![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()

introducción a las arqueas - bacteria halofila
bacteria halofila en ingles - aplicaciones biotecnologicas
(extraído de www.biologia.edu.ar )
El grupo más antiguo, las
arqueobacterias, son un conjunto de organismos y por sus especiales
características se considera que conforman un Dominio separado: Archaea.
Fenotípicamente, Archaea son muy parecidos a las Bacterias. La mayoría son
pequeños (0.5-5 micras) y con formas de bastones, cocos y espirilos. Las Archaea
generalmente se reproducen por fisión, como la mayoría de las Bacterias. Los
genomas de Archaea son de un tamaño sobre 2-4 Mbp, similar a la mayoría de las
Bacterias. Si bien lucen como bacterias poseen características bioquímicas y
genéticas que las alejan de ellas. Por ejemplo:
Por habitar ambientes "extremos", se las conocen también con el nombre de extremófilas.
Se considera que las condiciones de crecimiento semejan a las existentes en los primeros tiempos de la historia de la Tierra por ello a estos organismos se los denominó arqueobacterias (del griego arkhaios = antiguo).
 Los lípidos presentes en las membranas son únicos desde el punto de vista
químico, a diferencia de los eucariotas y las bacterias, en que los enlaces
éster son los responsables de la unión entre los ác. grasos y glicerol, los
lípidos de las Archaea poseen enlaces ÉTER para la unión del glicerol con
cadenas laterales hidrofóbicas. En lugar de ac. grasos poseen cadenas laterales
formadas por unidades repetitivas de una molécula hidrocarbonada como el
isopreno.
Los lípidos presentes en las membranas son únicos desde el punto de vista
químico, a diferencia de los eucariotas y las bacterias, en que los enlaces
éster son los responsables de la unión entre los ác. grasos y glicerol, los
lípidos de las Archaea poseen enlaces ÉTER para la unión del glicerol con
cadenas laterales hidrofóbicas. En lugar de ac. grasos poseen cadenas laterales
formadas por unidades repetitivas de una molécula hidrocarbonada como el
isopreno.
Los principales tipos de lípidos son los diéteres de glicerol. En algunos éteres las cadenas laterales (fentanil) se unen entre sí por enlaces covalentes formando una monocapa en lugar de la bicapa característica de las membranas, siendo más estables y resistentes, siendo habituales por lo tanto en las hipertemófilas.
Algunas arqueobacterias metanogénicas poseen la pared celular formada por un compuesto similar al peptidoglicano de las bacterias, por lo que denomina pseudopeptidoglicano, con enlaces glucosídicos 1,3 en lugar de los 1,4 de los peptidoglicano. En otras archaeas la pared se compone de polisacaridos, glicoproteínas o proteínas.
El tipo de pared más común es la capa superficial paracristalina (capa S) formada por proteína o glucoproteína, de simetría hexagonal.
La pared celular impide la lisis celular y le confiere la forma a la célula. Las paredes de las Archaea son resistentes naturalmente a la lisozima, debido a la ausencia de peptidoglicano.
La única arqueobacteria que carece de pared es Thermoplasma.
Sobre la base del análisis de la subunidad pequeña del ARN, las Archaea consisten en dos grupos filogenéticamente diferentes: Crenarchaeota y Euryarchaeota. Se diferencias por el tipo particular de ARN que presentan y por el ambiente en que habitan. Las Crenarchaeota (crenotas) es un grupo fisiológicamente homogéneo de hábitats enteramente termofílicos. en cambio las Euryarchaeota (euryotas) son un grupo fenotípicamente heterogéneo, que incluye a las metanogénicas, halófilas, etc.
Basados en su fisiología se distinguen:
Además de las características unificadoras de las arqueobacterias, (pared celular sin mureína, lípidos de membranas con enlaces éter, etc.), estos procariotas exhiben atributos bioquímicos que le permiten adaptarse a estos ambientes extremos. Las Crenarchaeota son principalmente hipertermofílicos dependientes del sulfuro y los Euryarchaeota son metanogénicos y halófilos extremos.

![]()



(extraído de http://bioinformatica.uab.es/biocomputacio/treballs98-99/Jara/web4.htm )
Las arqueobacterias halofílicas son un grupo diverso de microorganismos que habitan en ambientes salinos.Viven en ambientes naturales donde la concentración de sal es muy alta (hasta 5 molar o 25 por ciento de NaCl). Estos procariotas requieren la sal para el crecimiento, sus paredes celulares, ribosomas y enzimas, se estabilizan con el ión Na+.
El término halófilo extremo se utiliza para
indicar que estos organismos no solamente son halofílicos, sino que su
requerimiento de sal es muy alto, en algunos casos cercano a la saturación. Un
ejemplo de hábitat extremadamente salino son las salinas marinas: pequeños
depósitos llenos con agua de mar que se dejan evaporar, produciendo NaCl y otras
sales de valor comercial.

A medida que estas salinas se aproximan a los límites mínimos de salinidad para los halófilos extremos, las aguas se tornan rojizas púrpura, color que indica el desarrollo masivo, llamado floración de arqueobacterias halofílicas
las arqueobacterias halófilas pueden obtener energía a partir de la luz mediante un proceso que no es la fotosíntesis. Cuando baja la tensión de oxígeno al evaporarse el agua la bacteria exporta a la membrana celular una proteína llamada bacteriorrodopsina que lleva unido un grupo cromóforo carotenoido : el retinal que es de color púrpura.
Producción de bacteriorrodopsina
en
condiciones de baja aireación las halófilas extremas sintetizan e insertan
dentro de sus membranas una proteína denominada bacteriorrodopsina (similar
al pigmento del ojo, la rodopsina). Conjugada con la bacteriorrodopsina
está una molécula de retinal (parecida al carotenoide) que puede absorber la luz
y catalizar la transferencia de protones a través de la membrana citoplásmica.
Por su contenido en retinal, la bacteriorrodopsina es de color púrpura y las
células de Halobacterium adaptadas a un desarrollo en condiciones de
aireación altas, al verse en condiciones limitadas de oxígeno, cambian
gradualmente de un color naranja o rojo a uno más púrpura rojizo debido a la
inserción dentro de la membrana citoplásmica de bacteriorrodopsina.
La
bacteriorrodopsina es miembro de una familia de proteínas de membrana de
estructuras similares, pero con funciones diferentes. Así por ejemplo, el
receptor de luz rodopsina de los bastones de la retina de los vertebrados, y
algunos receptores proteicos celulares de superficie que unen hormonas
específicas, estan plegadas en siete hélices alfa transmembrana (al igual que la
bacteriorrodopsina) y no actúan como transportadores sino como transductores de
señales.
Bacteriorrodopsina y síntesis de ATP mediada por la luz
El retinal de la bacteriorrodopsina esta en reposo en forma trans cuando es excitada por la luz, lo que provoca su paso a forma cis con la consiguiente liberación de un protón hacia el exterior de la membrana. Entonces el retinal tiene tendencia a volver a la forma trans cogiendo un protón del citoplasma. De esta manera se consigue crear un gradiente de protones que será utilizado por una ATPasa de membrana para obtener ATP. bacteriorodopsina que reacciona con la luz formando un gradiente de protones a lo largo de la membrana que permite la síntesis de ATP. Este es el único ejemplo en la naturaleza de una fotofosforilación sin clorofila
tabla con el contenido de sales del medio e interior de una halobacteria
|
Ión |
Concentración del medio (M) |
Concentración del interior celular (M) |
|
Na+ |
3.3 |
0.8 |
|
K+ |
0.05 |
5.3 |
|
Mg2+ |
0.13 |
0.12 |
|
Cl- |
3.3 |
3.3 |

extraido textualmente de http://www.biochem.mpg.de/oesterhelt/genomics/Intro_Hsal.html pronto espero traducirlo y acortarlo, toda ayuda para la traducción es bien recibida ;)
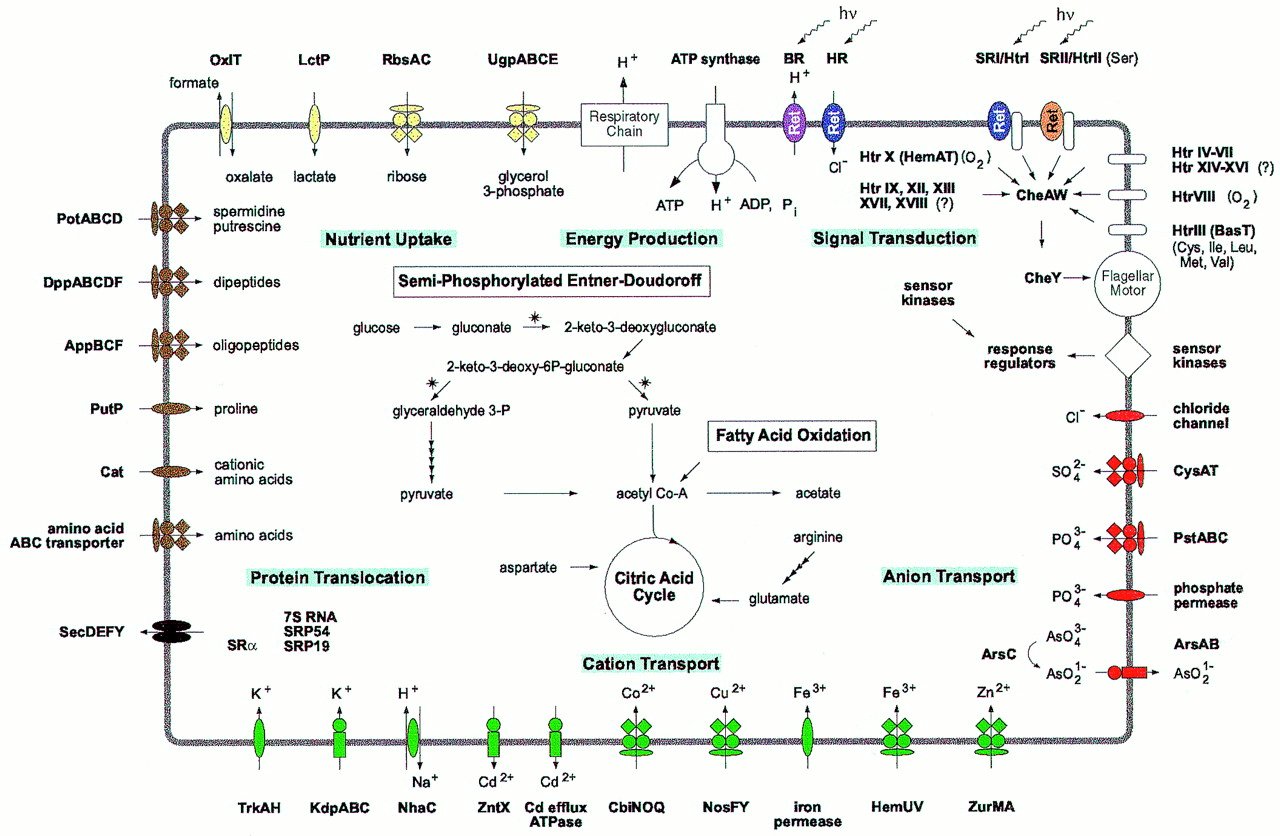
Halobacterium salinarum is a model organism for the halophilic branch of the archaea. It lives in highly saline environments (4M salt and higher) and is one of the few species known that can live in saturated salt solutions. Mass cultures of Halobacterium salinarum as shown in the pictures below can be recognized by their typical color, which originates from bacterioruberins. Halobacterium salinarum is depicted in its natural environment and as a species that colonizes salines.
It can live with light as only energy source due to the activity of the retinal protein bacteriorhodopsin, a light-driven proton pump, which has been studied in great detail and has become a paradigm of membrane proteins in general and transport proteins in particular. From this point, our focus has widened to study additional processes in which retinal proteins are involved: the energy metabolism of Halobacterium salinarum and the tactic responses with its associated signal transduction network.
Since many years, research in the department of membrane biochemistry at the Max-Planck-Institute of Biochemistry concentrates on the biology of Halobacterium salinarum.
Retinal proteins of Halobacterium salinarum
Halobacterium salinarum contains four retinal proteins, which are photosynthetic pigments with a retinal chromophore involved in light energy conversion and signal transduction.
The four retinal proteins are
Energy metabolism
Halophiles, unlike their closest relatives, the methanogenes, can grow under aerobic and anaerobic conditions.
Halobacterium has three distinct systems to gain energy.
Response to external stimuli (signal transduction)
Halobacterium is a flagellated organism which shows tactic behaviour. Besides being able to detect essential amino acids (chemotaxis) and osmotically active compounds (osmotaxis), it can respond to light (phototaxis) and can sense oxygen (aerotaxis).
The signal transduction cascade starts with the receptor/transducer, which may be composed of two distinct proteins or may be a single protein. The signal is forwarded to the switch of the flagellar motor through a two-component regulatory system consisting of the histidine kinase cheA and the response regulator cheY. During relay of the signal it is amplified and different signals are integrated. Adaptation involves methylation and demethylation of the transducer proteins by cheR (methyltransferase) and cheB (a regulated methylesterase). Genome analysis shows that Halobacterium contains 18 distinct transducers, indicating that it can sense a large number of distinct stimuli. Currently, stimuli are known for seven of these transducers.
----------------------------------------------------------------------------------------------------------------
Bacteriorhodopsin
I. Introduction
The retinal
protein bacteriorhodopsin is the major photosynthetic protein of the archaeon
Halobacterium salinarum. It converts the energy of "green" light
(500-650 nm, max 568 nm) into an electrochemical proton gradient, which in
turn is used for ATP production by ATP synthase. It functions as a
light-driven proton pump, transporting protons out of the cell, and
exemplifies vectorial catalysis.
Bacteriorhodopsin is the focus of much interest and has become a paradigm for
membrane proteins in general and transporters in particular. Its structure and
function have been analyzed in great detail using a multitude of different
experimental techniques and has become the best-understood example of
vectorial catalysis.
The reversible light-triggered color-change has allowed to develop
biotechnological applications of bacteriorhodopsin, e.g. its application in
optical information recording.
The further description is based on the review article by
Haupts, Tittor, and Oesterhelt (1999),
Oesterhelt (1998) and
Oesterhelt(1999).
II. Structure and function of bacteriorhodopsin
Bacteriorhodopsin - as all retinal proteins from Halobacterium - folds into a seven-transmebrane helix topology with short interconnecting loops. The helices (named A-G) are arranged in an arc-like structure and tightly surround a retinal molecule that is covalently bound via a Schiff base to a conserved lysine (Lys-216) on helix G. The cross-section of BR with residues important for proton transfer and the probable path of the proton is shown in Fig.1. The 3D structure is accessible through the PDB database, entry 1BRR. Fig. 2 shows a model of the sequence.
Retinal separates a cytoplasmic from an extracellular half channel that is lined by amino acids crucial for efficient proton transport by BR (especially Asp-96 in the cytoplasmic and Asp-85 in the extracellular half channel). The Schiff base between retinal and Lys-216 is located at the center of this channel. To allow vectorial proton transport, de- and reprotonation of the Schiff base must occur from different sides of the membrane. Thus, the accessibility of the Schiff base for Asp-96 and Asp-85 must be switched during the catalytic cycle. The geometry of the retinal, the protonation state of the Schiff base, and its precise electrostatic interaction with the surrounding charges (Asp-85, Asp-212, Arg-82) and dipoles tune the absorption maximum to fit its biological function.
III. The purple membrane
The surface
of Halobacterium salinarum contains membrane patches called the purple
membrane. The protein:lipid ratio is 75:25. The only protein in the purple
membrane is bacteriorhodopsin which forms a hexagonal 2-dimensional crystal
consisting of bacteriorhodopsin trimers.
The purple membrane can be easily isolated and permits mass production of
bacteriorhodopsin as is required for biotechnological applications.
IV. The catalytic cycle of bacteriorhodopsin
Absorption of a photon by bacteriorhodopsin initiates a catalytic cycle that leads to transport of a proton out of the cell. Several intermediates in the photocycle have been identified by spectroscopic techniques (Fig.3). By application of a multitude of biophysical techniques, the exact nature of the changes in each step of the cycle has been determined and has been related to transport function.
The cycle can be formally described in terms of six steps of isomerization (I), ion transport (T), and accessibility change (switch S). Retinal first photo-isomerizes from an all-trans to a 13-cis configuration followed by a proton transfer from the Schiff base to the proton acceptor Asp-85. To allow vectoriality, reprotonation of the Schiff base from Asp-85 must be excluded. Thus, its accessibility is switched from extracellular to intracellular. The Schiff base is then reprotonated from Asp-96 in the cytoplasmic channel. After reprotonation of Asp-96 from the cytoplasmic surface, retinal reisomerizes thermally and the accessibility of the Schiff base switches back to extracellular to reestablish the initial state. These steps represent the minimal number of steps needed to account for vectorial catalysis in wild-type bacteriorhodopsin.
V. The catalytic cycle step by step
Dynamic structural changes occuring in chromophore and protein during the light-induced reaction cycle can be detected either directly by time-resolved spectroscopic techniques (ultrafast laser spectroscopy, flash photolysis, ESR spectroscopy, FTIR spectroscopy) or by trapping intermediate states, determining their structures by static methods (NMR spectroscopy, electron microscopy, neutron scattering) and comparing it with the ground state.
In a stereoselective process, all-trans retinal is photoisomerized to 13-cis retinal. This process has been time-resolved to few femtoseconds. Within 500 fs, all-trans retinal isomerizes to 13-cis retinal, resulting in J600 which is converted to K590 within another 5 ps.
The K590 intermediate is transformed to the L550 intermediate within 2 µs. The hydrogen bonding interaction in the extracellular channel between the protonated Schiff base and Asp-85, which involves a water molecule, is strengthened.
The M state is reached from the L state within several microseconds. This transition involves transfer of a proton from the Schiff base to Asp-85 in the extracellular half-channel.
To allow vectorial proton transport, de- and reprotonation of the Schiff base must occur from different sides of the membrane. This accessibility switch occurs at the level of the M intermediate: M410(EC) to M410(CP). Thus, the originally described "M" intermediate is in fact split into two or more different intermediates all having yellow color.
Reprotonation of
the Schiff base from Asp-96 in the cytoplasmic half-channel occurs during
transformation from the M410(EC) to the N560 intermediate within milliseconds.
Reprotonation of Asp-96 by a proton from the cytoplasm also occurs during the
lifetime of the N560 intermediate.
It should be noted that Asp-96 functions as a proton storage for
reprotonation of the Schiff base. Therefore, the proton does not originate
directly from the cytoplasm. This detail solves the puzzling phenomenon that
the transport rate of this proton transporter is not pH-dependent (within
limits).
The transition of the N560 to the O640 intermediate is the thermal 13-cis to all-trans isomerization of retinal in the environment of protonated Asp-96 and protonated Asp-85.
Deprotonation of Asp-85 completes the catalytic cycle. Switching the accessibility of the Schiff base back from extracellular to intracellular occurs within ca 5 ms and results in restoration of the initial state.
VI. Site-directed mutagenesis of bacteriorhodopsin
An important tool for these studies, and also for spectroscopic investigations on structure and dynamics, is the possibility to produce specifically modified proteins by site directed mutagenesis and homologous overexpression. By this method the role of individual amino acid residues for the transport mechanism can be investigated, or reporter molecules can be introduced at certain positions. Several mutants interfere with the photocycle and and proton transport and may permit to trap intermediates of the catalytic cycle. The direct involvement of Asp-85 and Asp-96 in proton transfer has been demonstrated by analysis of mutants.
VII. Bacteriorhodopsin as material for optical information recording
Biotechnological applications on the basis of the colour change between purple
and yellow (long living intermediate M) is the basis for using of
bacteriorhodopsin for optical information recording. The technique has
advanced such that it could be used as a safety feature on chipcards.
For further details see
Prof. N. Hampp, University of Marburg
(external link)
VIII. Nanotechnology applied to the analysis of bacteriorhodopsin
Atomic
force spectroscopy has been used to handle single molecules of
bacteriorhodopsin in the purple membrane. This has permitted to measure the
force required to pull individual helices out of the membrane.
One of the molecular prerequisites for these experiments was the introduction
of Cys residues into bacteriorhodopsin by site-directed mutagenesis.
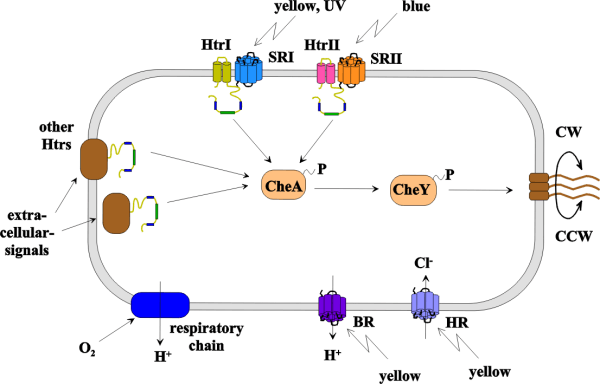
Investigation of the function of halobacterial transducer proteins (Htrs) in the signaltransduction-machinery of Halobacterium salinarum
To sense its
environment the halophilic archeaeon Halobacterium salinarum employs a
sophisticated protein-machinery consisting of several types of components:
External signals (light, chemicals, oxygen etc.) interact with receptor proteins
which can be light-receptors (like in the case of sensory rhodopsins I and II),
or binding proteins for chemicals (like BasB or CosB) (Kokoeva,
2000 and
Kokoeva, 2002) In other cases
the receptors can simultaneously be transducers (like HemAT which is thought to
bind oxygen with its heme-group (Hou,
2000).
These receptor proteins forward the external signals via
protein-protein-interactions to halobacterial transducer proteins (Htrs). The
Htrs - of which 18 have been found in H. salinarum by homology studies -
transmit the external signal to an intracellular signaling pathway of Che-proteins
starting with the autophosphorylation of the histidine kinase CheA (Rudolph,
1995) which then phosphorylates the response
regulator CheY. Phosphorylated CheY eventually induces a switch at the flagellar
motor causing a reversal of flagellar rotational direction. This makes the cell
reverse its swimming direction.
To enable the cells to adapt to a certain signal intensity and to respond only
to changes in signal strength (e.g. light intensity or substrate concentration)
the Htrs are subject to methylation/demethylation of glutamate residues which
are located in methylatable regions of the Htrs within a long helical coiled
coil structure, as shown for Tsr, an Htr homolog in E. coli (Kim,
1999). Adaptation of the cells is achieved by
changing the methylation state of the Htrs in response to external signals via
the actions of the methyltransferase CheR and the methylesterase CheB (reviewed
in:
Marwan, 1999).
In order to elucidate which role the Htrs are playing in environmental sensing,
a deletion approach was chosen. Suicide-vectors with a mevinoline resistance
marker were used to produce deletion mutants of H. salinarum. The
deletion-strains were verified by Southern-blots and the strains were
investigated by behavioural studies (Storch,
1999 and
Kokoeva, 2000).
|
|
The picture above shows a schematic
representation of the Htr-mediated signaling in H. salinarum. In this
organism four retinal containing photoactivatable proteins are present: the
proton pump bacteriorhodopsin (BR), the chloride pump halorhodopsin (HR) and
the sensory rhodopsins I and II (SRI and SRII). SRI and SRII confer phototaxis
via the Htrs I and II respectively (reviewed in:
Marwan, 1999). BR is also
involved in phototaxis (Bibikov,
1993,
Bibikov, 1991) but the
mechanism is still unknown.
Other Htrs were shown to be involved in chemotaxis. The cytoplasmic protein
Car (Storch,
1999) senses arginine, membrane spanning BasT (Kokoeva,
2000) confers chemotaxis towards the aminoacids
Met, Cys, Val, Leu and Ile via interaction with the binding protein BasB. CosT
together with its binding protein CosB mediates chemotaxis towards compatible
solutes like glycine betaine (Kokoeva,
2002). Htr8 and HemAT (Hou,
2000) were found to be involved in oxygen-taxis.
The elucidation of the functions of the other Htrs in H. salinarum is
one of the topics we are currently working on in our group.
(extraído de http://zdna2.umbi.umd.edu/~haloed/ )

La biotecnología es "el uso de la ciencia y de la ingeniería al uso directo o indirecto de los organismos vivos, o partes de los productos de organismos vivos, en sus formas naturales o modificadas."
1. Diseño de antibióticos nuevos.
2. Producción de las vesículas recombinant del gas para la separación, la presentación del antígeno y el desarrollo vaccíneo.
3. Producción industrial de biopolímeros y enzimas
4. Ingeniería genética de las plantas para aumentar su tolerancia de la sal.
5. Desarrollo de las enzimas estables para la catálisis
6. Uso de los proteasas para las salsas orientales de los pescados y de soja.
7. Desarrollo de las películas púrpuras de la membrana para las virutas de la bio-computadora de la olografía.
8. Extracción del caroteno para el uso como suplementos de alimentos o colorantes del alimenticios
9. Uso de los procesos metabólicos para limpiar las salmueras contaminadas del con demasiada sal
10. Uso de las lipasas para los detergentes.
---------------------------------------------------------------------------------------------------------
revista latinoamericana de microbiología, articulo cobre microorganismos halofilicos. www.medigraphic.com ==> bajar
archivos con varios apuntes que use de microbiología, encontrados en la red, con su correspondiente fuente ==> bajar