|
| |||
The research presented here is being performed at the All-Russian Institute of Plant Protection, Microbiological Control Department, in the Microsporidia Research Group. Part of the work is done through cooperation with different research groups, as indicated below. INSECT IMMUNITYExamination of insect immunity helps to understand why insects are resistant or susceptible to a variety of harmful factors of environment. One of the most interesting point is understanding that
irritation of insect immune system by one kind of agents cause altered sensitivity to others.
This leads to explanation and maybe prediction of dynamics of development of mixed infections and
of insect pathogenesis under stress conditions.
Host immune responses, underlying parasite-host interactions of microsporidia and insects, are poorly studied.
In our lab, we study various aspects of host immune reactions
in three microsporidia-insect parasite-host systems: Gryllus bimaculatus - Paranosema (=Nosema) grylli; Locusta migratoria - P. locustae; and Galleria mellonella - Vairimorpha ephestiae.
Cellular ReactionsMajor cellular reactions of crickets and locusts to microsporidian infection are
phagocytosis of parasite spores, incapsulation and melanization of infected cells. During early
stage of disease, no signs of cellular responses are found in cricket tissues. At this period,
proliferate parasite stages reside inside the host cells, avoiding non-self recognition by host
haemolymph. Melanized nodules and incapsulated cells within infected fat bodies and lymph glands
of crickets are associated with mature Paranosema grylli spores. At the acute stage of
disease, when mass sporogony of microsporidia occurs, up to 90% crickets manifest melanization
of infected tissues. Incapsulation of infected cells is mediated by plasmatocytes and
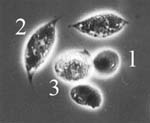
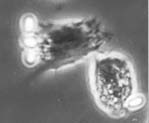
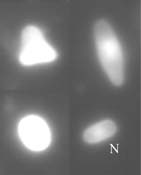
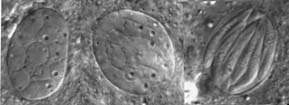
granulocytes, the major cell types of cricket haemocytes (Fig. 1) (Sokolova et al., 2000). Ruptured infected cells release large
amount of spores into the body cavity, were phagocytosis takes place (Fig. 2). Spores uptaken by
phagocytes both in vivo and in vitro undergo acidification and degradation within the phagosomes.
Some of the incapsulated and phagocytized spores are exfilamented, thus escaping elimination by
host immune system.
Humoral FactorsMany types of molecules in insect haemolymph plasma serve recognition of non-self,
opsonization of pathogens and further regulation of immune response in the insect organism.
Examination of agglutinins from the blood of cricket Gryllus bimaculatus is being performed with collaboration with
laboratory of insect pathology, headed by Dr. V.V. Glupov, at the Institute of Systematics
and Ecology of Animals, Novosibirsk, Russia. It was found found that spores of the specific cricket parasite,
microsporidian Paranosema grylli, are agglutinated by host haemolymph plasma, and that
accumulation of melanized cores containing microsporidian spore masses
cause removal of agglutinating activity from the plasma at the acute stage of disease.
The protein fraction with agglutinating activity has been partially purified and its
physico-chemical properties were examined. Our next goal is to study involvement of theseb
molecules in microsporidia recognition and triggering the defence reaction cascades under
conditions of infectious process.
Phenoloxidase cascade
Melanization, mediated by phenoloxidase (PO) system, is a principal defense reaction of insect haemolymph against invading pathogens, and many parasites exploit diverse strategies to suppress or avoid action of POs. Involvement of PO in interactions of microsporidia with insect immune system is poorly studied. We have demonstated previously, that microsporidia P. grylli suppress PO in haemocytes of G. bimaculatus at the acute stage of microsporidiosis (Sokolova et al., 1999, 2000). In the present study it was found that:
Haemocytes in vitro
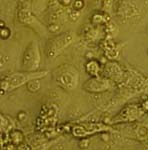
Cultivation of insect blood cells (haemocytes) is an important tool for examination of cellular immunity in insects. Being target cells for microsporidian infection, haemocytes could also serve as a model of host cells for cultivation of these obligate intracellular parasites. To our knowledge, no one have cultivated haemocytes from the cricket G. bimaculatus so far, an insect that is a suitable laboratory model for research in many fields of entomology, including insect pathology. The research was conducted in collaboration with laboratory of parasitology, headed by Prof. R. Entzeroth, Dresden University of Technology, Germany.
INSECT PATHOGENSInsect pathology is a fascinating world where interests of several disciplines meet: entomology, parasitology, microbiology, ecology etc., and applied sciences, such as biological control of pests and disease vectors, have a lot to do with it as well. Alongside with microsporidia research, which has the utmost priority in our scientific group, other pathogens of insects are also in the scope of our attention, both fundamental and applied.
Locust BiocontrolDevelopment of biological control measures of locusts and grasshoppers is one of most priority and fast developing directions in modern IPM programs. The most important from this point of view are entomopathogenic hyphomycetes (Deuteromycota, Hyphomycetes).
Due to a high level of reproduction of Calliptamus italicus and Locusta migratoria on the south of Russia, we studied activities of fungi against locusts under lab and field conditions, together with the Fungi research group (Dr. G.R. Lednev, Ph.D. student M.V. Levchenko) from our institute.
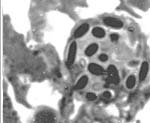
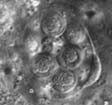
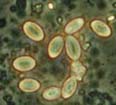
Mixed InfectionsInfections caused by more than one pathogen simultaneously are of particular interest, as in nature, where many types of organisms meet, probability of pathogenesis with mixed ethiology is quite high. It is also important to know how would reactions of an insect population to implication of chemical and biological control agents differ under influence of naturally occuring or introduced parasites. Besides microsporidia, in our lab cultures of orthopteran insects there's a set of other pathogens: coccidia (Fig. 9), gregarines and viruses in crickets; gregarines and amoebae (Fig. 10) in locusts. Observations of dynamics of development of these parasites are done during experimental microsporidiosis to elucidate role of microsporidia in mixed infections, as the latter suppress host immunity system. Moreover, in bioaasays with microsporidia and fungi (Beauveria bassiana and Metarhizium anisopliae), that have potential in locust biocontrol, we study alterations of insect sensitivity to application of fungi and their toxins at certain phases of microsporidiosis .
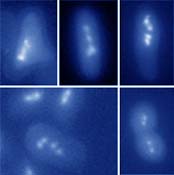
Diagnosis in lab and fieldIndentification of pathogens in field populations and lab cultures of insects is necessary for a wide spectre of reasons, including right interpretation of results of physiological studies, survey of prevalence and distribution of parasites in host populations, search for new species of protists and prediction of insect pest densities changes under pressure of pathogens. Main methods for diagnosis of pathogens in insects are routine light microscopical observation combined with histochemical staining. Staining with fluorescent probes for nuclei and chitin-like compounds (see, for example, Fig. 4) are employed for more precise recognition of parasitic stages and some characteristic features of their cells. Methods for rapid determination of viability of invasive stages of microsporidians and other pathogens are also being tested and compared in our lab.
|
||||||||||||||||||||||||||||||||||
| judas prokaryotes 2004 |