Hábitat y el Recurso Fruta
Es evidente que en algunos albergues encontramos diferencias significativas entre T y C, en términos de abundancia de los 5 géneros de árboles frutales más comunes, aunque no hubo un patrón consistente de un albergue y otro. Hubo diferencias significativas entre algunos albergues, después de haber agrupado los datos de T y C, particularmente en el caso de la Pona (Iriartea) y el bambú. El bambú estuvo completamente ausente de los dos albergues en el río Madre de Dios, sin embargo era relativamente común en aquellos ubicados en el río Tambopata. Aunque existen diferencias en abundancia, entre T y C y entre albergues en general, no hubo evidencia de alguna correlación simple entre un tipo de árbol y alguna especie de mamífero. Sin embargo, en terminos generales TRC tuvo la mayor abundancia de árboles frutales y una alta diversidad y abundancia de mamíferos, particularmente en terminos de primates y ungulados.
Especies con Insuficientes Datos
No se podria determinar con algún grado de confianza cual de las variables, caza o tránsito, podria ser la responsable de la variación en abundancia de 13 especies de mamífero, debido a que no se les encontró muy infrecuentemente o en un sólo albergue. Estas son: M. tridactyla, B. variegata, Dasypus sp., Aotus sp., C. albifrons, A. paniscus*, A. microtis, G. vitata, L. longicauda, L. pardalis, T. terrestris*, T. pecari*, y M. pratti. Sin embargo, algunas de estas especies (*) muestran señas de haber sido impactadas por niveles históricos de caza (es decir durante los últimos 20 años) en aquellos albergues situados cerca a comunidade o asentamientos humanos.
Especies Afectadas por la Caza
Se sabe que muchas poblaciones de fauna silvestre son comúnmente cazadas en la Amazonía por los habitantes locales bajo sistemas de subsistencia. Estudios previos han demonstrado que dicha caza puede disminuir seriamente la abundancia de las especies más perseguidas. La caza, es un impacto directo ya que es una forma de "cosecha" que disminuye una población de manera rápida. Consideramos por esta razón que es una variable de impacto mucho más poderosa que el tránsito turístico que definitivamente no actua de la misma manera y teoricamente sus efectos son indirectos en la mayoría de los casos. Hemos identificado que actualmente, la caza es frecuente cerca a dos albergues (SACHA y CAPH) e historicamente tambien ha estado presente alrededor de otros dos (ECO y EI). Es necesario, entonces, determinar que especies actualmente presentan evidencia de impacto por esta variable.
En Madre de Dios, nueve de las especies de mamíferos censados, son comúnmente perseguidos por cazadores, estas incluyen: la Maquisapa, el Machin Blanco, el Machin Negro, el Coto, la Sachavaca, la Huangana, el Sajino, el Venado Rojo, y el Añuje. Además, es claro que los felinos, tal como el Tigrillo y el Jaguar, si son encontrados, también son cazados ya que son considerados, por muchos, como peste o amenaza para el ganado y animales domésticos menores, y porque aún existe (aunque pequeño) un mercado para sus pieles. En inspecciones visuales de los IKA, de las nueve especies mencionadas, fue plenamente evidente que el Coto y el Machin Negro fueron significativamente afectados por la caza en SACHA y CAPH. El Sajino, el Venado Rojo y el Añuje no mostraron evidencias de impacto alguno, y parece que estas especies presentan características reproductivas y de comportamiento que les otorga la capacidad de soportar los niveles de caza existente. Hubo insuficientes datos para análizar el impacto de las demás especies: la Maquisapa, el Machin Blanco, la Sachavaca y la Huangana. Estas especies se discuten lineas abajo.
La abundancia relativa de huellas de los dos felinos más comunes, indican que los dos son afectados por los altos niveles de caza alrededor de SACHA y CAPH, aunque el Tigrillo aparentemente es más resistente.
Maquisapa (Ateles paniscus)
La Maquisapa esta evidentemente extinta localmente, alrededor de los 4 albergues situados cerca los asentamientos humanos de los rios Tambopata y Madre de Dios, donde hay o ha habido una presión de caza (Figura 3.25). Información proporcionada por los habitantes locales indica que esta especie ha desaparecido de gran parte de las áreas cercanas a los rios navegables y esto mucho antes de la aparición del turismo en la zona. Sin embargo recolectabamos información sobre tres avistamientos no confirmados de esta especie en la parte baja del río Tambopata que indica que esta especie tiene la potencialidad de recolonizar el área. El único albergue donde se observó esta especie fue en TRC, donde aún existe una población saludable, aunque hay evidencia que el tránsito humano puede estar afectando negativamente la distribución local. Esta hipótesis de todas maneras requiere de más investigación para estar completamente seguro. En Manú donde la especie también es común Yoon (1997) encontró que esta no muestra señales de ser impactada por el tránsito turístico, aunque los lugares de estudio que Yoon utilizó tuvieron intensidades de tránsito más bajas que aquellos que actualmente experimenta el TRC.
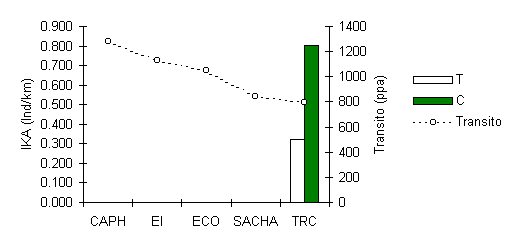
Figura 3.25. IKA de la Maquisapa (Ateles paniscus).
Sachavaca (Tapirus terrestris)
En el caso de esta especie hubo insuficientes datos, en términos de encuentros visuales, para realizar análices estadísticos. Aunque los datos de abundancia relativa de huellas revelaron que era más abundante en TRC y EI (Figura 3.21), los dos albergues con la menor presión de caza. Un análisis de correlación de Pearson indicó que la abundancia de huellas era correlacionada negativamente con la presión de caza (Pearson = -0.75), aunque no significativamente. No hay evidencia que esta especie este afectada en lo absoluto por el tránsito turístico.
Huangana (Tayassu pecari)
Esta especie también fue observada insuficientemente para determinar la importancia de las variables de impacto bajo estudio, aunque los datos reflejan la opinión de la poblacion local, que señala que la especie se aleja de asentamientos humanos permanentes y es más abundante en áreas aisladas como TRC, opinión también revelada en otros estudios de esta especie (Fragoso 1991, Kirkby et al. 1998, Kirkby 1999). Presumimos que esto es resultado de la presión de caza cerca a comunidades. Esta especie además es suceptible a enfermedades y se presume que la abundancia regiónal fluctua fuertemente a través de las decadas debido a esto, ya que su comportamiento de vivir en grandes grupos sociales incrementa la posibilidad de transmisión. Enfermedades introducidas por el hombre, a través de los animales domésticos, son implicadas en este fenómeno. Esto también podria explicar la ausencia de la especie en áreas donde hay asentamientos humanos. Después de haber sido eliminados de un área, debido a la caza y/o enfermedades, la recolonización natural es extremedamente lenta.
No encontramos evidencia para sugerir que el turismo tiene un impacto sobre esta especie, y es poco probable que sea encontrada en el futuro. El territorio de un grupo de esta especie es enorme, a veces del orden de cientos de kilómetros cuadrados, entonces es poco probable que esta especie pueda percibir adecuadamente (en términos espaciales) áreas de alto tránsito turístico y activamente evitarlas, ya que sus visitas a áreas utilizadas por turistas son muy infrecuentes y usualmente no duran más de unas horas. Sin embargo, la experiencia de uno de los autores (CK) con esta especie, sugiere que los grupos encontrados en TRC eran relativamente más tímidos y más propensos a huir a distancias más grandes que en otras zonas de Madre de Dios donde la presencia humana es nula. Es posible presumir que el olor del ser humano alrededor de las trochas turísticas en el TRC genera un estado de inquietud en esta especie y la hace propensa a la huída.
Como recurso turístico inusual y excitante (ya que viven en manadas grandes y generan bastante ruido) esta especie debería recibir más protección a nivel local. Para esto la caza cerca albergues tendría que ser limitada a una intensidad que las poblaciones actuales de esta especie pueden soportar.
Los Efectos del Tránsito Turístico
Los resultados de las pruebas-t para muestras pares, para investigar si hubo diferencia significativa entre T y C en términos del %IKA de 13 especies (Tabla 3.19) - incluyendo el machin Negro y el Coto - revelaron que dos especies, el Venado Rojo y el Machin Negro, eran significativamente más abundante a lo largo de C, y que dos especies, el Oso Hormiguero y la Ardilla Ploma, eran significativamente más abundate a lo largo de T (Tabla 3.19).
Tabla 3.19. Resultados de las pruebas-t para muestras pares (1 cola) comparando la abundancia de 13 especies entre T y C [El análisis utilizó datos de %IKA]. ca = especies principalmente impactadas por la caza. * p < 0.05, ** p < 0.01, + casí significativa.
| Especies | Promedio IKA T |
Promedio IKA C |
p |
. |
| Oso Hormiguero | 0.016 |
0.006 |
0.03 |
*T |
| Pichico | 0.787 |
0.580 |
0.41 |
. |
| Fraile | 1.399 |
1.053 |
0.30 |
. |
| Tocón | 0.046 |
0.071 |
0.23 |
. |
| Coto (ca) | 0.163 |
0.073 |
0.06 |
+T |
| Machin negro (ca) | 0.329 |
0.465 |
0.05 |
*C |
| Achuñi | 0.047 |
0.074 |
0.47 |
. |
| Manco | 0.031 |
0.035 |
0.43 |
. |
| Sajino | 0.066 |
0.092 |
0.20 |
. |
| Venado Rojo | 0.005 |
0.026 |
0.01 |
*C |
| Ardilla Ploma | 0.096 |
0.069 |
0.03 |
*T |
| Ardilla Roja | 0.493 |
0.430 |
0.38 |
. |
| Añuje | 0.138 |
0.134 |
0.36 |
. |
Los análices de correlación de Pearson entre el IKA T y el tránsito turístico, de las mismas 13 especies anteriormente mencionados, revelaron 3 resultados significativos (Tabla 3.20). Dos especies eran correlacionadas positiva y significativamente (el Pichico y la Ardilla Roja) y una correlacionada negativamente (el Venado Rojo), la misma que también fue identificado en los análices de pruebas-t para muestras pares (Tabla 3.19). La diferencia entre las desviaciones estándares (DS) entre T y C para el Pichico y la Ardilla Roja también era grande y muestra que la abundancia en C era más estable entre albergues que la en T, una característica esperada en especies afectadas por el turismo.
Tabla 3.20. Correlación de Pearson, IKA T vs. tránsito, y la diferencia porcentual entre T y C en términos de la desviación estándard del promedio (DS). * p < 0.05, (T) = DS T > DS C.
| Especies | Pearson |
. | % Diferencia DS (T vs. C) |
| Oso Hormiguero | - 0.539 |
. | . |
| Pichico | + 0.916 |
* | 272 % (T) |
| Fraile | - 0.854 |
* | 251 % (T) |
| Tocón | + 0.384 |
. | . |
| Coto (ca) | - 0.226 |
. | . |
| Machin negro (ca) | - 0.532 |
. | . |
| Achuñi | - 0.477 |
. | . |
| Manco | - 0.460 |
. | . |
| Sajino | - 0.044 |
. | . |
| Venado Rojo | - 0.883 |
* | . |
| Ardilla Ploma | - 0.498 |
. | . |
| Ardilla Roja | + 0.946 |
* | 167% (T) |
| Añuje | + 0.517 |
191% (T) |
Oso Hormiguero (Tamandua tetradactyla)
El análisis de pruebas-t y la Figura 3.26 muestran claramente que esta especie es relativamente más común en los T que en los C. En CAPH la especie no fue registrada durante ningún censo, aunque se observó durante encuentros fortuitos en áreas no bajo estudio, aunque, nuevamente cerca a las trochas turísticas. Un análisis de correlación de Pearson entre IKA en T y la intensidad del tránsito de un albergue a otro no reveló ningún patrón significativo, aunque debido a la muestra de sólo 14 encuentros es posible que haya insuficientes datos para estar seguros sobre la relación entre estas variables, y sugerimos que se haga más estudios de esta especie para resolver esto.
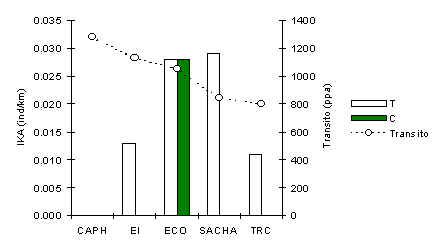
Figura 3.26. IKA del Oso Hormiguero.
Machin Negro (Cebus apella)
El patrón de abundanica de este primate muestra, claramente, su suceptilibilidad a la caza como fue mencionado anteriormente (Figura 3.27). Sin embargo, habia un patrón secundario significativo, en el que su abundancia era consistentemente mayor en los C (Tabla 3.19), particularmente en aquellos albergues donde existió niveles históricos de caza, por estar cerca asentamientos humanos permanentes. Entonces, aunque esta especie esta impactada por la caza, en estas zonas la presencia de turistas en su hábitat también es un factor negativo que reduce aún más su población. Esto puede ser una manifestación de que la especie tiene un temor natural al hombre, en aquellas poblaciones locales donde han aprendido a temerles en base a encuentros con cazadores. Podemos especular que en estos albergues la especie no puede diferenciar entre un turista y un cazador y tiende a huir de las trochas turísticas como consecuencia. Un análisis de correlación de Pearson entre el IKA en T y la intensidad de tránsito turístico en los 3 albergues menos afectados por la caza (TRC, EI, ECO) reveló que habia una relación negativa, aunque no significativa (Pearson = -0.95, p < 0.10).
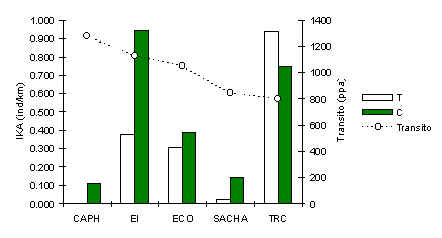
Figura 3.27. IKA del Machin Negro.
Coto (Alouatta seniculus)
El patrón de abundancia de esta especie de primate, tal como el machin Negro, muestra que ha sido severamente impactada por la caza aldedor de SACHA y CAPH (Figura 3.28). Sin embargo, es muy interesante que en la mayoría de los albergues, incluyendo aquellos mencionados, la abundancia es mayor a lo largo de los T, particularmente en EI y TRC donde la presión de caza es nula, indicando que el turismo es una variable de impacto secundario pero importante de todas maneras ya que incrementa la abundancia a nivel local.
Una correlación de Pearson entre el IKA en T y la intensidad de tránsito turístico en los 3 albergues menos afectados por la caza (TRC, EI, ECO) reveló que no habia relación entre estos factores. Solamente podemos especular, en esta oportunidad, sobre las razones subyacentes de la diferencia entre T y C. Podria ser una reducción en la presión de depredación, aunque no hay suficientes datos sobre las poblaciones de sus principales depredadores como el Aguila Harpia (Harpia harpyja), y/o una reducción en los parámetros de competencia inter-específica para recursos. Se podria concluir que la especie ha logrado un grado de habituación hacia la presencia de turistas en su hábitat, que a su vez podria haber incrementado su población cerca a las trochas y/o incrementando las posibilidades de observarlas, ya que son normalmente de comportamiento tímido y secreto.
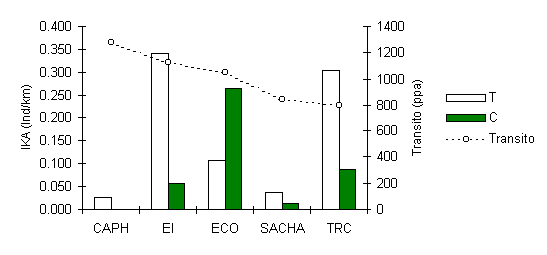
Figura 3.28. IKA del Coto.
Venado Rojo (Mazama americana)
Los análices revelaron que esta especie es la más impactada por el tránsito turístico. En todos los albergues su abundancia fue mayor en los C (Figura 3.29), sin embargo la relación era menos fuerte en TRC. Una correlación de Pearson entre el IKA en T y la intensidad de tránsito turístico en cada albergue, reveló una relación negativa y significativa (Pearson = –0.88, p < 0.05) que respalda el resultado anterior.
Además de ser comúnmente cazada en Madre de Dios, parece que las intensidades de caza encontradas en esta oportunidad no son suficientes para afectar su abundancia. Los resultados muestran que el Venado Rojo huye permanentemente de las zonas cercanas a las trochas turísticas, y las mayores densidades en áreas más alejadas se mantienen en niveles mayores que lo normal, particularmente alrededor de aquellos albergues que tienen historia de caza, ya que hay evidencia que los depredadores grandes como el Jaguar son menos comunes en aquellos albergues.
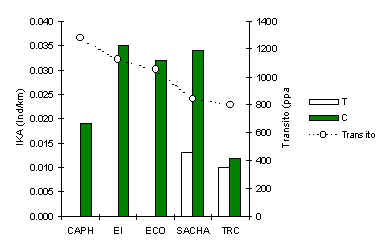
Figura 3.29. IKA del Venado Rojo.
Ardilla Ploma (Sciurus ignitus)
En cuatro de los cinco albergues esta especie fue significativamente más abundante a lo largo de los T (Figura 3.30). Aunque, un análisis de correlación de Pearson entre IKA de T y el tránsito turístico en los cinco albergues no reveló ninguna relación (Tabla 3.20). La fluctuación en abundancia promedio de un albergue a otro es probablemente el resultado de un variable del hábitat, depredador o competidor, del cual no tenemos mayor información, aunque es claro que el efecto del tránsito es fuerte y el patrón en TRC indica que la especie es probablemente sensitiva a niveles de tránsito menores a las investigadas en esta oportunidad. También, la variabilidad entre los IKA en C era menor que en T (Desviación Estándard; T = 0.06, C = 0.04).
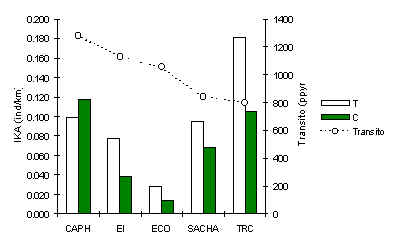
Figura 3.30. IKA de la Ardilla Ploma.
Pichico (Saguinus fuscicollis)
El patrón de abundancia de este primate (Figura 3.31), es muy similar a aquello observado en la Ardilla Roja, mamífero de tamaño similar. La correlación Pearson entre IKA de T y tránsito reveló una relación positiva y significativa (Tabla 3.20). El mismo tipo de análisis, pero utilizando los datos de IKA de C, también reveló una relación positiva aunque esta vez no era significativa. Estos resultados nos lleva a concluir que esta especie esta siendo afectada por niveles de tránsito progresivamente y al incrementar la intensidad del tránsito el efecto tiende a extender a mayores distancias de las trochas turísticas. Además, la desviación estandar entre los IKA en T de un albergue a otro era mucho mayor que la de C (Desviación Estandard; T = 0.66, C = 0.24).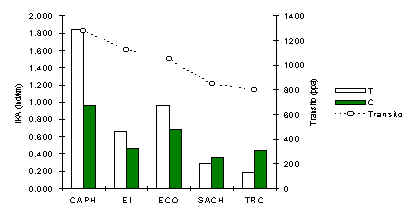
Figura 3.31. IKA del Pichico.
Ardilla Roja (Sciurus spadiceus)
El análisis de correlación Pearson entre el IKA de T y tránsito (Pearson = +0.95, p < 0.01), encontraron relaciones positivas y significativas que nos llevan a concluir que esta especie también es muy sensitiva a la magnitud de tránsito bajo estudio. Además, tal como el Pichico la diferencia en la variación estandard entre T y C fue alta y un síntoma de impacto turístico. Como fue mencionado anteriormente el patrón de abundancia de esta especie de ardilla (Figura 3.32) es muy similar al del Pichico. Un análisis de regresión entre los IKA de estas especies, mostró una relación positiva y muy significativa (Z = 66.517, p < 0.001). Estas especies podrian tener una fuerte relación inter-específica. Los datos recolectados sobre comportamiento mostraron que hubo varias ocasiones, en diferentes albergues, cuando estas especies interaccionaron y formaron asociaciones por tiempos cortos durante los cuales las especies parecian estar forrageando juntas, aunque no hubo suficiente evidencia para encontrar una relación significativa entre sus abundancias generales. Además, hay poca evidencia en este u otros estudios que estas especies comparten aspectos de su dieta cotidiana o esfuerzo antidepredador. Pero presentamos evidencia que las dos especies son generalmente más abundantes en bosques donde el recurso fruta es relativamente reducido.
Concluimos entonces que estas especies son afectadas de manera similar por un variable externa que probablemente sea el turismo, de manera directa a través de la habituación al turista, y/o indirectamente a través de la falta de los depredadores como; halcones, tigrillos, boas, etc. - hipótesis que es consistente con el tamaño de estas especies y el hábitat que utilizan, es decir que forragean frecuentemente en el sotobosque y subdosel.
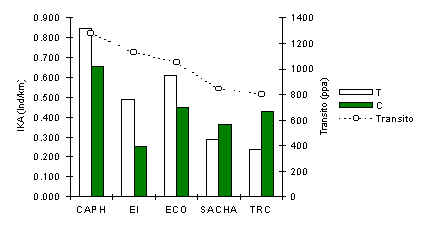
Figura 3.32. IKA de la Ardilla Roja.
Fraile (Saimiri boliviensis)
El patrón de abundancia de esta especie de mono (Figura 3.33) muestra variaciones considerables entre T y C, y entre albegues en general. La variabilidad en abundancia entre albergues fue generalmente más estable en C (Desviación Estandard; T = 1.46, C = 0.58), y un análisis de correlación Pearson entre IKA de T y tránsito turístico reveló una relación nagativa significativa (Tabla 3.20), síntomas que indican que el tránsito de turistas es posiblemente un factor negativo que podria afectar la abundancia de esta especie. Sin embargo, el tamaño de los territorios de los grupos de esta especie es muy grande, que junto con la metodología empleada genera una situación donde es difícil identificar una diferencia significativa entre la abundancia de T y C, si realmente existia, porque la distancia promedio de separación entre estas áreas de muestreo en cada albergue esta dentro de la distancia que un grupo normal cubriría en una temporada. Esto significa que los grupos residentes en cada albergue, aunque posiblemente tímidos a la presencia de turistas o afectados indirectamente por ellos, todavía tienden a usar áreas de bosque cerca las trochas turísticas para forragear y transitar de un lado de su territorio a otro. Estas ideas no son conclusivas, pero al introducir la variable hábitat, en términos del recurso fruta, hay una relación significativa. Un análisis de regresión entre la abundancia promedia de fruta en cada albergue y la abundancia promedia de esta especie mostró una relación positiva y significativa (F = 19.086, p = 0.02). Además los bosques alrededor de SACHA y TRC son estructuralmente más maduros con menos perturbación del dosel (Lloyd y Palomino, capitulo 5.0), factores que benefician las poblaciones de insectos, uno de los constituyentes importantes del dieta de este primate. Aunque no podemos descartar por completo el efecto del tránsito, parece que el hábitat juega un rol importante en la distribución del Fraile.
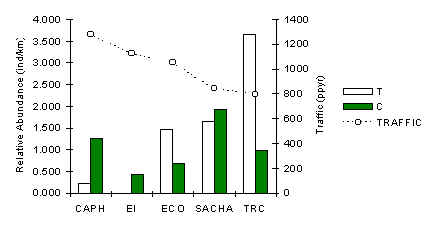
Figura 3.33. IKA del Fraile.
Achuñi (Nasua nasua)
Esta especie esta presente en todos los albergues (Figura 3.34), aunque los análices arriba mencionados no encontraron una relación significativa con el tránsito turístico. La característica más importante es la poca abundancia del Achuñi en CAPH y EI, los dos albegues más antiguos de la zona. Una relación negativa y significativa se mostro al comparar la abundancia promedia en cada albergue con el número de años que los albergues habian estado operando (p = 0.02) (Figura 3.35). Entonces, en una escala macro esta especie parece ser impactada por el turismo, en términos de la presencia de turistas en su hábitat durante un tiempo extendido, aunque no es claro si los niveles de tránsito actual estan afectando esta espcie en los albegues más jovenes. También se podria especular que la poca abundancia encontrada en CAPH y EI es un factor del recurso fruta, tal como el caso del Fraile antes mencionado, aunque los datos de TRC son más reducidos que lo esperado si fuese asi, aunque esto podria ser por la presencia de mayores depredadores y mayor competencia para el recurso fruta. En esta oportunidad no podemos distinguir adecuadamente entre estos factores y la especie requiere más estudio.
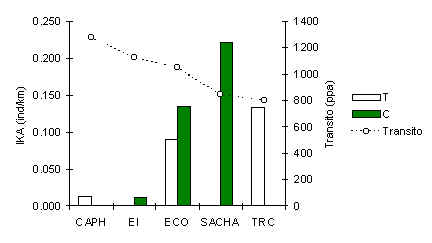
Figura 3.34. IKA del Achuñi.
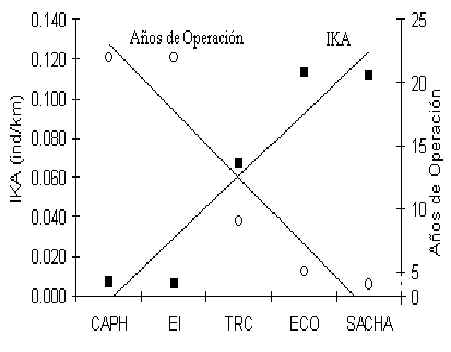
Figura 3.35. La relación entre el IKA del Achuñi en cada albergue y los años desde que albergue empezo a operar.
Similaridades entre Albergues
De las especies discutidas arriba, 5 de ellas mostraban señales de haber sido afectadas significativamente por el tránsito turístico: Venado Rojo, Pichico, Machin Negro, Ardilla Ploma, y la Ardilla Roja. Se utilizó un análisis de conjuntos jerárquicos para combinar los datos de estas especies para confirmar si los patrones entre T y C eran consistentes de un albergue a otro. Para realizar esto se tuvo que estandarizar los datos de todas estas especies, esto se hizo en términos de porcentage de IKA, es decir T% vs. C%. El resultado (Figura 3.36) muestra claramente una separación entre T y C indicando que estas especies, como grupo, son consistentemente afectadas por esta variable y son los mejores candidatos como Especies Indicadoras de Impacto Turístico (EIIT). Un análisis similar pero con datos agrupados por albergue (Figura 3.37), mostró que los dos albergues con los niveles de tránsito más altos eran los más similares en terminos de la abundancia de estas cinco especies.
EIC ------------------+
CAPHC ------------------+----+
+--+
SACHAC ---------------------+-+ +
ECOC ---------------------+ +--+
TRCC --------------------------+ +
+-----------------
EIT ------------------+ +
CAPHT ------------------+----+ +
+--+ +
SACHAT ---------------------+-+ + +
ECOT ---------------------+ +--+
TRCT --------------------------+
Figura 3.36. Dendrograma mostrando la relación entre T y C de todos los albergues, basado en el % del IKA de 5 especies (Venado Rojo, Pichico, Machin Negro, Ardilla Ploma, y la Ardilla Roja).
EI ---------------+
CAPH ---------------+---+
+--+
SACHA -----------------+-+ +
ECO -----------------+ +---------
TRC ----------------------+
Figura 3.37. Dendrograma mostrando la relación entre albergues, basado en los IKA agrupados de las mismas 5 especies.
Efectos a Nivel de Comunidad
Primates
TRC tiene un máximo de especies de primate para su localidad, debido a su estado de aislamiento (Figura 3.43). El Machin Blanco, aunque no observado durante los censos, fue avistado durante encuentros fortuitos de este albergue. En general, los albergues situados cerca a los asentamientos humanos permanentes, tuvieron una biomasa de primate reducida en comparación con TRC (Figura 3.38). Este patrón fue más evidente en SACHA y CAPH donde el Machin Negro y el Coto están siendo exterminados. Un análisis de pruebas-t para muestras pares (1 cola) reveló que hubo una diferencia casí significativa entre T y C en términos de %IKB (Tabla 3.22). Además, el promedio del IKB de cada albergue no era fuertemente relacionado al recurso fruta disponible. Esto nos lleva a concluir que la caza sobre todo esta afectando negativamente a esta comunidad, y provee respaldo adicional a las conclusiones introducidas originalmente por Freese et al. (1982) que el factor principal afectando la biomasa de los primates Amazónicos es la caza furtiva.
La estructura de la comunidad en los albergues fuertemente impactados por la caza (SACHA y CAPH) es muy similar, y a su vez son dominados por las especies de primate más pequeños, como el Pichico y el Fraile, especies que son más eficientes reproductivamente cuando escapan de los efectos de la depredación y la competencia interespecífica, a su vez resultado del turismo. Además, pueden soportar una menor disponibilidad de fruta en su hábitat debido a sus dietas que son relativamente diversas e incluyen insectos. Donde la presión de caza es baja o completamente ausente, las comunidades de primates son dominadas por las especies grandes, particularmente el Coto, el Machin Negro, y en el caso TRC la Maquisapa.
Es interesante que el albergue más similar a TRC es EI, que sugiere que la falta de caza alrededor de EI ha sido de beneficio en la mantención de la estabilidad de la comunidad en esta zona, aunque la biomasa total de primates ha sido afectada debido a la falta del recurso fruta y la inmigración reducida debido a la presión de caza fuera de las áreas visitadas por el turista.
El Tocón siempre parece ser un miembro menor de esta comunidad, y aprentemente favorece los bosques más perturbados y donde el bambú es común. Esta conclusión es consistente con estudios más específicos de las preferencias de hábitat de esta especie en Tambopata (Warner 1999).
Tabla 3.21. Datos referentes al peso corporal adulto (kg) de 18 especies, incluyendo el valor de ¾ del mismo en el caso de especies sociales donde juveniles fueron comúnmente encontrados. * el peso utilizado en los cálculos de IKB. 1 Clutton-Brock et al. 1977, 2 Emmons 1984, 3 Robinson et al. 1986, 4 Ayres et al. 1991, 5 Mittermeier 1991, 6 Peres 1993, 7 Johns et al. 1987.
| Comunidad | Peso corporal del adulto (kg) | Promedio (kg) |
¾ Promedio(kg) |
| Primates | . | . | . |
| Pichico | 0.402, 0.363, 0.396 | 0.38 |
0.29* |
| Fraile | 0.671, 1.002, 0.803, 0.906 | 0.84 |
0.63* |
| Tocón | 0.681, 0.802, 1.173, 0.704 | 0.84 |
0.63* |
| Coto | 7.002, 6.193, 7.005, 6.506 | 6.67 |
5.00* |
| Machin Negro | 2.501, 3.002, 3.453, 3.705, 2.206 | 2.97 |
2.23* |
| Maquisapa | 7.83, 8.506, 8.107 | 8.13 |
6.10* |
| Ungulados | . | . | . |
| Sachavaca | 1103, 1775 | 143.65* |
. |
| Huangana | 18.003, 28.565 | 23.28 |
17.46* |
| Sajino | 17.523, 12.005 | 14.76 |
11.07* |
| Venado Rojo | 26.103, 23.54, 15.005 | 21.53* |
. |
| Carnívoros | . | . | . |
| Manco | 3.983 | 3.98* |
. |
| Achuñi | 3.883, 3.405, 3.004 | 3.43 |
2.57* |
| Perro de Monte | 7.002 | 7.00* |
. |
| Tigrillo | 10.462 | 10.46* |
. |
| Roedores | . | . | . |
| Ardilla Ploma | 0.202, 0.383 | 0.29 |
0.22* |
| Ardilla Roja | 0.602 | 0.60 |
0.45* |
| Myoprocta sp. | 1.53 | 1.5* |
|
| Añuje | 3.603, 4.205 | 3.90 |
2.93* |
Ungulados
Nuevamente, el TRC era el único albergue en tener un complemento total de especies de ungulados para su localidad (Figura 3.42). La Huangana y la Sachavaca además eran abundantes en relación a otros albergues. A consequencia este albergue tuvo la mayor IKB (Figura 3.39). Al otro extremo, los dos albergues con el mayor tránsito turístico, además han estado establecidos por más tiempo que cualquier otro albergue (CAPH y EI), el Sajino y el Venado Rojo eran los únicos que se podria observar, aunque la Sachavaca estaba presente en números muy reducidos pero sólo fue registrado a través de sus huellas. Un análisis de pruebas-t para muestras pares (1 cola) reveló que hubo significativamente más biomasa de ungulado en los C que en los T (Tabla 3.22) indicando que el turismo podria ser responsible de la variación en biomasa de esta especie.
Roedores
Esta comunidad fue dominada en cada albergue por el Añuje (Figura 3.44). También era evidente que el Myoprocta sp. siempre era un constituente menor de la comunidad pero también fue más común en los albergues más perturnados en términos de hábitat y tránsito turístico. La IKB de los roedores llego a su máximo en CAPH (Figura 3.40). El patrón de IKB era casí lo opuesto al encontrado por los primates, y se ve claramente que esta comunidad recibe una ventaja en aquellos albergues bajo una alta presión de caza (SACHA y CAPH), que sugiere que estan beneficiando de una reducción en la abundancia de depredadores y/o una reducción en la competencia inter-específica. Un análisis de Pruebas-t para muestras pares (1 cola) reveló que no hubo diferencia significativa entre T y C en términos de IKB (Tabla 3.22), indicando que el turismo no tiende a tener un efecto sobre esta comunidad en términos generales.
Carnívoros
TRC tuvo la mayor diversidad de especies carnívoras, que incluyo el Perro de Monte Plomo (A. microtis), que generalmente es muy raro en la mayoría de su rango geográfico. Dos especies eran relativamente comunes en todos los albergues, el Achuñi o el Manco (Figura 3.45), especies reconocidas por su dieta omnivora que incluye a la vez bastante fruta. En SACHA el valor de la biomasa era mayor que otros albergues (Figura 3.41) gracias a la abundancia de estas especies, aunque el albergue con la mayor biomasa de carnívoros con dietas completamente carnívoros, tal como los felinos y los perros de monte, era TRC. Un análisis de correlación de Spearman entre IKB de T y tránsito turístico reveló una correlación negativa y significativa (rs = -0.9, p = 0.05), un resultado similar con respecto a la abundancia de huellas del Tigrillo (Figura 3.19). Aunque, el análisis de pruebas-t para muetras pares no mostro una relación significativa entre T y C. Al combinar los datos de T y C se vio con que SACHA y el TRC se destacaron en tener significativamente más biomasa que los otros albergues. Estas dos albergues son relativamente nuevo y tienen los niveles de tránsito más bajos. Hay, entonces pocas señas que este grupo taxonómico esta afectado por la magnitud del tránsito, sin embargo, la biomasa del Achuñi y el Manco esta muy relacionada con el recurso fruta, entonces el hábitat podria estar teniendo un efecto más importante (Tabla 3.22). Esta comunidad requiere de mayores estudios para incrementar la confiabilidad de las estimaciones de abundacia.
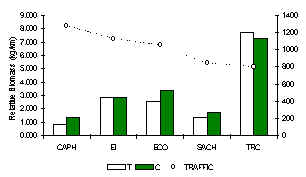
Figura 3.38. IKB de los primates.
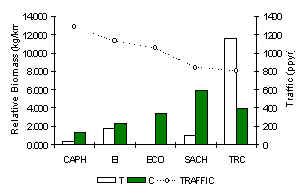
Figura 3.39. IKB de los ungulados.
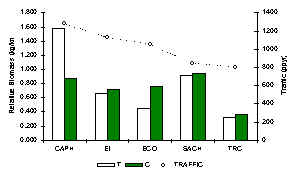
Figura 3.40. IKB de los roedores.
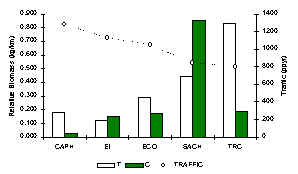
Figura 3.41. IKB de los carnívoros.
Tabla 3.22. Resultados de pruebas-t para muestras pares (1 cola) mostrando diferencias entre T y C en términos de comunidad taxonómica, * p < 0.05, + casí significativo.
| Albergue | Primates |
Ungulados |
Carnívoros |
Roedores |
Mamíferos en General |
| No. de Species | 6 |
4 |
4 |
4 |
18 |
| p | 0.07 +C |
0.05*C |
0.46 |
0.40 |
0.16 |
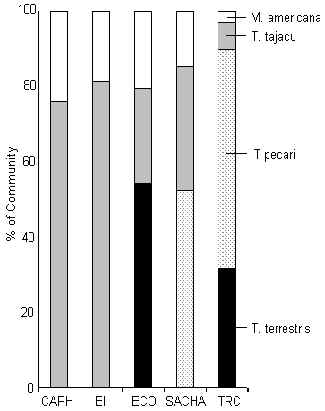
Figura 3.42. Ungulados (%IKB).
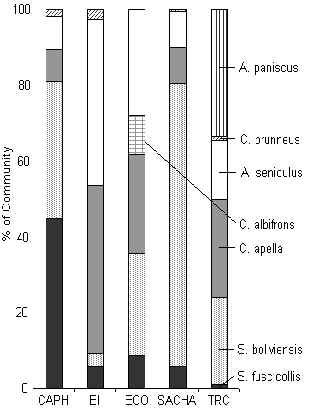
Figura 3.43. Primates (%IKB).
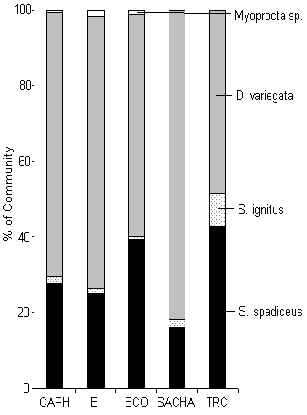
Figura 3.44. Roedores (%IKB).
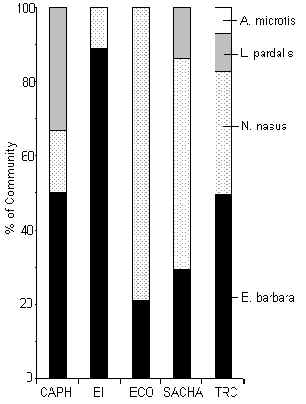
Figura 3.45. Carnívoros (%IKB).
Mamíferos en General
La biomasa de mamíferos en general, en terminos de los IKB de las 18 especies discutidas anteriormente, era mayor en TRC (Figura 3.46), y concluimos que este albergue por lo tanto tiene una comunidad de mamíferos relativamente saludable y estable. Los demas albergues tuvieron biomasas deprimidas en comparación probablemente resultado de una mayor presión de caza historica o una reducción en el recurso fruta alrededor de estos. El resultado de la prueba-t para muestras pares que comparó T y C en terminos de %IKB (Tabla 3.22) mostró que el tránsito turístico no es responsable de la variación en biomasa de mamíferos de un albergue a otro.
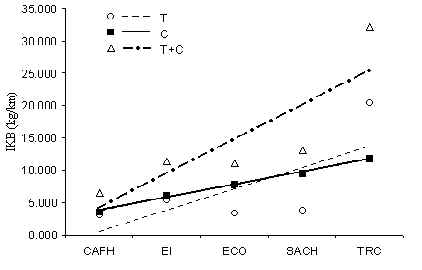
Figura 3.46. IKB de 18 especies de mamífero.