
La sistemática filogenética, ahora frecuentemente referida como "cladística", "cladismo", "Hennigismo" o "Método Hennigiano", fue introducida entre los biólogos por el entomólogo alemán Willi Hennig (1950), con un libro que es el pilar de la historia de la biología comparativa y la ciencia evolutiva. La meta de Hennig fue un método objetivo para la reconstrucción de filogenias, que son las genealogías de los organismos, por el establecimiento de clasificaciones que reflejen dichas filogenias. Su método fue una respuesta al hábito generalizado entre los biólogos y paleontólogos de clasificar los organismos sobre la base de criterios heterogéneos (morfológicos, etológicos, etc.) y de acuerdo a su totalidad de similitudes. Desde el siglo XVIII, muchas clasificaciones reflejaron mas la opinión personal de los "especialistas" que la historia real de los grupos, y dependieron principalmente de la importancia dada, a un carácter en particular, por el especialista en cuestión. De aquí los frecuentes y ridículos debates sobre las clasificaciones, que contribuyeron a dar la imagen popular a los "naturalistas" de biólogos anticuados. Para enlazar las clasificaciones a un evento único, no arbitrario, por ejemplo la historia de la vida misma, Hennig abatió el peso de las autoridades establecidas en este campo de las ciencias de la vida y también ejecutó las predicciones de Darwin de que las clasificaciones podían ser genealogías (Tassy, 1983).
Aunque el interés por las ideas de Hennig fue reconocido hasta los años 50's por Kiria Kolf (1955), Gunther (1956) y Brundin (1956), que discutieron y polemizaron sobre la sistemática filogenética, esta no se generalizó sino hasta dos décadas después, este dilatamiento se debió, probablemente, a barreras del lenguaje y fue vencido por la traducción al inglés de muchas versiones revisadas del libro de Hennig por Davies y Zangert (1966), así como la popularización de las ideas de Hennig por parte de Brundin (1966-1968). Estas publicaciones ocuparon un lugar principal dentro de los métodos revolucionarios en filogenia y sistemática.
Además de su MAGNUM OPUS, Hennig escribió un buen número de notas anteriormente (Hennig 1936, 1948, 1949) en que sus ideas se vislumbraban claramente, y varias notas posteriores en defensa de su método (Hennig 1953, 1974, 1975). Hennig murió en 1976, justo cuando se iniciaba un debate acerca de la cladística. Este debate pudo continuarse a lo largo de la última década en los números del Journal of Systematic Zoology, y de una manera más oscura en volúmenes recientes de Nature, sirviendo como un campo de batalla abierto; Systematic Zoology pudo mantenerse en la ciencia como uno de los mayores catalizadores en esta renovación de los conceptos biológicos y Yo aliento al estudiante interesado a leer las series de artículos en el Journal que hacen una crónica de la evolución cladística, desde la Sistemática Filogenética Hennigiana clásica hasta los llamados "Cladistas Transformados" (o "Cladistas Modernos").
De acuerdo con Brundin (1972), la contribución de Hennig puede ser recordada como "uno de los mayores avances metodológicos en la biología causal, después de Darwin". La historia de la sistemática filogenética puede ser también encontrada marcadamente sumarizada por Dupuis (1978). Los puntos de vista originales sobre el lugar de la Cladística en la historia de la biología son expresados por Nelson ( ) y Platnick (1981).
El propósito de este artículo es presentar los principios generales de la sistemática filogenética de Hennig, así como sus subsecuentes transformaciones por algunos teóricos de la cladística, posteriormente, también, se tratará de pronosticar el posible desarrollo futuro de la cladística, considerando los factores material y humano.
Los fundamentos de la cladística han sido explicados muchas veces, ya sea en forma semipopular (Bonde, 1975; Patterson, 1978; 81a, 82a; Janvier et al., 1980; Goujet, 1981) o en excelentes libros de texto (Cracraft, 1979; Eldredge and Cracraft, 1980; Nelson and Platnick, 1981; Wiley, 1981) y se pueden concentrar algunos aspectos controversiales que pueden ser eterna discusión en los 80's. Dichos aspectos son el uso de la parsimonia en la reconstrucción de filogenias y las preguntas sobre implicaciones evolutivas de la cladística. También se da una hipótesis sobre algunos escenarios posibles para el futuro de la cladística, considerando su surgimiento en varios campos como la Biogeografía, Geología, Fisiología o Bioquímica.
La sistemática filogenética o cladística pertenece a uno de los dos campos principales de la biología, denominada BIOLOGIA COMPARATIVA por Nelson (1970). La Biología Comparativa concierne a la diversidad de los organismos, la distribución de caracteres en ellos y el posible significado de este patrón (Wiley, 1981). En contraste la BIOLOGIA GENERAL, tiene que ver con los procesos que generan la diversidad, y uno de los mayores fracasos de la biología moderna es que frecuentemente se descuida el aspecto comparativo de los problemas. En otras palabras, es mucha la importancia que se le da a la pregunta "¿Por qué?" Y muy poca la que se le da a la pregunta "¿Qué?". El propósito de la Biología Comparativa es estudiar que son realmente las cosas, o como unas están relacionadas con otras, la especulación subsecuente de porque es esto pertenece a la Biología General. Nelson & Platnick (1981) y otros Cladistas, consideran que este énfasis generalizado fue heredado de Wallace (1855) y Darwin (1859) quienes, por medio de la proposición de un modelo del proceso evolutivo (Selección Natural) iniciaron la caída de las pautas de análisis.
El término filogenia fue recopilado por Haeckel (1865) y significa en su sentido original la historia del desarrollo paleontológico de una especie. Actualmente la filogenia expresa las interrelaciones de los miembros de un grupo, como resultado de la evolución debida a la descendencia y transformación de los carácteries hereditarios; la reconstrucción de la filogenia está basada en la búsqueda de caracteres compartidos por uno o más organismos. Ya que los diferentes caracteres han aparecido en diferentes épocas en la historia de un grupo de organismos, estos muestran una jerarquía en su distribución, o en una generalidad que está relacionada con su antigüedad.
La cronología de la ocurrencia de unos caracteres conduce a la reconstrucción de un ARBOL FILOGENETICO del grupo en que las especies ancestrales son inferidas como puntos de ramificación.
La SISTEMÁTICA es el estudio de la diversidad de organismos, e intenta relacionar esto en base a cualquier criterio de relacionamiento. Se han propuestos varios sistemas por parte de naturalistas desde la época de Aristóteles, y el sistema construido por Linneo (1758) es sin duda la base de la sistemática actual. Dentro de la sistemática no-hennigiana, se utilizan tres criterios mayores para agrupar organismos y categorizar dichos agrupamientos: La totalidad de similitudes, similitudes ecológicas y, en algunos casos, relaciones filogenéticas, cuando están expresadas en caracteres conspicuos (pelos en mamíferos, aves con plumas, etc.). Por esta razón los sistemáticos que usan todos estos criterios se llaman a sí mismos SISTEMATICOS ECLECTICOS (Simpson, 1975) (ECLECTISISMO: Método que consiste en reunir lo mejor de la doctrina de varios sistemas). Sin embargo muchos Sistemáticos Eclécticos como Simpson, Mayr y sus seguidores dan mucho peso a los procesos evolutivos y tratan de producir clasificaciones que expresen descendencia. De este modo, ellos denominan grupos ancestrales y grupos descendientes, y el rango de este último expresa frecuentemente el grado de divergencia, como es estimado por criterios que incluyen la opinión personal del sistemático o "ingenuidad" (Simpson, 1961). Ya que se supone que estas clasificaciones expresan la evolución de grupos, los Sistemáticos Eclécticos son llamados también SISTEMATICOS EVOLUTIVOS.
Los sistemáticos que solo usan el criterio de totalidad de similitudes, pudiendo usar cualquier método para manipular estos datos, son llamados SISTEMATICOS FENETICOS.
Finalmente, los sistemáticos que consideran que las clasificaciones reflejan solo relaciones filogenéticas y desconocen la divergencia morfológica, son denominados SISTEMATICOS FILOGENETICOS, ya que la mejor manera de expresar las relaciones filogenéticas son los diagramas ramificados denominados CLADOGRAMAS (Klados= rama, en griego) y debido a que los Sistemáticos Filogenéticos solo forman grupos naturales (=CLADOS), en consideración, Mayr (1969) concibió el término "CLADISTAS" para referirse a esos sistemáticos.
La TAXONOMIA es el aspecto técnico de la sistemática, Simpson (1944, 1961) la define como la base teórica de los principios, reglas y términos de la clasificación. El término taxonomía, concebido por Candolle (1813), designa todas las actividades concernientes al uso de un nombre unido a un grupo de organismos, o TAXON (plural TAXA ó TAXONES), así como la descripción de esos grupos y la historia de su descubrimiento. La especie es considerada generalmente como la única categoría no arbitraria, habiendo tenido, no obstante, muchas diferentes definiciones que reflejan la controversia sobre sus limites. Las especies están relativamente bien limitadas en la naturaleza, por ejemplo, debido a su aislamiento reproductivo, pero los criterios definitorios de especie, han sido, sin embargo, fuertemente discutidos por los biólogos. Las categorías supraespecíficas no poseen la misma realidad que las especies; estas son agrupamientos del hombre basados en criterios escogidos por los sistemáticos, los intentos de encontrar criterios que definan categorías supraespecíficas han sido un fracaso. Dubois (1982) por ejemplo, propuso recientemente una definición de género sobre la base de la habilidad de las especies congenéricas de producir híbridos viables, pero Yo dudo que esta definición pueda ser aceptada por todos los biólogos.
En su sentido original, como Hennig lo definió, la Sistemática Filogenética descansa en el postulado de una descendencia como modificación o evolución en el sentido moderno, ha tenido lugar. Por lo tanto, los caracteres observados en cualquier organismo pueden mostrar un estado primitivo o uno avanzado (derivado). Como podemos ver, muchos cladistas rechazan este postulado evolutivo, considerándolo innecesario. Esta nueva forma de la Cladística, frecuentemente denominada como Cladística Transformada, es difícil de entender por los biólogos entregados a la tradición evolutiva, y es algunas veces erróneamente considerada como un creacionismo científico. El fundamento de la Sistemática Filogenética de Hennig es que las similitudes de los organismos no siempre tienen el mismo significado o apariencia en las relaciones entre estos. Los organismos o grupos de organismos se asemejan unos a otros porque comparten caracteres avanzados o primitivos. Hennig llamo a los avanzados (o derivados) caracteres APOMORFICOS y a los primitivos, caracteres PLESIOMORFICOS.
Cuando dichos caracteres están compartidos por varios individuos entonces son caracteres SINAPOMORFICOS O SIMPLESIOMORFICOS. Hennig considera que las sinapomorfías sola indican relaciones entre organismos, y que todos los organismos que comparten una sinapomorfía forman un grupo monofilético, que es una historia aparte. La fúrcula y la presencia de plumas son sinapomorfías de los pájaros que forman el grupo de las aves. Una sinapomorfía puede desaparecer secundariamente o empezar a modificarse dentro de un gran grupo monofilético que la define; la ontogenia o convergencias con otras sinapomorfías revelan, frecuentemente, aquellos casos de REVERSION (Ejemplo, la pérdida de pelo en las ballenas). Un taxón o grupo monofilético incluye una especie ancestral común y todas las especies que descienden de ella y es reconocido por al menos una apomorfía que aparece en el ancestro común. Una sinapomorfía puede ser un carácter HOMOLOGO, que es un carácter heredado de un ancestro común. Nosotros podemos ver posteriormente que los criterios para determinar una homología son muy discutidos y que los "Cladistas Transformados" proponen ahora, con buenas razones, sinonimizar sinapomorfía y homología.
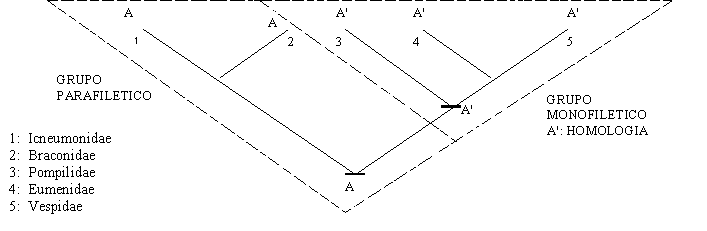
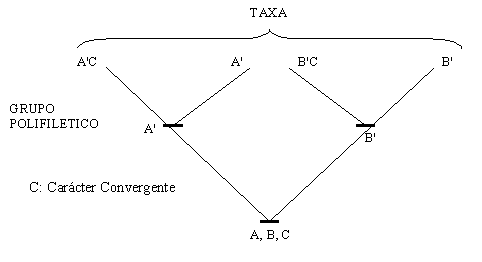
Las simplesiomorfías, o caracteres primitivos, compartidas indican un ancestro común, pero no inmediato y exclusivo. Los grupos de taxa definidos por simplesiomorfías fueron llamados PARAFILETICOS por Hennig; por ejemplo, los peces, como grupo definido por la presencia de aletas pareadas, son un grupo parafilético debido a que los miembros pares de los tetrápodos son aletas modificadas y porque algunos de los peces comparten sinapomorfías con los tetrápodos, que no están presentes en los otros peces. Por lo tanto, las aletas pares de los peces son una simplesiomorfía para ellos, pero una sinapomorfía para los gnatostomados (Peces más tetrápodos). En contraste, un grupo que incluya a los tetrápodos y a aquellos peces que comparten ciertas sinapomorfías con ellos, como los huesos mandibulares, puede ser monofilético (Como los Osteichthyes en el sentido cladista, que incluye a los tetrápodos). Aunque estos ejemplos pueden parecer triviales, los taxa parafiléticos son, sin embargo, muy utilizados, no solo en palabras comunes, sino también en escritos científicos.
Ejemplo:
Los sistemáticos Filogenéticos utilizan un método HIPOTETICO-DEDUCTIVO basado en la comparación de los patrones de distribución de caracteres dentro de taxa vivientes y, subsecuentemente, fósiles; ellos buscan patrones congruentes y proponen una hipótesis de relaciones filogenéticas basada en la DISTRIBUCION CONGRUENTE DEL MAYOR NUMERO DE SINAPOMORFIAS, dicha hipótesis es considerada como la más parsimoniosa, hasta que es refutada por una hipótesis de mayor parsimonia (han seguido el descubrimiento de nuevos caracteres). La historia real de un grupo es desconocida y solo puede ser aproximada cada vez por una sucesión de hipótesis y refutaciones. El uso de PARSIMONIA como criterio para preferir una hipótesis de relación filogenética a otra, es la escogencia de la hipótesis accesible a REFUTACION o FALSIFICACIÓN. A este respecto la hipótesis Hennigiana de relaciones filogenéticas, expresada por un cladograma o árbol filogenético, es consistente con la definición de Popper sobre hipótesis científicas (Popper, 1959, 63, 72; Bock, 1974; Platnick & Gaffney, 1977, 78a, b) y esta aproximación a la filogenia está conforme con el método de Bernard (1865) en investigación científica. Actualmente existe un fuerte debate sobre el uso del principio de parsimonia en la reconstrucción filogenética. En síntesis una clasificación Hennigiana puede ser considerada como una PREDICCION que puede ser refutada por un nuevo patrón de distribución de caracteres.
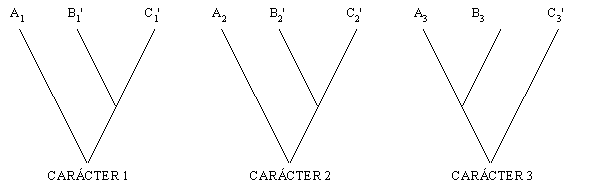
El análisis de tres caracteres homólogos 1, 2 y 3 en tres taxa A, B y C, puede llevar a hipótesis conflictivas sobre las relaciones entre estos taxa, de acuerdo a los caracteres bajo consideración. Los caracteres 1 y 2 muestran patrones congruentes, con A siendo grupo hermano de B + C, mientras que el carácter 3 lleva a una hipótesis diferente, con C siendo grupo hermano de A + B
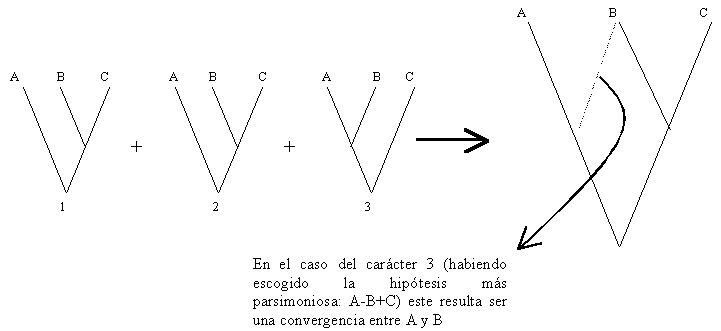
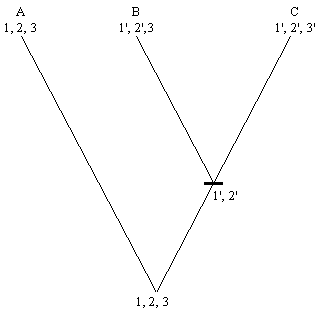
La investigación de otro carácter (4) puede revelar el mismo patrón para el carácter 3. Se tendría, por lo tanto, dos hipótesis con el mismo número de caracteres congruentes, y no se tiene criterio para escoger. Esta incertidumbre es expresada por un cladograma no resuelto, que muestra una tricotomía.
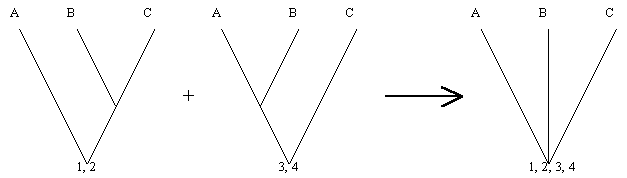
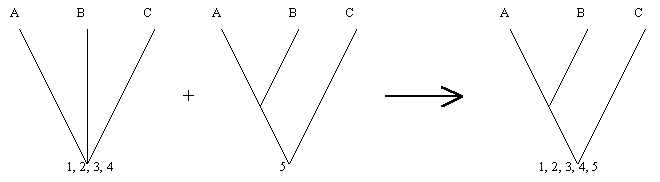
Dos especies o grupos de especies mas estrechamente relacionadas una a otra que a cualquier otra especie, en una clasificación filogenética, son llamados ESPECIES HERMANAS O GRUPOS HERMANOS, y poseen un mismo rango taxonómico, aunque, sin embargo, muchos de ellos difieran en divergencia o importancia en número de especie. Por los tanto, cuando una o más especies son el hermano de un grupo muy grande, esta debe ser colocada en un taxón supraespecífico, cuyo rango sea igual al del grupo mayor. Por ejemplo, los Monotremata o Prototeria (Ornitorrincos y Equidna), representados solo por seis (6) especies vivientes, son el grupo hermano de Theria (Marsupiales y Placentados), representado por más de 5000 especies y poseen el mismo rango taxonómico en la clasificación filogenética: Cada grupo representa una subclase.
En respuesta al criticismo de los taxónomos evolutivos, quienes consideran que la clasificación filogenética puede ser imposible de usar debido al número excesivamente grande de categorías (Rangos: en principio uno para cada dicotomía del cladograma), los cladistas han propuesto varias convenciones para simplificar las clasificaciones filogenéticas. Dichas clasificaciones resultan ser las MINIMAMENTE REDUNDANTES (Wiley, 1979). Por ejemplo, cuando un taxón que contiene una sola especie recibe un alto rango taxonómico debido a que es hermano de un taxa de rango igualmente alto, no es necesario crear una sucesión de taxa supraespecíficos para esta especie.
La SECUENCIA FILOGENETICA (Nelson, 1974) es otra convención de acuerdo a la cual los taxa terminal de un cladograma asimétrico tienen el mismo rango de los demás taxa terminales. Estos taxa son listados, en una clasificación, en orden a su secuencia de ramificación. Dicha clasificación puede reflejar exactamente el cladograma que evita nuevos nombre para cada dicotomía.
Finalmente Patterson y Rosen (1977) propusieron el término PLESION como un rango no específico para los grupos monofiléticos fósiles que son insertados en una clasificación basada en grupos extintos. Con esta convención, una clasificación filogenética basada en grupos extintos no necesita ser modificada por el descubrimiento de un nuevo grupo monofilético fósil.
Falta un tercer aspecto en el método de Hennig, junto al uso exclusivo de sinapomorfías en la reconstrucción filogenética y el uso exclusivo de relaciones filogenéticas para establecer clasificaciones, Hennig propuso un criterio para establecer el ESTADO DE LOS CARACTERES (Apomórficos y Plesiomórficos), el mayor criterio es el "esquema de argumentación" mejor conocido actualmente como el OUT-GROUP o REGLA DEL OUT-GROUP: cuando se desean analizar caracteres en los miembros de un grupo monofilético, se deben buscar los homólogos a estos en su grupo hermano. Si se presentan estos homólogos, resultan ser plesiomórficos para el grupo monofilético en consideración; pero si el homólogo está ausente en el grupo hermano y es único al grupo monofilético bajo estudio, entonces es apomórfico a este.
Ejemplo: Los cinco dedos de los mamíferos son un carácter plesiomórfico, debido a que también están presentes en su grupo hermano tradicional, los Diápsidos (Lagartos, serpiente, pájaros y cocodrilos), aunque ha ocurrido una reducción secundaria aquí y allá (Aves). Este test de OUT-GROUP puede ser reforzado con comparaciones con las Tortugas (Consideradas por algunos filogenetistas como el grupo hermano de los Mamíferos + Diápsidos) o los Lisanfibios (Grupo hermano de todos los amniotas). Por lo tanto, los cinco dedos no son solo plesiomórficos par los mamíferos, sino también para todos los amniotas. Sin embargo, la comparación OUT-GROUP con el grupo hermano de los Tetrápodos, los peces pulmonados, muestra que este es apomórfico a los últimos (los Peces no tienen cinco dedos).
La regla del OUT-GROUP ha estado bajo una fuerte polémica recientemente (Nelson, 1978) por lo que debe ser utilizada cuidadosamente.
Por fortuna Hennig propuso criterios accesorios para la determinación de los estados de carácter: El Orden Geológico de Sucesión de Caracteres, su Sucesión Biogeográfica, Correlación de Transformación de Caracteres Ontogenéticos. El primer criterio accesorio de ARGUMENTO PALEONTOLOGICO ha sido criticado por Nelson (1978), quien lo rechaza como irrefutable y por ser un simple "ídolo de la academia". El mismo Hennig (1966) consideró que este criterio solo podía ser usado cuando todos los otros criterios hubieran fracasado. Usualmente, pero no siempre, los caracteres del representante más antigua, geológicamente, de un grupo son más primitivos que sus homólogos existentes. Archaeopterix, el ave más antigua conocida, muestra características primitivas (como los dientes).
El criterio accesorio secundario, también llamado REGLA DE PROGRESION, está relacionado con la Biogeografía. Hennig (1966) supuso que el taxa más primitivo de un grupo podía ser encontrado más cerca del centro de origen del grupo que de los más derivados. Esta regla ha sido discutida por Nelson (1975), Brundin (1981) y Slater (1981) y actualmente está mejor entendida gracias a la VICARIANZA BIOGEOGRAFICA, más que por el ajuste dispersalista ideado por Hennig, a partir de la Biogeografía Darwiniana.
El tercer criterio accesorio, o criterio de CORRELACION DE TRANSFORMACION DE SERIES, consiste en la comparación sucesiva de estados de carácter de diferentes series de transformación. Por ejemplo, si dos caracteres, o más, muestran modificaciones que se incrementan regularmente en una serie de taxa existentes, esto puede ser considerado como una indicación de la polaridad de otros caracteres en el mismo taxa. Este criterio es simplemente una extensión del test de HOMOLOGIA CONGRUENTE (Patterson, 1982b), apoyado por los cladistas modernos - Cuando una mayoría de patrones de distribución homologa (= sinapomorfía) son congruentes, la probabilidad de que cada patrón proporcione una buena hipótesis de polaridad de estados de carácter, es mayor; sin embargo para Wiley (1981), la transformación de series de dos caracteres es más interesante cuando estos chocan, ya que esto indica que algo está mal en el análisis.
Finalmente se tiene el criterio ONTOGENETICO, que es el orden de aparición de los caracteres o estados de carácter, durante la ontogenia de un organismo. Actualmente es considerado por Lovtrup (1974, 1978), Nelson (1978) y Patterson (1982b) como más que un criterio accesorio. Está mas cerca de la visión de Von Baer (1828), aunque no es idéntica a esta. De acuerdo con Von Baer (1828) los caracteres mas generales aparecen tempranamente en el desarrollo de un individuo, mientras que los más particulares lo hacen después. Por lo tanto, los caracteres más Apomórficos son aquellos que son más particulares y que están restringidos al mínimo número de organismos. El criterio ontogenético es considerado, ahora, como el único confiable por los "cladistas Transformados".

