|
First Page | Habituation | Sensitization | Classical Conditioning | Somatotopic Map Plasticity | Implications
|
|
The Somatotopic Map in the Brain is
Modifiable by Experience. |
|
The structure of columns concerned with ocular dominance in area 17 of the cerebral cortex can be greatly
modified by experience during an early critical period. If one eye is closed during the peak of the critical period, even for only one week, the columns devoted to that eye shrink and those devoted to the open eye expand. This ability to modify ocular dominance columns is restricted to a relatively short period just after birth, but it raises an intriguing question: To what degree can altered sensory experience in later life produce changes in the architecture of the brain - in the size of cortical columns, or even in the precise details of the various sensory and motor maps? The work we have just reviewed in simple animals indicates that learning produces structural and functional changes in specific nerve cells. In mammals, and especially in humans, in whom each component of function is represented by hundreds of thousands of nerve cells, learning is likely to lead to alterations in many nerve cells and is therefore likely to be reflected in changes in the basic pattern of interconnections characteristic of the various sensory and motor systems. This is indeed what appears to be happening. The most detailed evidence has come from studies on the somatic sensory system. |
|
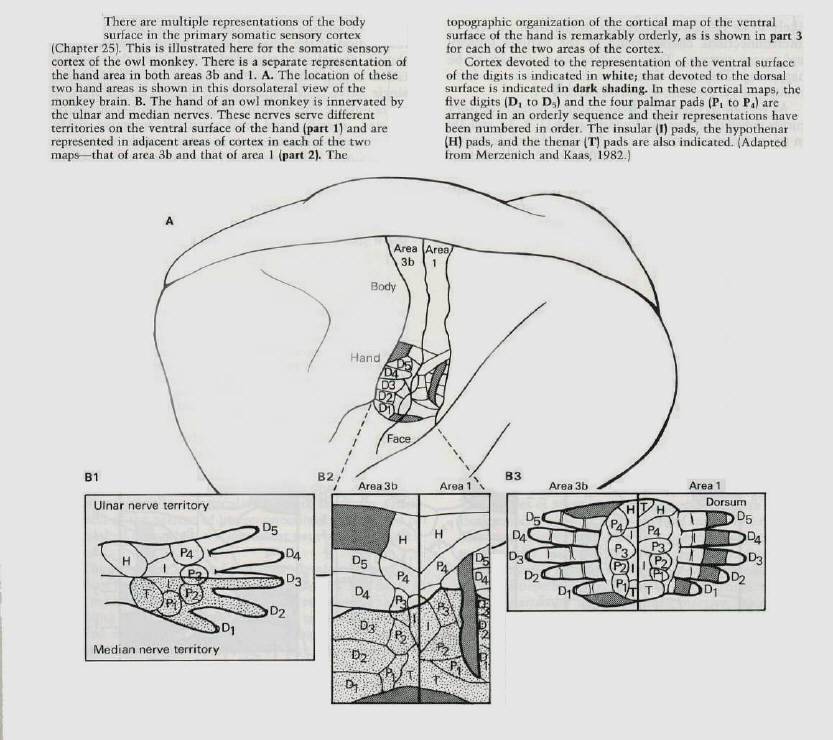
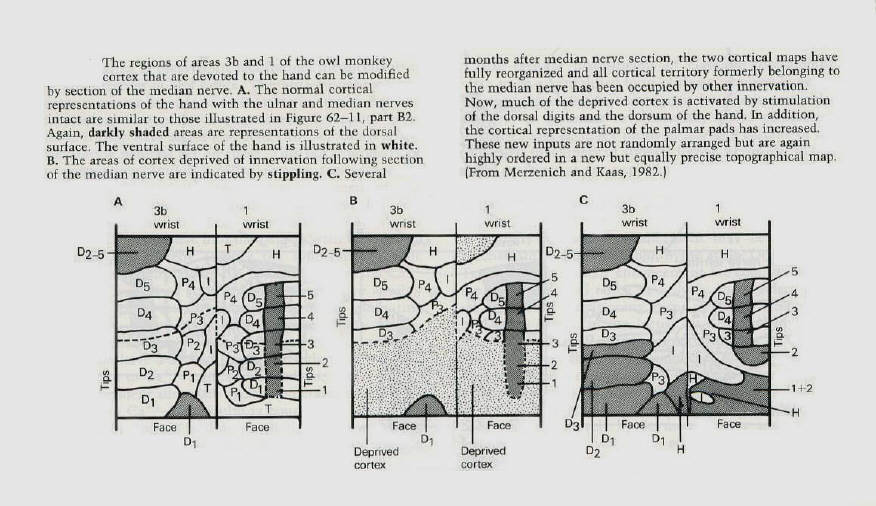
The primary somatic sensory cortex consists of four Brodmann's areas (1,2,3a and 3b) in the postcentral
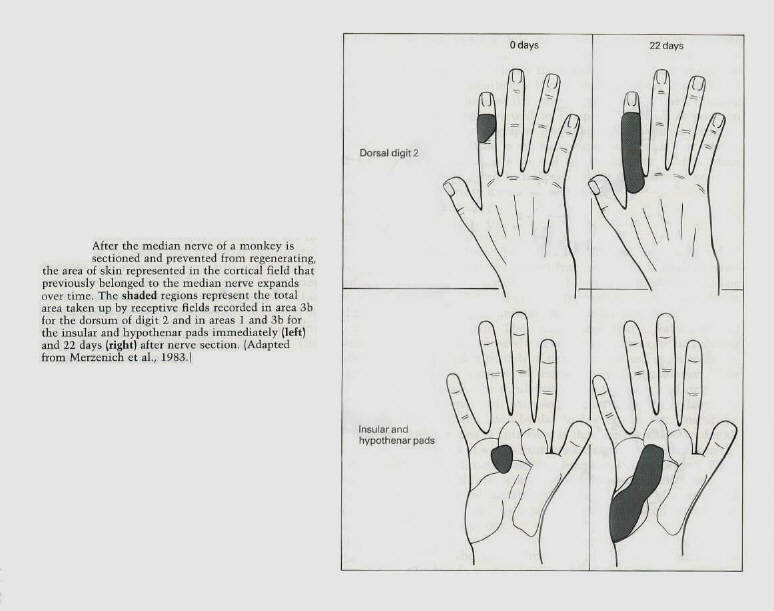
gyrus. Each of these areas represents a separate map of the body surface (figure 11). Michael Merzenich and J.H. Kaas at the University of California in San Francisco recently found that the cortical maps can change in a systematic and use - dependent manner. They first encountered this phenomenon by exploring the recovery of function after nerve injury in monkeys. They severed the median nerve in the hand, which innervates the cutaneous receptors on much of the ventral surface of the hand, the palm and the glabrous portions of digits 1,2, and 3 (figure 11). After cutting the nerve, one would expect those cortical areas committed to denervated parts of the hand to be unresponsive and silent. However, when the cortex was remapped even immediately after denervation, only a portion of the territory devoted to the median nerve was unresponsive. Merzenich and Kaas found that the adjacent cortex had expanded its influence into the denervated cortex: a significant part of the cortical area of the median nerve could now be activated by stimulating the neighboring parts of the hand outside the territory of the median nerve. For example, the region formerly activated by the ventral (palmar) surface of the thumb (digit 1) was now activated by the dorsal (hairy) surface of that finger. Thus, a fragmentary representation of the adjacent region of the dorsum of the thumb immediately expanded and replaced the representation of the ventral surface (figure 12). Moreover, single nerve cells responding to stimuli on the hand in areas outside the territory of the median nerve had restricted, specific, and well-organized receptive fields. These single-cell recordings revealed that the dorsal surface of finger 2 that previously had a small representation in the median nerve territory of the cortex had a much larger territory several weeks after the median nerve had been cut. Similarly, areas on insular pads that previously had a modest representation expanded substantially in the weeks after sectioning of the nerve (figure 13). |
|
These findings indicate that the organization apparent in the maps of the somatic and perhaps all sensory
systems reflects only part of the total pattern of anatomical connections - the maps reveal only the dominant pattern of organization in a particular region of the brain. Other aspects of the pattern of interconnections are revealed or unmasked only when the dominant pathways are inactivated.
The map that is found immediately after cutting the median nerve changes further over the next few days. If
the median nerve is not allowed to regenerate, the remaining silent parts of the cortex are taken over and occupied by adjacent normally innervated digits and by parts of the hand innervated by the ulna and radial nerves. Within 3 weeks after nerve section almost all the former cortical territory of the median nerve is reoccupied (figures 12 and 13). |
|
Changes in the Somatotopic Map Produced by Learning May Contribute to
the Biological Expression of Individuality. |
|
The studies of Merzenich and Kaas demonstrate that cortical somatic sensory maps are not static, but
dynamic. The maps can expand into functionally vacated sectors to represent the bordering skin regions in finer grain. These changes suggest that even in adult monkeys there is a use-dependent competition for cortical territory. Once a particular input becomes inactive, its representational territory can be captured by inputs from adjacent, normally innervated skin.
The reorganization is manifested in the cortex, but it is likely that the primary change is at a lower level in the
brain. As first illustrated by Patrick Wall and David Egger at University College London, reorganization occurs at least in part at the level of the dorsal column nuclei, which contain the first synapses of the somatic sensory system. Organizational changes are therefore probably a general property of the system and occur throughout the somatic afferent pathway. The fact that anatomical changes occur so early in sensory processing suggests that the substrates of higher mental processing are also labile and capable of influence by experience.
The cortical maps of an adult, and indeed the whole adult sensory system, are probably subject to constant
modification on the basis of use or activity of the peripheral sensory pathways. Since all of us are brought up in somewhat different environments and use our bodies and senses differently, it is clear that the architecture of our brains will be shaped in unique ways. This architecture, in turn, will interact importantly with our unique genetic constitution to shape the biological basis and therefore the psychological expression of our individuality.
Two further studies by Merzenich have provided evidence consistent with this view. First, he studied normal
animals and found that the topographical maps vary considerably from one animal to another (figure 14). This study, of course, does not separate the effects of different experiences from the consequences of different heredity. Merzenich and his colleagues therefore investigated the factors that underlie this variability in a second series of experiments. They encouraged monkeys to use their middle fingers at the expense of other fingers in the hand by having the monkey press a rotary disc for food with only these fingers. After several thousand bar presses, the area in the cortex devoted to the middle fingers was greatly expanded. These findings suggest that such practice acts on pre-existing patterns of connections and strengthens their effectiveness. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}