 alarmas en rob�tica neuroetol�gica
alarmas en rob�tica neuroetol�gica
Definici�n: La voz neuroetolog�a es una derivaci�n de "etolog�a" (Lorenz, 1981), que es el estudio de la conducta, en sus ambientes naturales, de animales viviendo su existencia diaria. Se denomina entonces neuroetolog�a (Camhi, 1984) al estudio de los mecanismos neuronales subyacentes asociados a esa conducta natural. Se define neuroetolog�a computacional (Beer, 1991) al modelado por computadora del control neural de la conducta de animales provistos de sistemas nerviosos, empezando por los m�s simples. Se puede denominar neuroetolog�a rob�tica (Beer, 1992) a la rama del control e instrumentaci�n dedicada al dise�o de "gerentes artificiales" o gestores artificiales con capacidad de acci�n aut�noma y flexible, orientada a la aplicaci�n de las ense�anzas de la neuroetolog�a computacional.
Daniel Dennett (1978) - en su texto Why not the whole iguana - recomienda encarar el modelado - ya no de sub-sub-sistemas de la v�a l�gico - simb�lica para acceder al ideal de la inteligencia artificia - sino de creaturas cognitivas completas, que podr�an ser iguanas marcianas con tres ruedas. Luego, en 1987, lo piensa mejor y predica apartarse de la profundizaci�n en microcompetencias humanas como el juego del ajedrez para encarar la macrocompetencia de animales simples (ya no extra�as iguanas extraterrestres), macrocompetencia que busca verlos como totalidad generadora de conductas.
Mientras tanto Hopfield (1982, 1984) presenta sus circuitos neurales basados en integradores operacionales como modelos de neuronas, integradores cuya entrada posee una descarga a tierra conectada en paralelo a trav�s de una bater�a y una conductancia, con las cuales este autor da cuenta de las propiedades de las membranas neuronales.

- En la Fig. 2.4 se representa el circuito el�ctrico que sirve de correlato a la membrana neuronal externa - se trata de un circu�to conductancia R - condensador o bater�a C en paralelo. La idea es de Hopfield.
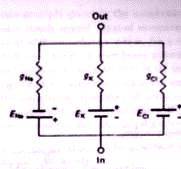
- En la Fig 2.3 Kandel y Schwartz(1985) simulan el potencial de descanso mediante un circu�to especial. Este esquema descarta muchos aniones y cationes y se concentra en el papel del i�n sodio Na
Como resultado de los flujos difusivos durante el estado de reposo la neurona est� cargada negativamente. Al salir del estado de reposo, algunas neuronas hacen una excursi�n que las vuelve transitoriamente m�s negativas, con efectos inhibitorios. Esta es la acci�n inhibidora que se marca con un c�rculo en los diagramas. Otras posibilidades diferentes son las de hacer una excursi�n que las vuelve muy positivas, lo cual es t�pico de las neuronas excitatrices. La acci�n excitadora se marca con un tri�ngulo. Los cambios en el potencial de la membrana se indican en la Fig 2.5.
Fig.2.5.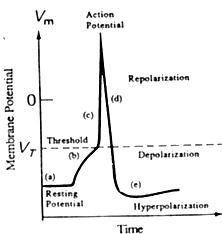
- Fig 2.5 - Durante un t�pico disparo neuronal, los cambios en el potencial de membrana en las diferentes etapas, permiten diferenciar (a) una zona de potencial de descanso, (b) una zona de despolarizaci�n resultante de la aplicaci�n de una corriente externa, (c) una zona que supera al umbral VTen que se produce el disparo y el cambio deviene explosivo. El m�ximo marca el potencial de acci�n del disparo. El proceso se reversa bruscamente y decae en (d), mientras sigue efimeramente en la zona de repolarizaci�n, con tendencia a la despolarizaci�nl y avanza hacia la zona de hiperpolarizaci�n (e). menor a�n que el mismo potencial de descanso, as�ntota a la cual tiende suavemente, (a).
Fig. 2.6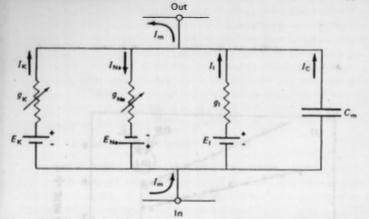
- Fig 2.6 - Un diagrama m�s completo es el que incorpora una bater�a como el que propon�a Hopfield, reemplazando al i�n cloro por una corriente m�s generalizada que abarca todas las influencias el�ctricas activas que surgen en la membrana, que no sean ni el flujo del i�n sodio ni el del i�n potasio, ambos anotados como conductancias variables.
Fig. 3.2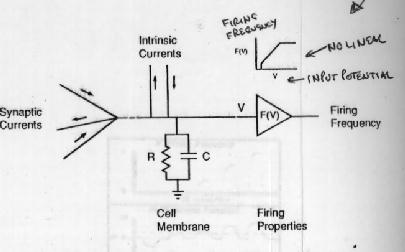
- Fig 3.2 - Simplificando por analog�a con Hopfield (Fig 2.4) se obtiene este dise�o, donde la gran novedad es la de establecer nuevas corrientes intr�nsecas de entrada o de salida que pueden excitar o inhibir y que simulan el saldo no contabilizado por la toma a tierra RC de Hopfield. Esas corrientes intr�nsecas no siempre son constantes y su modulaci�n tiene varios resultados diferentes
MODELO MATEMATICO
El valor del potencial VN (t) de cada neurona N tiene la siguiente forma general

con la siguiente nomenclatura (en ingl�s):

y conocido dicho valor se lo transforma (mediante el integrador operacional) en frecuencia de disparo, segun el modelo no lineal que aparece en el �ngulo superior derecho de la fig. 3.2.
Una red es un conjunto que posibilita posibles c�lculos simult�neos con estas ecuaciones y gr�ficos del tipo de �stos del �ngulo superior derecho. Una ecuaci�n diferencial es una neurona de la fig. 3.2 y de los diversos mapas de redes neurales que aparecen a continuaci�n como Figs I, II y II..
Las ecuaciones del modelo matem�tico se resuelven integr�ndolas por el m�todo de Euler con pasos de aproximadamente 5 milisegundos.Cada cuatro (en general N) pasos se actualiza el modelo f�sico y gr�fico del insecto, modelo que aparece en pantalla, situado en un ambiente que se puede alterar a voluntad, siempre en dos dimensiones tanto para el ambiente como para el insecto.
ALGUNAS SUBRREDES COMPORTAMENTALES
Simplemente para indicar temas en los que Randall Beer result� inspirador para nuestro trabajo, a continuaci�n aparece un ejemplo (Fig I) de una taxa (reconocer por el gradiente la presencia de alimento) y otro (Figs II y III) de un generador de patrones de acci�n fija (permitir el andar de las seis patas del insecto con seis marcapasos).
EJEMPLOS DE CIRCUITOS NEURONALES CON TAXAS, GENERADORES DE PATRONES Y QUE IMPLICAN ALARMAS
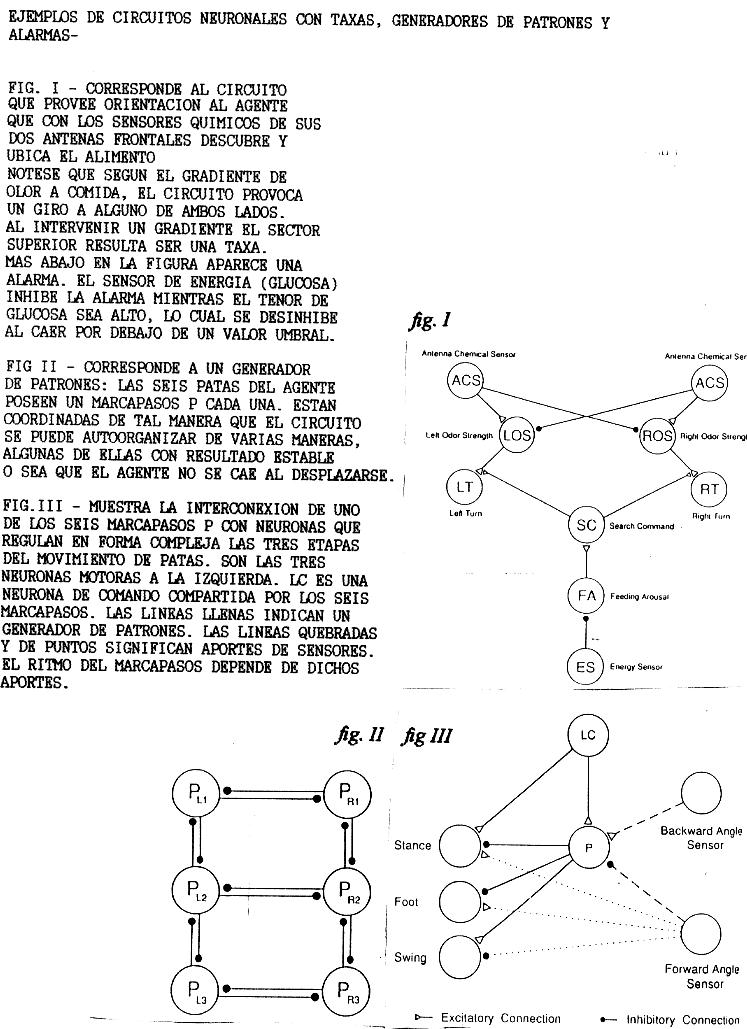
ALARMAS
Daniel Dennett en su libro Conciousness Explained retoma un tema sobre el cual hab�a insistido fuertemente en publicaciones previas. El animal con cerebro en evoluci�n consigue ir armando sistemas de alarma que lo mantienen innatamente prevenido de riesgos y depredadores, as� como de no dejar de aprovechar innatamente circunstancias favorables. Esas alarmas son asimismo la base de todo sistema neo-cortical superior, arrastrando entonces la misma configuraci�n y recursos previos de alarmas superadas por las condiciones cambiantes del ecosistema al cual el animal se hab�a adaptado. Si se parte de la regla biol�gica de que todo sistema nervioso ya superado por cambios en el ambiente, es evolutivamente aprovechado por una nueva configuraci�n neuronal para enfrentar nuevos problemas del ambiente, es v�lido pensar lo mismo con respecto a las alarmas neuroetol�gicas.
Nuestro trabajo se basa en hipotetizar que los sistemas evolutivos de alarmas neuroetol�gicas tienen algunos patrones en com�n que permiten encontrar explicaciones generales para los sistemas neuroetol�gicos asociados con las funciones neo-corticales superiores. La teor�a de las alarmas constru�da tendr�a claras consecuencias sobre la fabricaci�n de sistemas de automatizaci�n y control industriales, con atributos del tipo del que se pretender�a de un gestor o "gerente artificial", esto es, con suficiente autonom�a frente a situaciones inesperadas.
Como consecuencia del estudio previamente explicado, podemos formular respuestas, bastante significativas pero tentativas, a una serie de importantes preguntas que buscan dejar en claro algunas conclusiones generalizantes.
1) �Qu� similitud tienen los sistemas nerviosos de diferentes animales, con especial referencia a los sistemas de alarma?
- Alarmas por la detecci�n de alimento y de agua
- Alarma de saciedad
- Alarmas asociadas al desplazamiento hacia el alimento.
- Alarma para enfrentar o huir o transformar enfrentamiento en huida y viceversa.
- Alarma por dejar de aprovechar una opci�n de reproducci�n (actitud masculina) o de dejar de verificar aptitud del cortejante (actitud femenina).
- Circuitos de alarma similares en reflejos, taxas y generadores de patrones de conducta.
- "Ciclos decisionales neuroetol�gicos".
Respuesta tentativa: Todos los sistemas nerviosos de animales poseen:
2) �Qu� concepto neuroetol�gico es m�s fundamental y general que la inteligencia?
Respuesta tentativa: El de flexibilidad que conduce a la autonom�a (independencia de un reglamento externo) en los seres vivientes, sobre todo en los provistos de sistemas nerviosos.
3) �Cu�l es la relaci�n entre la inteligencia y las alarmas evolucionadas?
Respuesta tentativa: La inteligencia (excluyendo de ella el aprendizaje y la memoria) es una conducta adaptiva. Ella es respuesta flexible a las alarmas resultantes de condiciones complejas en ambientes complejos.
4) �Cu�les son los mecanismos neuroetol�gicos que respaldan una conducta inteligente?
Respuesta tentativa: Son mecanismos que le permiten al agente la caracterizaci�n borrosa o n�tida de su objetivo en cada momento pero no incluyen mecanismos para caracterizar la trayectoria intencional previa o posterior. Incluyen alarmas de tipo �tico y dentro de la ley aceptada. El plan general cumple un papel m�nimo y la ejecuci�n del objetivo inmediato cumple un papel m�ximo durante la actividad del agente.
5) �Es necesaria o n� la apelaci�n a las representaciones internas de los objetivos generales para la actividad de un agente inteligente?
Respuesta tentativa: Continuando la respuesta previa, la acci�n del agente est� situada, o sea que resulta contingente con la situaci�n actual que se est� desenvolviendo. No necesita haber - y quiz�s no hay - una prescripci�n a priori de c�mo se va a desarrollar la interacci�n con el mundo real. A posteriori las funciones intelectuales superiores omiten - en general - las motivaciones cruciales, rescatan las motivaciones y acciones de cada momento y racionalizan o disfrazan toda la acci�n en una s�ntesis con la cual aparece que hubiese habido un plan a satisfacer. El pensamiento primitivo ignora en general esta s�ntesis, que es m�s bien caracter�stica del pensamiento culto, entrenado en sacar conclusiones y presentar s�ntesis. El pensamiento culto va torciendo al pensamiento primitivo de manera que se unifique la PLANIFICACION con la EJECUCION.
6) Desde un comienzo, �c�mo se logran las estructuras adecuadas para conseguir una din�mica de mecanismos internos?
Respuesta tentativa: A esta pregunta corresponde una contestaci�n hist�rico-evolucionaria, que rescata de estadios primitivos estructuras que por azar resultaron adecuadas y sobreimpone a ellas otras nuevas estructuras tentativas, siempre basadas en circuitos que las contingencias del ambiente hicieron quedar en desuso.
7) �C�mo se consigue hacer aparecer una conducta orientada hacia objetivos?
Respuesta tentativa: A trav�s de mecanismos que no incluyen la representaci�n interna de la situaci�n del agente inteligente, a la manera del cerebro-B de Marvin Minsky que no participa directamente de las percepciones del cerebro-A del mismo autor. La conducta orientada hacia objetivos se deriva m�s bien de algunos estados o din�micas internas del sistema nervioso del agente que modulan las interacciones entre las diferentes conductas (A y B) posibilitadas por las subrredes neurales (A y B) existentes.
8) En el caso de un agente computacional en una computadora, �c�mo se logra reproducir en �l una conducta inteligente?
Respuesta tentativa: Empezando por seres simples provistos de sistemas nerviosos esquem�ticos, tales como los buscados en estudios como el que aqu� se insin�a, esto es, con robots bidimensionales imitando invertebrados hexap�dicos (o sea insectos con 6 patas). Los seres humanos, con todas sus posibilidades y restricciones conductuales (�ticas, etc.), son, en pocas palabras, demasiado complejos para imitar el dise�o de sus mecanismos internos.
9) �C�mo una descripci�n simb�lica externa al agente, del tipo IF... THEN..., se correlaciona con una conducta interna de una red neural que genera la conducta del agente?
Respuesta tentativa: En algunos casos es muy problem�tico, como en el caso de la autoorganizaci�n del andar o aire de un desplazamiento del agente, que es funci�n de la velocidad, de la posici�n inicial de las patas, de los aportes de sensores (ver Fig III), etc., Estos resultan ser datos b�sicos para que los marcapasos se autoorganicen sin mucha posibilidad de acierto por parte de una regla l�gico- simb�lica, que no puede contemplar todos los casos posibles.
10) �Cu�les son los mecanismos internos por los cuales aparecen transiciones de conducta gatilladas por condiciones iniciales internas al agente?
Respuesta tentativa: No se pueden formular reglas generales. Cada neurona exhibe sus propias din�micas. Existen efectos de avanacci�n y de retroalimentaci�n. Aparecen sinapsis modificatorias, en forma brusca, de conductas. Hay hormonas y neurohumores bioqu�micos.
11) �C�mo se implementan las bifurcaciones en una decisi�n hacia una posibilidad u otra?
Respuesta tentativa: Sin una neurona de comando; en cambio, con aportes no-jer�rquicos de diferentes subrredes
12)�Qu� aplicaciones tiene?
reflejos, taxas, generadores de patrones de conductas de acci�n fija
la vera efigie de Periplaneta computatrix con 78 "neuronas", o sea, los c�rculos coloreados
simulaci�n electr�nica de cada una de las "neuronas" de Periplaneta computatrix
Respuesta tentativa: Directas, en el dise�o de robots o de sistemas autom�ticos con esbozos de capacidad gerencial aut�noma, sujeta a restricciones o alarmas
BIBLIOGRAFIA
Simon H - Las Ciencias de lo Artificial
Tank DW, Hopfield JJ - Investigaci�n y Ciencia (1984)
Hopfield JJ, Tank DW - Biological Cybernetics, 52, 141-152
Dennett D - Conciousness Explained
Beer R - Intelligence as Adaptive Behavior: An experiment in Computational Neuroethology - Academic Press (1990)
Beer R, McKenna Th - Invertebrate Neuroethology and Robotics - Academic Press (1992)
Williams P, Williams G - Programa Neural Systems (NS), en Shareware, Rt 1, Box 302, Gravel Switch, KY 40328, con explicaciones para su ejecuci�n.
Leggett DJ - Instrumental Simplex Optimization - Journal of Chemical Education - 60,9,1983, p. 707
Crick F, Ansanuma C-en Rumelhart y McClelland (eds) Parallel Distributed Processes, Vol 1, (1986) MIT Press, p. 333
Lorenz KZ - The Foundations of Ethology - Simon & Shuster (1981)
Camhi JM - Neuroethology. Sinauer Associates (1984)
Becke CH von der - Optimizaci�n de Suprasistemas, UTN, (1989)
Becke CH von der, Barral R y Motto A- Neuronal Processing Systems - I. Explorations on Cortical Processing with Non Hierarchical Structures, Medyfinol (1994)
Becke CH von der y Contou Carrere JE -Control por inteligencia artificial Alimentaci�n latinoamericana (1988)
{kind=link}
29.jun.1999
Pulsar tecla de vuelta
Temas de investigaci�n de Carlos von der Becke y colaboradores, en el Glosario del primero de los nombrados.