|
First Page | Habituation | Sensitization | Classical Conditioning | Somatotopic Map Plasticity | Implications
|
|
Sensitization Involves an Enhancement
of Synaptic Transmission. |
|
Sensitization is a more complex form of learning than habituation: it is the enhancement of an animal's reflex
responses as a result of the presentation of a strong or noxious stimulant. In contrast to habituation, sensitization causes an animal to pay attention to a variety of stimuli, even previously innocuous ones, because they are potentially accompanied by painful or dangerous consequences. Sensitization, like habituation, can last from minutes to days and weeks, depending on the pattern of stimulation. |
|
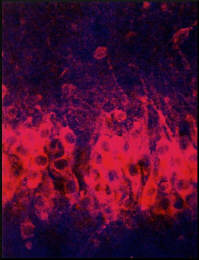
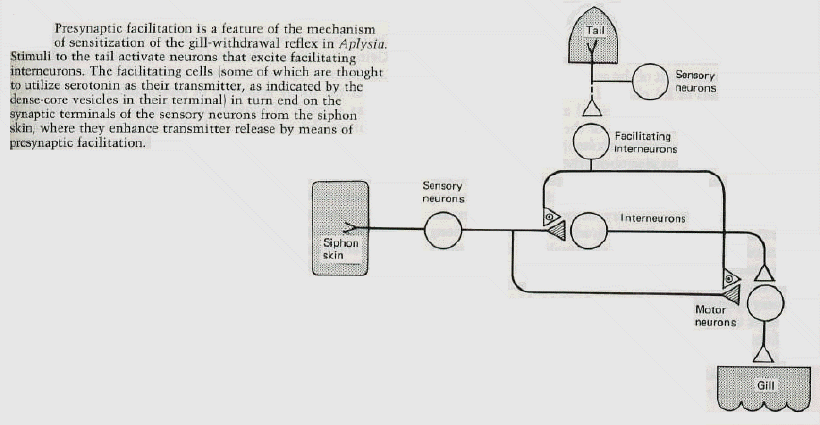
At the cellular level, sensitization in Aplysia also involves an
alteration of synaptic transmission at the synapses made by the sensory neurons on the motor neurons and interneurons. The same synaptic locus can therefore be regulated in opposite ways by opposing forms of learning: its activity can be depressed by habituation and enhanced by sensitization. In sensitization, however, another mechanism comes into play. This mechanism is presynaptic facilitation, which is mediated by an axo-axonic synapse, a synapse on a synapse (figure 3). The sensitizing stimuli activate a group of facilitating interneurons, which synapse on or near the terminals of the sensory cells. These facilitating neurons enhance transmitter release from sensory neuron terminals by causing an increase in the amount of cyclic adenosine 3', 5'-monophosphate (cyclic AMP) in the sensory neurons. Because experimental application of serotonin mimics the actions of the facilitating interneurons and the natural sensitizing stimuli, and because the terminal regions of the sensory neurons receive serotonergic innervation, some of the facilitators are thought to be serotonergic. |
|
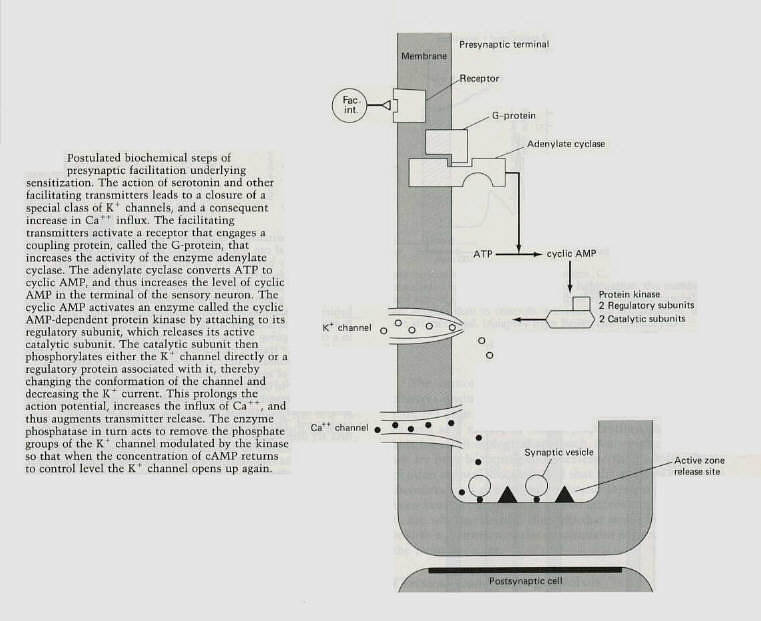
On the basis of pharmacological and biochemical studies, Klein, Castellucci, Robert Hawkins, Steven Siegelbaum,
Lise Bernier, James Schweartz and Kandel have pieced together the likely sequence of biochemical steps that occur as a result of sensitization (figure 5). According to this model, serotonin, which is thought to be released by some of the facilitating neurons in the gill and siphon withdrawal reflex, activates a serotonin receptor in the membrane of the presynaptic terminal of the sensory neuron. The serotonin receptor engages a coupling protein (G-protein), which in turn activates an adenylate cyclase. Stimulation of the adenlyate cyclase increases the concentration of cyclic AMP within the terminal. Cyclic AMP then activates a protein kinase - an enzyme thought to be the common site of action for cyclic AMP in eukaryotic cells. Protein kinases add phosphate groups to proteins, thereby changing their charge and consequently their shape. This process, call phosphorylation, can lead to either an increase or a decrease in the activity of a protein. In the case of sensitization the activated protein kinase has been shown to phosphorylate a novel K+ channel protein (the serotonin sensitive K+ channel) or a protein that is associated with it. This serotonin-modulated K+ channel has been found to participate selectively in sensitization. Phosphorylation of this channel or associated proteins reduces a component of the K+ current that normally repolarizes the action potential. Reduction of this current prolongs the action potential and thereby allows Ca++ channels to be activated for longer periods of time.
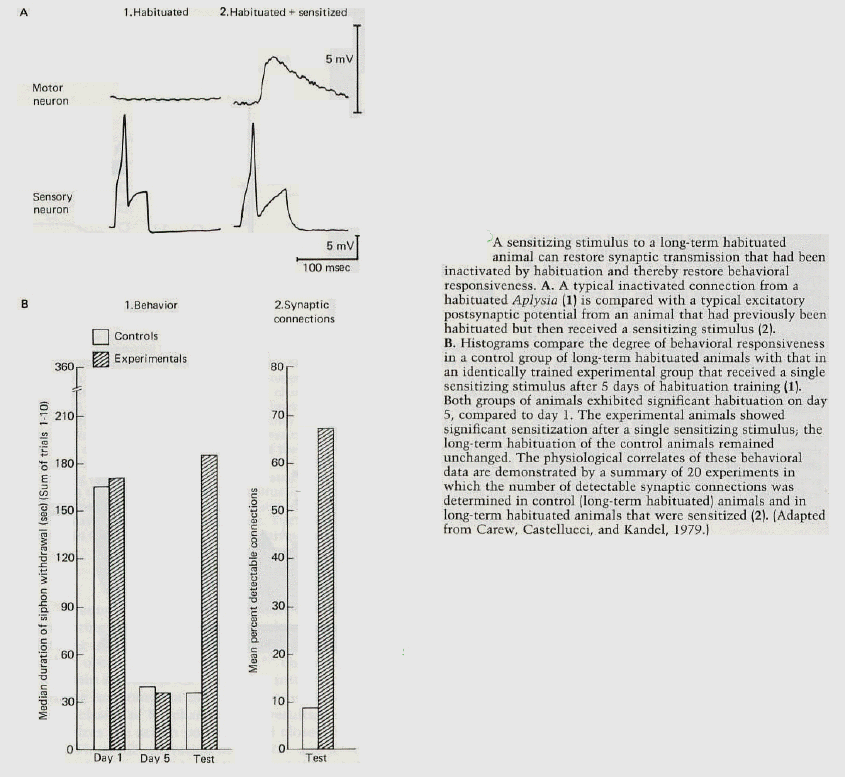
Sensitization can reverse the synaptic depression of habituation. Sensitization is an effective form of learning; it so
enhances behavioral responsiveness that it can reverse the synaptic and behavioral depression that occurs not only in short-term but even in long-term habituation (figure 6). Thus, synaptic pathways, determined by innate genetic and developmental processes, can be funcitonally interrupted and then functionally restored (or dishabituated) by simple learning experiences! The training procedures that produce this learning are relatively modest, comparable to the social experience of one person speaking to another. By extension from what we know of the mechanisms of simple forms of learning, we may assume that when two people speak, no only do they make eye contact and voice contact, but through behavior the actions of the neurons in the brain of each person have a direct and long lasting effect on the functioning of the neurons in the brain of the other. |
|
Long-term Habituation and Sensitization Produce
Morphological Changes. |
|
Habituation and sensitization are learning processes that can turn the synapses of specific neurons off and on. In
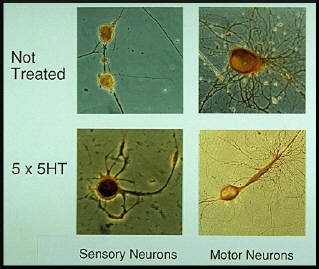
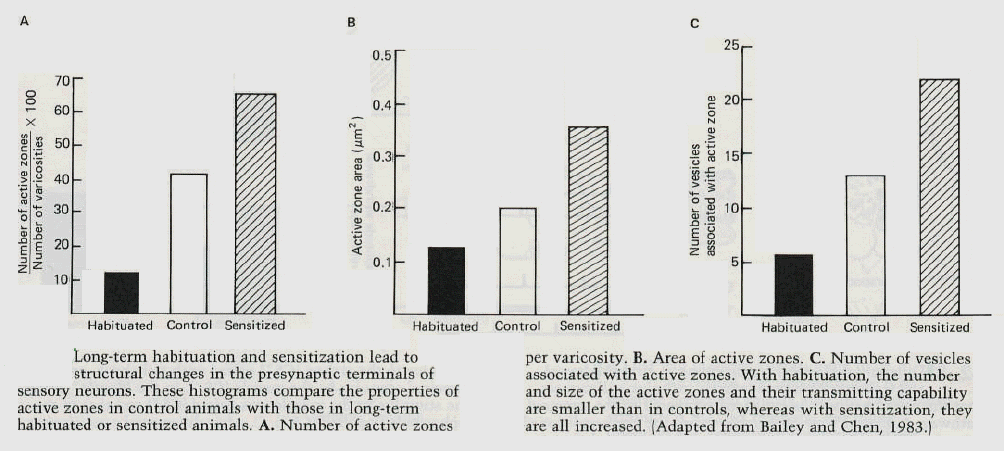
long-term learning, is this functional change accompanied by a morphological change? To answer this questions, Craig Bailey and Mary Chen at the College of Physicians and Surgeons of Columbia University have carried out a morphological analysis on the sensory neurons of the gill-withdrawal reflex in Aplysia. They injected the sensory neurons with the electron dense marker horseradish peroxidase and visualised their synaptic terminals with the electron microscope. This technique enabled investigators to compare synapses that have undergone long-term learning with control synapses. In particular, they analyzed changes both in the number and distribution of synaptic vesicles (the likely storage sites of transmitter quanta) and in the size and extent of active zones (the sites from which transmitter is actually released). Active zones contain arrays of submembranous dense particles where the vesicles are positioned and their contents released. The active zones are located in varicose expansions of the axonal processes. However, not all varicosities contain active zones. In sensory neurons from untrained animals, only 40% of terminals have active zones, the rest do not. Long-term habituation reduces the presence of active zones to only 10% of the terminals. Moreover, even the few remaining active zones are smaller than in control varicosities. In contrast, sensitization increases the incidence of active zones to 65% of terminals, and the average size of each zone is larger than in control animals (figure 7). |
|
Cell-Biological Studies of Habituation and Sensitization Have Provided Some
Basic Insights into the Mechanisms of Learning and Memory. |
|
Several points emerge from a consideration of these simple examples of learning and memory. One is that both
short and long term memory can be localized to particular sites. In these instances, at least, memory is not dependent on widely distributed processes. These memories are also not dependent on reverberating activity in closed neuronal loops. Second, learning need not involve gross anatomical rearrangement in the nervous system. No nerve cells or even synaptic pathways are created or destroyed; rather, learning of habituation and sensitization results from changes in the effectiveness of previously existing chemical synaptic connections. Finally, learning need not depend on special "memory" neurons - or by extrapolation, memory centers in the brain - whose only function is to store information. Learning can result from changes in neurons that also act as integral components of a normal reflex pathway. It is therefore likley that different types of experiences may be stored in a large number of different cells that are involved in a variety of other functions. Learning itself is not necessarily an independent process; rather, it can be a modification of other, already existing processes.
The demonstration of the molecular and structural changes during even these simple forms of learning has
required a variety of techniques developed only recently, as well as a variety of simple preparations that are suitable for a cell-biological approach. No wonder, then, that we are only beginning to understand the biological basis of other mental processes and that the analysis of mental disorders such as schizophrenia and depression has so far been beyond our reach. It will be interesting in the future to see whether mental illnesses that result from learning lead to a distortion of the connections in specific areas of the brain. |
{kind=link}
{kind=link}
{kind=link}
{kind=link}