|
First Page | Habituation | Sensitization | Classical Conditioning | Somatotopic Map Plasticity | Implications
|
|
Classical Conditioning Involves an
Activity-Dependent Enhancement of Presynaptic Facilitation. |
|
Classical conditioning is a slightly more complex form of
learning than sensitization because the subject is not concerned with learning about the properties of one stimulus (the sensitizing stimulus) but must learn a temporal association between two stimuli. In classical conditioning an initially weak or ineffective conditioned stimulus becomes highly effective in producing a behavioral response after it has been paired in time with a strong unconditioned stimulus. The timing is critical to the outcome. |
|
For classical conditioning to work, the conditioned stimulus must precede the unconditioned stimulus and it must
usually do so by a critical interval of about .5 to 1.0 sec. This form of learning is therefore noncommutative. It does not work if the conditioned stimulus follows the unconditioned stimulus. Classical conditioning is fascinating because, as we saw earlier, it represents the learning of a fundamental causal relationship. When an animal has been conditioned, it has learned that the conditioned stimulus predicts unconditioned stimulus. Thus through classical conditioning, an organism is able to discriminate cause and effect relationships in the environment. In contrast, nonassociative learning such as sensitization does not require temporal pairing of stimuli and does not teach the animal to expect any relationship between stimuli. Often a reflex can be modified by sensitization as well as by classical conditioning. In such cases, the response enhancement produced by classical conditioning is greater or lasts longer than the enhancement produced by sensitization. Moreover, whereas sensitization indiscriminately affects a broad range of defensive responses, classical conditioning enhances only a restricted range of responses appropriate to the pairing of the conditioned stimulus and the unconditioned stimulus.
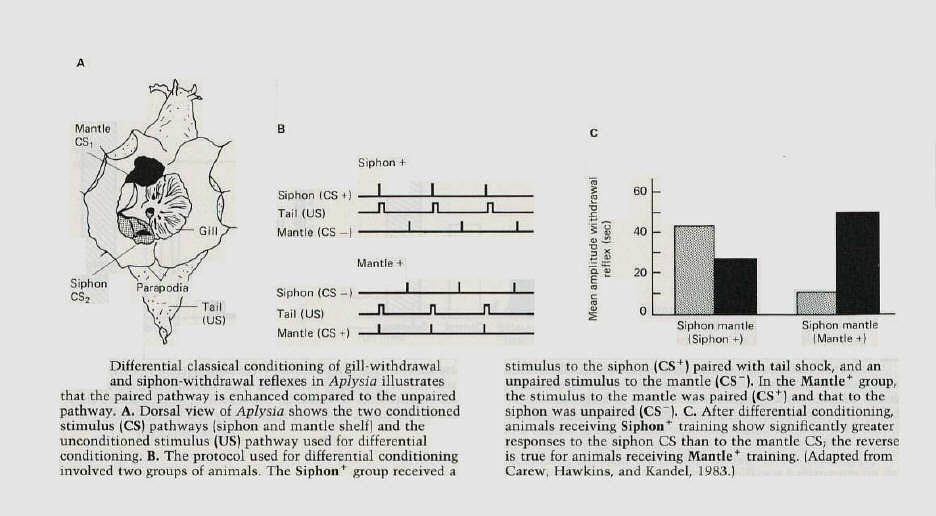
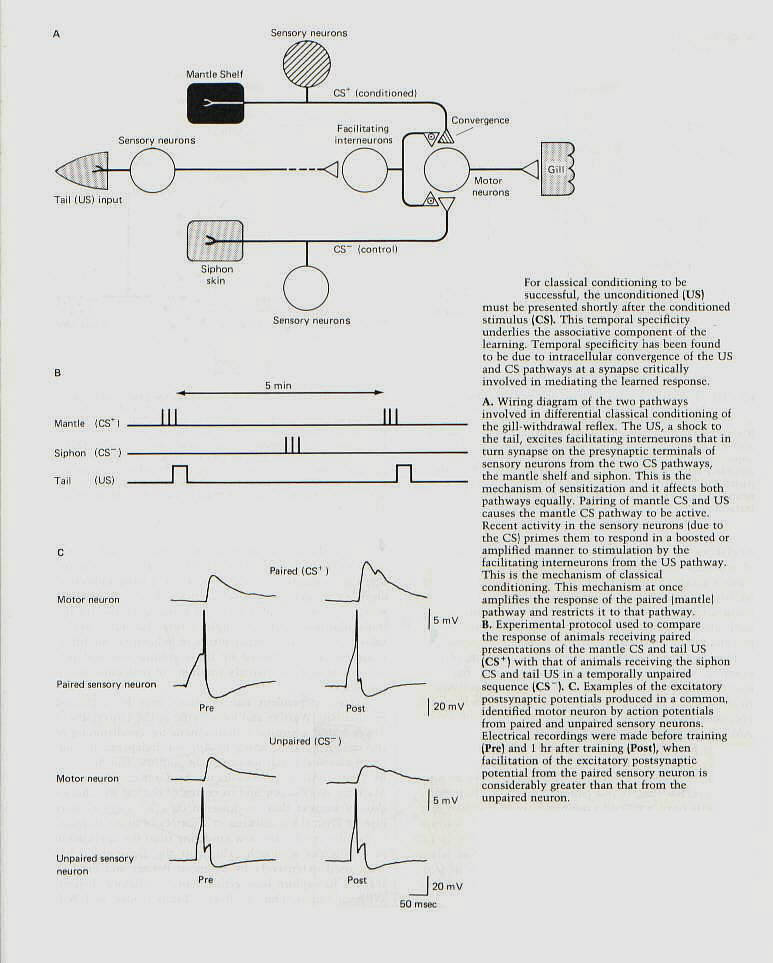
The gill and siphon withdrawal reflex of Aplysia can be enhanced by classical conditioning as well as by
sensitization. Moreover, the withdrawal reflex is capable of being differentially conditioned. The withdrawal can be elicited by stimulating not only the siphon but also a nearby structure called the mantle shelf. Each of these areas is innervated by its own population of sensory neurons, and each pathway can be activated independently to serve as a conditioned stimulus (figure 8-A). These pathways can be differentially conditioned by pairing the unconditioned stimulus (a strong shock to the tail) with stimuli to either the siphon or the mantle shelf and stimulating the other pathway in a way that is unpaired with the tail shock (figure 8-B). After such training, the response to stimulation of the paired site is significantly greater than the response to stimulation of the unpaired site (figure 8-C). |
|
Unlike nonassociative learning, associative learning requires temporal specificity - a specific timing of stimuli. What
then are the cellular mechanisms whereby this temporal pairing of stimuli leads to learning that is both more robust than nonassociative forms and more restricted in its distribution (being limited to the paired pathways)? Temporal specificity results from a convergence of the conditioned and unconditioned stimuli at the level of individual sensory neurons in the conditioned stimulus pathway. Greater presynaptic facilitation of the sensory neurons - greater depression of the serotonin-sensitive K+ channel leading to greater influx of Ca++ and more transmitter release - occurs in response to the unconditioned stimulus when the sensory neurons have just been active because of exposure ot the conditioned stimulus (figure 9). Thus, the facilitation of the connection from a sensory neuron to a follower neuron is amplified if-and only if- the sensory neuron is active just before the arrival of the input from the facilitating neurons that are activated by the unconditioned stimulus. Activity in the sensory neurons that follows the unconditioned stimulus has no effect. Moreover, for facilitation to be amplified, activity in the sensory neurons must precede the unconditioned stimulus by a brief critical period. We have seen in our discussion of neuronal modulation during development and critical periods that activity is often an important step in the fine tuning of synaptic connections. Here we can see quite clearly that appropriately timed activity is also important for learning in the mature organism.
These experiments indicate that the mechanism of classical conditioning of the withdrawal reflex is actually an
elaboration of presynaptic facilitation, the mechanism of sensitization of the reflex. The sensory neurons undergo a greater amount of facilitation in response to the facilitating neurons or to experimentally applied serotonin if they were active just before the serotonin or the facilitating neurons act on them. This novel propety of presynaptic facilitation is called activity dependence. |
|
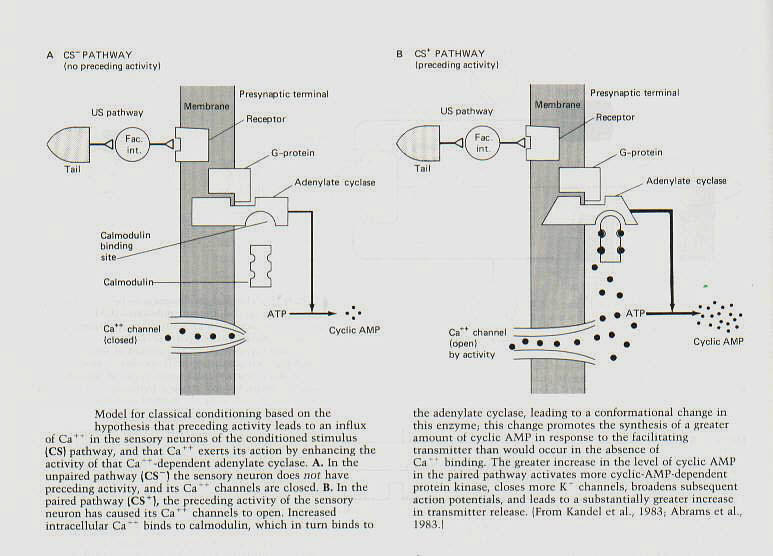
How is this activity-dependent enhancement of presynaptic facilitation achieved? One of the consequences of
activity is to allow Ca++ to move into the neuron with each action potential and thereby amplify the synthesis of cyclic AMP by serotonin (figure 10): specifically, the Ca++ influx is thought ot act through calmodulin to amplify the activation of the adenylate cyclase by the facilitating transmitter. Indeed, most cells of the body have an adenylate cyclase that is independent of Ca++. But many brain cells have a second type of adenylate cyclase that is sensitive to Ca++ - calmodulin, and htis Ca++ -calmodulin-dependent adenylate cyclase generates more cyclic AMP when Ca++ -calmodulin is bound to it than when it is not. In Aplysia (and as we shall see below in Drosophilia as well) this second form of adenylate cyclase might be important for classical conditioning. |
|
The cellular mechanism of classical conditioning seem to be an
amplified form of those involved in sensitization, suggesting that there may be a basic molecular alphabet or grammar to elementary forms of learning whereby more complex forms of learning partake of the molecular machinery of simpler forms. According to this view, a variety of distinct forms of behavioral modifications could be achieved by the combinations and permutations of a surprisingly small set of molecular building blocks. |
|
Activity-dependent enhancement may be a general mechanism. Walters and John Byrne at the University of Texas
found a similar enhancement for conditioning of the tail-withdrawal reflex in Aplysia. Independent studies of classical conditioning in the mollusc Hermissenda by Daniel Alkon, in the locust by Graham Hoyle and Marjorie Woollacott and in cortex of the cat by Charles Woody suggest that modulation of a K+ channel may also be critical for learning in a variety of other animals.
Similar results are now emerging from the application of a different approach. The fruit fly, Drosophilia, has been
used extensively by Seymour Benzer and his colleagues to explore how genes control behavior. Benzer, William Harris, Duncan Byers, Yadin Dudai, and William Quinn have found that the fruit fly can be classically conditioned. Using genetic selection procedures, Quinn, Dudai, and Byers then isolated single-gene mutants that could not learn. Four of these mutants have now been studied in detail and have two interesting features. First, in addition to being unable to learn classical conditioning, some of these mutants cannot learn sensitization. Second, all of the mutants have a defect in the cyclic AMP cascade. One lacks one type of cyclic AMP phosphodiesterase, the enzyme that degrades cyclic AMP. As a result, the fly has abnormally high levels of cyclic AMP that are thought to be out of the range of normal modulation. Other learning mutants have defects in the serotonin receptor, the biogenic amine transmitters, or the Ca++ -calmodulin dependent adenylate cyclase. Thus, both cell-biological approaches in Aplysia and genetic approaches in Drosophilia indicate that the cyclic AMP cascade is important in elementary forms of learning and memory storage. |
{kind=link}
{kind=link}
{kind=link}