| STOMATAL GUARD CELL MEASUREMENTS USING LEAF PRINTS |
An addendum to this page offering some hi tech options, as well as links, is
accessed by
CLICKING HERE.
Many of us are aware of the value of knowing the ploidy level of the cymbidiums
with which we are working. It often happens that one looks at a cymbidium and
would like to get some idea of its ploidy level, either because it is to be
used in breeding, to understand more about it, or because it is different to
what was expected. As a chromosome count is usually too difficult to perform or
have done, I am presenting a technique here which should be able to help
towards clarifying the ploidy level question.
It is awe-inspiring to consider the number of people and the huge amount
of work done, representing the inherited knowledge of the ages, which has
allowed us to reach our present state of knowledge. With regard to the stomatal
guard cell measurements and their ploidy implications alone, one must think of
Robert Hooke and his microscope (1665), early plant microanatomists and
interpreters like Malpighi (1675) and Robert Brown (1831), the Cell Theory of
Schleiden and Schwann (1838). In 1848, Hofmeister observed dark staining
elements in cell nuclei, which Strasburger described and illustrated in 1875-7
coining the word "chromosome" for them. In 1882, Strasburger was the
one who stated that the number of pairs of elements (chromosomes) is generally
the same for all plants in a species, but may differ from species to species.
Gerlach's establishment of stain technique (1858) was an important technical
milestone. The theory of natural selection of Darwin and Wallace (1858) and
Gregor Mendel's work on genetics (1866) number amongst the greatest
intellectual achievements of the 19th century.
The 20th century brought Winkler's introduction of the term "genome"
(1916), Winge's proposed theory of the polyploid origin of plant species (1917)
and Belling's Aceto-carmine squash technique for chromosome counting (1921).
The application of colchicine to cause the ploidy level to double was
discovered by Nebel and Ruttle (1937) following the first isolation,
characterisation and naming of colchicine by Dustin in 1932. GW Burns's work
showed that plants of one species, naturally having varying levels of ploidy
could be reliably separated into ploidal groups by comparing the areas of
lower leaf epidermal cells from different plants (1942). I do not know who
extrapolated this idea to stomatal guard cell measurement, but in 1954 Don
Wimber submitted the thesis "Stomatal size in relationship to ploidy level
in the cultivated Cymbidium" to the Claremont Graduate School for his
Masters degree. I am unable to find any published material emanating from this
thesis.
(P.S. By courtesy of Mark Gibbel, I have received a copy of this now 50 year-old thesis. It appears that the aforementioned Winkler was the first person to show a relationship between stomatal cell size and ploidy level. In the Summary at the end of this thesis, Wimber says:
"If there are significant differences between stomatal measurements of the parental plants of different ploidy levels, progeny from such matings, as diploid X triploid or triploid X tetraploid, possibly may be classified according to their chromosome number through examination of the stomata.".
Thanks, Mark.)
Don Wimber was my introduction to guard cell measurements, through his 1967
papers in the Cymbidium Society News (vol. 22(5), p. 112) and in the Proceedings
of the 5th World Orchid Conference (p. 27, together with Ann Van Cott). Merv
Dunn of Australia also contributed notes in two articles published in the
Orchid Advocate of 1978 (vol. 4 (1), p. 22, and vol.4 (3), p. 84).
The theory is essentially simple. The volume of a cell in a particular
plant is directly proportional to the amount of DNA present, so doubling of
the amount of DNA, which occurs when a plant is converted from a diploid to a
tetraploid, causes the cell to have double the volume. Since it is the volume
that doubles, the increase in the size of the cell in any one dimension is
approximately 1.25. (1.25 x 1.25 x 1.25 = ca. 2). By comparing the lengths of
the guard cells of different, closely related plants, one can usually take an
educated guess at the ploidy level.
In the genus Cymbidium, there is, throughout, a reasonably stable diploid stomatal guard cell
measurement in the region of 27 - 28 microns long. From this, one can
extrapolate a triploid size of about 31 -32 microns, a tetraploid size of 34 -
35 microns and a hexaploid size of 39 - 43 microns. These are approximate, and
there are probably very many exceptions. The technique has particular value
when one is trying to make some sense of an unusual plant in an otherwise
fairly even batch. It also has other uses, such as finding converted plants
amongst a batch of colchicine treated plantlets.
Other genera may be less accommodating. It has been found that the higher a
plant grows above the ground, the smaller the guard cells tend to be. So when
one is dealing with a genus like Zygopetalum, where both terrestrial and epiphytic plants are used in breeding, adding to
that the complication of agamospermy and probable micro-species formation,
trying to make sense of guard cell measurements in Zygopetalum hybrids is a totally futile pursuit.
Technically, there is nothing particularly difficult here. One requires a
microscope with a 40x objective and a 10x eyepiece. An eyepiece micrometer is a
microscope eyepiece fitted with a disc of glass upon which a small measuring
scale has been etched, known as a graticule. The graticule fits inside the
eyepiece on top of the field restrictor. In a fancy setup, one uses a focusing
eyepiece, where the distance between the front lens and the graticule can be
adjusted for focusing, but in practice, unless one's vision is abnormal, there
is normally enough thread on the front lens component to afford adequate
focusing with a standard eyepiece. With this setup, one can simultaneously view
the graticule and the sample, and with correct alignment of the sample, measure
the length of, in our case, the stomatal guard cell.
One may simply compare the average measurements of the guard cells of
various plants, one with the other, without defining the exact measurement.
Another approach would be to determine the actual measurement by calibrating
the eyepiece micrometer against another graticule of known length placed on the
microscope stage, or by using a specially made slide with a graduated scale of
known length engraved on it, known as a stage micrometer. By this means, one
can then calculate what length one unit of the eyepiece micrometer actually
represents.
Originally, I took thin slices of leaf-surface tissue by means of a sharp
razor blade, wet mounted these on a slide with coverslip and proceeded with the
measurement procedure. Even if the centre if the leaf slice was a bit on the
thick side, there was normally sufficient thin material towards the edges of
the preparation to find guard cells for measurement. I have always had concerns
when performing this procedure. It is necessary to remove pieces of leaf from a
number of plants. As far as I am concerned, no matter how carefully one
proceeds, there is always a chance of sap contamination and with that, virus
transmission.
In 2001, I came across the work of R.J. Ferry, Sr, Leafprints and Statistical Analyses: Findings and Implications published in the Orchid Digest, Jan.-Feb.-Mar. 2000 (vol. 64 (1), pp. 21-6). In this
article, Bob Ferry described his work towards his doctoral thesis, which
involved statistically analysing leaf epidermal cell area sizes in Stanhopea species and arriving at some good scientific conclusions. What particularly
interested me was his technique of making leafprints, a procedure based on the
relatively obscure 1965 works of E.M. Stoddard (Identifying plants by leaf epidermal characters. - Connecticut Agric. Exp. Sta. Circ. 227, p. 3) and R.J. Petroski (Clonal identification by means of epidermal characters. - Orchidata vol. 5 p. 191). The procedure used here requires the application of a
thin smear of a rapidly drying solution of some clear plastic substance to the
leaf surface. The smear is allowed to dry and is transfered to a piece of clear
pressure-sensitive tape, the tape then being attached directly to a slide for
examination. Although a clear nail polish was recommended as the print medium
in the earlier papers, Ferry recommended a xylol/styrofoam mixture.
Xylol (=xylene or dimethyl benzene) is a solvent used by microscopists for
cleaning lenses. (Some components of microscope lenses are mounted in media
which are soluble in alcohol, but unaffected by xylol.). Most microscope owners
have or should have xylol to hand. For the styrofoam component, a clean piece
of styrofoam or styrofoam granules will work. To make up the reagent, one
places a small quantity of the xylol in a small glass bottle which has a metal or bakelite lid (xylol migrates through polystyrene
in quick succession), and then one adds small pieces of styrofoam, piece by
piece, until a solution with the consistency of thin honey is produced. At this
point, there are often numerous, small air-bubbles in the solution, so the
solution should stand for an hour for these bubbles to rise to the surface and
dissipate.
Using an earbud or matchstick, apply a small quantity of the
xylol/styrofoam to the underside of a mature leaf of the plant one is testing
and then smear the solution thinly over this surface. Allow to dry; a process
which should not take more than a few minutes in warm weather. If you are using
a lot of xylol, remember to ensure that there is good ventilation, as it is
somewhat toxic. When the smear is dry, apply a short (ca. 1 inch) piece of
clear pressure-sensitive adhesive tape over the treated area, rub gently to
make the smear adhere to the tape, lift off the impression and attach the tape,
sticky side down, to a clean slide. Examine under microscope and measure the
guard cells.
I usually measure the length of 10 separate cells per sample, average
these and by using a factor that I have previously calculated, determine the
average length of the guard cells in microns.
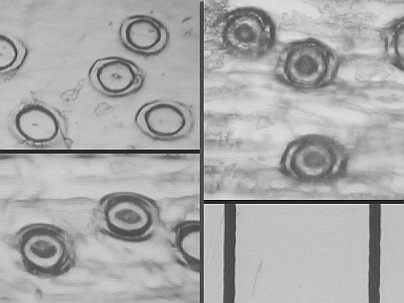
| Top left: C. Esmeralda AM/RHS - av. 27 microns across Stomatal Guard Cells | Top right: C. Lady Lucy 'Clarisse' - av. 32 microns |
| Bottom left: C. Balkis 'Nevada' - av. 33.5 microns | Bottom right: Segment of a graticule where one division equals 100 microns. |
In interpreting the results, one should be careful not to become overconfident
in the reliability of this procedure, and whilst the results can be very
useful, it is important to remember that the only way one can determine the
ploidy level of any plant with reasonable reliability, is to conduct a
chromosome count, a much more difficult procedure.
Why use this procedure at all ? Basically, it is an easy method of
improving the accuracy of one's educated guesses with regard to the ploidy
level of any plant, and it is much more accurate than the educated guesses that
some people suck out of their thumbs.
It is possible to make an eyepiece micrometer graticule at home. On slide
film, photograph, sightly overexposed, a 100 unit-long line, hand-drawn or
computer-printed on paper, so that the line takes up approximately one third of
the height of the view field of a 35 mm camera. When the film is developed,
mark a circle on the film around the photographed scale, of a size that will
fit closely into the tube of your microscope eyepiece (usually about 20 mm.
diameter), using something like a suitable sized coin as a template, cut it out
and fit it into your eyepiece so that it lies flat on the field restrictor. It
can then be used as a micrometer and if you want to calibrate it, you will need
to borrow a stage micrometer to do the job.
Although guard cell measurements are not used much in any field of research, I
was interested to find an abstract on the Web, at http://deserttortoise.org/abstract/abstracts1998/abs26.html concerning work done on the palaeoecology of Larrea by Kimberly L. Hunter of Salisbury State University, MD, from which I have
drawn the following two interesting quotes:
"Three morphologically cryptic ploidy levels have been identified by Yang in L. tridentata (creosote bush): diploid (2n=26) in the Chihuahuan Desert, tetraploid (2n=52) in the Sonoran Desert, and hexaploid (2n=78) in the Mojave Desert. We determined ploidy levels within the southwestern deserts using guard cell size, based on work of J. Masterson."
and
"Polyploidy changes across the range of L. tridentata from the end of the glacial maximum to the present were inferred from measurements of guard cells of Larrea leaves preserved in pack rat middens, in the warm deserts of North America. Diploids and tetraploids were both present in the lower Colorado River Valley 10,000 to 21,000 years before present (B.P.), and were replaced by tetraploids and possibly hexaploids before 8,100 yr B.P. Hexaploids were present in the Mojave Desert by 6,500 yr B.P."
An addendum to this page offering some hi tech options, as well as links, is accessed by
© 2003 Greig Russell
This page was updated 20th November 2004.