizes in response to a shift toward a drier habitat, and subsequently outcompeted their surrounding groups through inter-group scramble competition. (Cheney & Seyfarth, 1987; Isbell et al., 1990; cited in Isbell & Young, 1996). The Isbell and Young Hypothesis has been criticized on a number of grounds by such paleoanthropologists as Kevin Hunt, who developed the Postural Feeding Hypothesis.6 He argues that while a fully bipedal individual may gain a locomotory advantage over a quadruped, a facultative biped will not. He points out that the intermembral index of approximately one hundred percent for chimpanzees and proto-hominid ancestors would not have facilitated efficient bipedalism. However, according to an American Journal of Physical Anthropology paper, “These and other results from the recent literature suggest that increased lower limb length provided no selective advantage in locomotion and other explanations should be sought.” (Webb, 1996). Therefore, fore- to hindlimb proportions as a major factor in early hominid evolution should best not be accepted.7 Notes:
Copyright ©2001-2003, Allegra H., all rights reserved. Please contact me via e-mail if you wish to reproduce this material.
|













