DNA-PK (DNA-dependent protein kinase) 主要是由一個具有催化活性的次單元體DNA-PKcs 以及一個具有結合DNA 能力而調控其激活性的次單元體Ku 所組成,最近幾年的研究,發現DNA-PK 是雙股DNA 斷裂(DNA Double-Strand Break; DSB)的修補及產生抗體多樣性V(D)J 重組[V(D)J recombination]作用中的重要成員之一。除此之外,DNA-PK 在基因的轉錄調控與核染色質 (Chromatin)及端粒(Telomere)結構上的維持也具有相當的重要性。
隨著物質文明的進步,生物體本身基因的完整性極易遭受一些外在環境的因素所影響,輻射線的傷害或是一些化學試劑的破壞都會造成DNA 的 DSBs (Double-strand breaks),DSBs 也會在免疫球蛋白質的位置特一性V(D)J 重組過程中產生。 假若DNA 的DSBs 無法修補或是不正確的修補都將會使染色體產生變異導致細胞的癌化或是死亡,因 此對真核細胞來說DSBs 的修補與維護保有基因的完整性是非常重要的,其中最主要的修補機制就是 NHEJ (Nonhomologous end-joining),這種修復機制的特性是兩頭的重組DNA 分子並不需要互補的 DNA 序列。
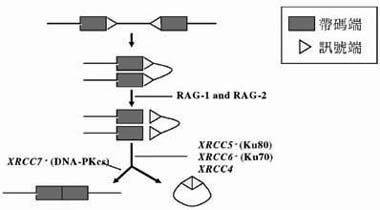
在免疫細胞的分化和發育上,NHEJ 在抗體多樣性的V(D)J 重組作用亦扮演極為重要的角色,經由V(D)J 重組可產生多樣性的抗體和T 細胞接受子以認識及對抗外來的抗原。所謂的V(D)J 重組乃是由任一V (Variable)基因片段和任一的D (Diversity) 或J (Joining)基因的片段隨機連結在一起以解譯成可認識抗原的功能區。簡單來說,V(D)J 重組作用可分成兩階段,第一、兩個淋巴細胞特有的蛋白質,RAG1 和RAG2 (重組活化基因)認識 RSS (重組訊號序列)於V ,D 和J 基因兩側,進行雙股 DNA 斷裂(DSB),產生齊頭訊號端(Signal ends)和髮夾狀帶碼端(Hairpined coding ends)。第二、經由NHEJ 的作用,齊頭訊號端和髮夾狀帶碼端分別連結在一起。目前已知參與連結的蛋白質和NHEJ 的DNA DSB 修復為相同的蛋白質。有四個重要的互補分佈群組(Complementation)在NHEJ 中扮演重要的角色,分別是XRCC4, XRCC5, XRCC6 及 XRCC7 。XRCC4 與DNA Ligase IV 共存在同一 多蛋白質複合體中,XRCC4 可以增加DNA Ligase IV 的活性。而XRCC5, XRCC6 及XRCC7 則分別代表Ku80, Ku70 及DNA-PKcs 的基因。因此, DNA-PK 在NHEJ 中也扮演一個極為重要的角色。
在許多動物及細胞的模式實驗中,已顯示出 DNA-PK 在V(D)J 重組的重要性。SCID (Severe combined immunodeficiency)小鼠有DNA-PKcs 基因的突變,造成其對輻射線的高度敏感性與V(D) J 重組的嚴重缺陷;同樣地,Ku70 及Ku80 的 Knockout Mice 也對輻射線有高度的敏感性。一般認為DNA-PK 的活化模式是透過Ku 的作用,Ku 可以結合到受損雙股DNA 的斷裂處,似乎Ku 可以辨認DNA 的DSBs ,結合在DNA 的Ku 經由其立體結構的變異,促使DNA-PKcs 在DNA/Ku 的複合體上組合,以形成一個具有反應活性的多蛋白質複合體 DNA/Ku/DNA-PKcs ,此反應複合體可以促使 NHEJ 成員的組合或是會影響到一些參與NHEJ 因子的活性。此種方式,一方面可以防止受損DNA 的末 端繼續受到其他Nuclease 的作用,另一方面Ku/ DNA-PKcs 可以增加DNA 連接脢(DNA Ligase)的活性,促使受損DNA 的末端相互連接。
DNA-PK 的活性會受到其相互作用的蛋白質所影響。C1D 直接與DNA-PKcs 中的Leucine Zipper 區域相互作用,結合在DNA 的C1D 可以在DNA end-independent 的狀況下活化DNA-PK ,此種活化的模式可能是透過改變DNA 雙股螺旋的結構所導致。此外,HMG1 及HMG2 也能增加DNA-PK 的活性,因此DNA-PK 的活化可能會受到Chromatin Context 的作用所影響。另外Lyn tyrosine kinase 則以破壞DNA-PKcs/Ku 複合體的作用而調控 DNA-PK 的活性。而HSF1 藉著直接與Ku 及DNA-PKcs 的相互作用以促進DNA-PK 與DNA 的穩定性,增加DNA-PK 的活性。
此外,DNA-PK 的活性也會受到一些不同方式的後轉譯修飾(Post-translational Modification)所調控。DNA -PKcs 的自我磷酸化 (autophosphorylation)會使其與Ku 分開而抑制 DNA-PK 的催化活性;另外就是c-Abl 對DNA-PK 的調控模式,活化的c-Abl 會使DNA-PKcs 磷酸化,促使DNA-PKcs/Ku 複合體的分開而抑制 DNA-PK 的催化活性。此外,poly (ADP-ribose) polymerase (PARP)對DNA-PK 的ADP-ribosylation 會增加DNA-PK 的磷酸激活性;而在細胞的凋零過程中,DNA-PKcs 會被Caspase-3 所分解而喪失激脢活性;PKC δ 也會抑制DNA-PK 的激脢活性。
雖然DNA-PK 在DSB 的修補與V(D)J 的重組中是扮演如此重要的角色,但此重要性也僅止於在基因層次的一些證據與了解,至於DNA-PK 在這些生理作用的詳細生化特性與分子作用機制仍待進一步的研究與探討。因此鑑定DNA-PK 在生理作用的標 的將會使我們對DNA-PK 的作用機制有更進一步的了解,尤其是那些與DNA-PK 共存在多蛋白質複合體中並可以調控DNA-PK 活性的因子。
本實驗室曾經探討TIF-1b 在bAGP(a1 Acid Glycoprotein)基因表現上所扮演的角色,TIF-1b 可以和C/EBP b及GR (Glucocorticoid Receptor) 相互作用而共存在同一多蛋白質複合體中,扮演一個輔助活化子(Coactivator)促使AGP 基因的表現。此外,我們也選殖到一個同隸屬於TIF-1b這類包含PHD-finger domain (C4HC3)的新基因,除了探討此新基因在AGP 基因調控上所扮演的角色外,我們同時也發現此新基因所密碼化的新蛋白質可與DNA-PK 共存在同一多蛋白質的複合體中,此新蛋白質可以促進DNA-PK 在試管中的激脢活性;因此,我們將之命名為DPKAP (DNA-PKActivating Protein)。
DPKAP 有1189 個胺基酸,主要分佈在細胞核 中,會和DNA-PK 共存在同一多蛋白質複合體中,而且也是DNA-PK 的受脢質之一;至於DPKAP 增加DNA-PK 激脢活性的反應機制,主要是DPKAP 可以增加Ku 對DNA 的結合能力,並輔助DNA-PK 整個活化複合體在DNA 分子上的組合,同時 DPKAP 也增加DNA 與DNA-PK 多蛋白質複合體的穩定性。
因為DPKAP 在物理和生化上和DNA-PK 有密切的關聯,同時由於DNA-PK 在DNA DSB 的修復 和V(D)J 重組中扮演著如此重要的角色,因此 DPKAP 也有可能會參與V(D)J 重組作用的調節,於是我們亦探討DPKAP 在V(D)J 重組中所扮演之可能角色。初步的結果顯示DPKAP 可以同時抑制訊號端和帶碼端的連結作用。為了進一步探討此抑制作用和DNA-PK 的關係,兩個DNA-PK 的突變種, SB (1-297 胺基酸,移除C-端892 個胺基酸)和 RA-1 (487-1189 胺基酸,移除N-端的480 個胺基酸),亦用來做相同的V(D)J 重組作用分析。目前已知SB 無法和DNA-PKCS 相互結合,也不能增加 DNA-PK 激脢的活性;RA-1 則可和DNA-PKCS 相互結合。研究的結果顯示,SB 失去該V(D)J 重組的抑制作用。RA-1 仍可抑制訊號端的連結作用,但不是帶碼端的連結。因此DPKAP 在V(D)J 重組作用的調節似乎是扮演著抑制的角色,而此抑制作用可能 和DNA-PK 有關。對於DNA-PK 在V(D)J 重組上之 確切的功能區目前仍在做進一步的研究,以釐清是 否DPKAP 藉由調節DNA-PK 的活性來調控V(D)J 重組作用,抑或是經由未知的因子來調節V(D)J 重組作用。
是否DPKAP 調控DNA-PK 的激脢活性,在生物體中還隱含著其他的生理功能與意義呢?綜合言之,DPKAP 可以增加DNA-PK 在試管中的激脢活性。是否被DPKAP 所活化的DNA-PK 會使得 RPA32 (Replication Protein A)更容易被磷酸化而抑制DNA 複製(DNA Replication)的進行?是否 DPKAP 會影響DNA-PK 在基因轉錄調控的活性以及DNA-PK 在DSB 所扮演的重要角色?另一方面, DPKAP 是否會影響DNA-PK 在核染色質 (Chromatin)及端粒(Telomere)結構維持上的調控作用?這些DPKAP 在生物體內所可能隱含的生理功能以及重要性都尚待進一步的探討與深究。
參與DSB 修補的基因對V(D)J 重組的重要性:重組活化基因RAG1 和RAG2 認識重組訊諕序列(RSS)於V, D 和J 基因的兩側,進行雙股DNA 斷裂(DSB),產生齊頭的訊號端(Signal ends)和髮夾狀的帶碼端(Hairpined coding ends),再經由NHEJ 的作用,使齊頭的訊號端和髮夾狀的帶碼端各自連結在一起;而不同修補基因的變異性會對V(D)J 重組造成不一 樣的缺陷,例如SRCC7 (即是小鼠的scid locus )的基因突變會使帶碼端無法連接, XRCC4, XRCC5 及XRCC6 的基因突變會同時導致帶碼端與訊號端無法分別各自連接。
visitor 
