Irreducible Complexity: The (Cosmic) Emperor's New Clothes
Michael Behe, an Associate Professor of Biochemistry at Lehigh University, published a book called Darwin's Black Box in 1996. Since that time he has been at the forefront of the Intelligent Design Movement.
- We will be looking at Behe's main argument for Intelligent Design, his concept of Irreducible Complexity in the complex processes of biochemistry.
- We will see that his argument is flawed for a number of reasons:
- Irreducible Complexity is a very slippery concept.
- His argument is untestable.
- His either-or logic between ID and Darwinism is a false dichotomy.
- His claim that biochemistry has stymied Darwinian evolution is demonstrably false.
- His claim that some complex processes, such a blood clotting, could not be produced by step-by-step Darwinian evolution is wrong.
- All the six major examples have possible evolutionary precursors and some are not even irreducibly complex.
- His "super Adamite" bacteria is an impossibility.
- We then provide some conclusions on irreducible complexity as an argument for God's existence.
Behe's Irreducible Complexity
Behe's argument starts by noting that advances in biochemistry in the twentieth century, with the aid of sophisticated tools and techniques such as X-Ray crystallography, nuclear magnetic resonance and electron microscopy, have enabled the structure of the cell and its biochemical inner workings to be understood in detail for the first time. These complex processes were unknown to Darwin, who essentially treated the cell as a black box. With the advances mentioned above, the black box now stands open. Thus according to Behe, "biochemistry offers a Lilliputian challenge to Darwin." and that "Now the black box of the cell has been opened, and the infinitesmal world that stands revealed must be explained." [1]According to Behe, the main biochemical challenge to Darwin comes from irreducibly complex systems. He defines the term as such:
| By irreducibly complex I mean a single system composed of several well-matched, interacting parts that contribute to the basic function, wherein the removal of any one of the parts causes the system to effectively cease functioning. An irreducibly complex system cannot be produced directly (that is, by continuously improving the initial function, which continues to work by the same mechanism) by slight, successive modifications of a precursor system, because any precursor to an irreducibly complex system that is missing a part is by definition nonfunctional. An irreducibly complex biological system, if there is such a thing, would be a powerful challenge to Darwinian evolution.[2] |
To familize his reader with the concept, he introduced the common contraption, the humble mousetrap. According to Behe, the mouse trap has five components, the wooden platform (the base), the hammer (to crush the mouse), the spring (to provide force for the hammer), the catch (to sense the mouse) and the holding bar (to hold the hammer back while waiting for the mouse). The mousetrap is irreducibly complex because every component is required for the system to work. Once any one part is missing, the trap will fail to catch the mouse. Thus there can be no physical precustor to the moustrap that is simpler, since any step by step improvement is impossible. The system must function with all five parts or it doesn't work at all.
Next Behe goes on to give in exquisite detail six examples of irreducibly complex systems in the black box:
- The mechanics of bacterial flagellum:
The system needs three separate components working together: namely the filament (the "oar" or "propellor"), the "M" Ring (the "rotor") and a motor that draws energy from a flow of acid through the bacterial membrane.[3] - The Cilium:
The cilium, a "hairlike" looking structure that protrudes out of some cells, consists of microtubules (the strands that slide up an down), dynein (the motor that slides one set of microtubules over another allowing the cilium to bend) and nexin (which is the linker that keeps the adjacent microtubes together.) That way when dynein pushes one set of microtubules up against another, the movement is converted into a bending motion. This provides the cell with the capability to swim.[4] - Blood clotting:
The mechanics of blood clotting is a cascade in that one reaction is required to trigger another (think falling dominos) until the final step-which is the "trimming" of the protein fibrinogen into fibrin. Fibrin forms a biological "wiremesh" that entraps blod cells and keeps them from flowing out of the body. Working backwards one requires the protein thrombin to "cut" fibrinogen into fibrin. But thrombin exists in the bloodstream as the inactive prothrombin. To activate prothrombin, we need an active "Stuart factor" to cleave prothrombin and accelerin to speed up the process so that it will happen quick enough to be of any help. What activates Stuart factor and accelerin? Still more proteins! [5] - Cellular transport systems:
The mechanics of how proteins are transported within the cell is introduced next. Two forms of transport, gated transport and vesicular transport are required to get proteins from their origin in the cytoplasm [a] to the lysosome.[b]Gated transport is called thus due to it's similarity to our everyday experience of passing through a guarded (electronically or otherwise) gate. This system require three basic components to work: an identification tag, a scanner (to verify identification) and a gate (that is activated by the scanner). The system needs all three components to work otherwise it will not work. Thus in a cell, when a protein is to be manufactured, one of the first steps is for the mRNA [c] to be transported out from the nucleus into the cytoplasm. This requires gated transport of the mRNA at the nuclear pore. Proteins in the pore reads a signal from the RNA (the scanner reads the identification tag) and opens the pore (gate is opened).
Vesicular transport is even more complicated. Vesicular transport involves transporting the protein within a body (the vesicle) to another location (such as the lysosome). This requires six items to function properly, (1) an identification tag for the protein to be recognizable by the budding vesicle, (2) a transport vehicle(the vesicle), (3)a receptor or scanner in the vesicle to recognize the right protein, (4) an indentification tag for the vesicle itself, (5) a scaner on the lysosome and (6) a gate on the lysosome that it activated by the scanner.
- The immune system:
The immune system is introduced in a few separate sections. We will look at the "recognition" part-where the body's immune system recognizes a foreign invader and starts producing antibodies.The "B" cells have protruding Y-Shaped fingers (the antibody), each with many different permutations of binding sites, that enable the cell to latch on to a foreign body. Thus there are many different flavors of B cells, each with it's own binding site combination. Once the correct B-Cell latches on, it takes a piece of the foreign cell's membrane into it. Here the foreign molecule is chopped up and is stuck to a protein called MHC. This is then returned to the membrane of the Cell. Now helper-T cells come along and binds to the B cell. If it confirms that this indeed is a foreign substance, it sends a message ( by secreting a substance called interleukin) to that particular B Cell to start to reproduce. (That's the major reason why the B cell is bound to the antibody, so that the system doesn't have to go around looking for the "right" B cell to reproduce.) Eventually they start to produce "free" antibodies (those not bound to the membrane of the B cell) which are released in large amounts into the plasma. According to Behe even here, the system is irreducibly complex- you need, as a minimum (1) the membrane bound form of the antibody, (2) the messenger (such as interleukin) and (3) the exported (free) form of the antibody. Without (1) you can't find the right cell that makes the particular antibody. Without (2), you can't send the message to the cell to reproduce. And without (3) there is nothing to send out to the bloodstream to fight the invasion.
How do the B-cells make so many flavours of itself? By a complicated procedure of gene splicing. Basically there are three clusters of genes that are varied. An example taken from Keith Rodison, would make this easier to understand [Something Behe's seems to avoid doing in this section of his book] The genome for antibody schematically looks like this V1-V2-V3-V4-D1-D2-D3-J1-J2-C, with V, D and J being the variable clusters. To make a B-cell, one gene segment is drawn from V, one from D, one from J and a constant one from C. Thus a possible antibody genome would be V2-D1-J2-C and another may be V1-D1-J1-C. With the large number of gene segments in the human genome, these can lead to millions of combinations for the binding site-thus it is virtually certain to bind to any foreign molecule. Here is, according to Behe, another example of irreducible complexity-since a primitive system with only one or a few antibody molecules would be "like a propeller turning at one revolution a day". In other words, without the complicated gene splicing in clonal selection we would not have a functioning immune system at all.[6]
- Metabolic Pathways:
The final example is on metabolic pathways. Behe's example of this is the synthesis of AMP (adenosine monophosphate). Adenine (one of the four bases of DNA [d]) is derived from AMP. Thus to get adenine you must first get AMP. The synthesis of AMP in the cell requires 13 steps utilizing 12 different enzymes. The precursor molecules, the intermediate forms before AMP is reached, play no independent role elsewhere. Their very existence are purely as stepping stones towards AMP (or GMP). This process, according to Behe, is another example of irreducible complexity. Since there is no use of the intermediate molecules, the whole pathway could not have evolved in a Darwinian step by step fashion.[7]
One of Behe's central claims throughout all these examples is that there is nothing in the literature (in scientific journals) which comes close to explaining the possible evolution of these systems. Thus to him, nobody knows how these complex systems come to be.
It is important to note that Behe is not a young earth creationist. He also accepts the fact that all organisms share a common ancestor, i.e. they evolved. This is what he wrote in his book:
| For the record, I have no reason to doubt that the universe is the billions of years old that physicists say it is. Further, I find the idea of common descent (that all organisms share a common ancestor) fairly convincing, and have no particular reason to doubt it.[8] |
However if these irreducibly complex systems did not arrive here by evolution, how did these come to be? After summarily dismissing two other alternatives (Lynn Margulis' "symbiotic" theory of the origin of the eucaryotic [f] cells and Stuart Kauffman's "complexity theory") he concluded that the only option left is intelligent design. However, even to Behe, detecting whether something has been designed or not is not easy:
| The upshot of this conclusion-that anything could have been purposely designed-is that we cannot know that something has not been designed. The scientific problem then becomes, how do we confidently detect design?...For discrete physical systems-if there is not a gradual route to their production-design is evident when a number of separate interacting components are ordered in such a way as to accomplish a function beyond it's individual components. The greater the specificity of the interacting components required to produce the function, the greater is our confidence in the conclusion of design.[9] |
In other words, when a system is irreducibly complex, it is designed.
According to Behe, the designer made all the irreducibly complex systems discussed above and others in the first cell almost four billion years ago[e]:
| Suppose that nearly four billion years ago the designer made the first cell, already containing all the irreducibly complex biochemical systems discussed here and many other. (one can postulate that the designs for systems that were to be used later, such as blood clotting, were present but not "turned on."...) [10] |
The above, in a (rather large) nutshell, is what Irreducible Complexity is all about. Now let us see what is wrong with it.
Back to the top
Slippery Irreducibility
Irreducible complexity is a concept that sounds clear enough, yet upon closer examination, it is hard to be certain if it can be applied in any meaningful way. Remember from Behe's definition above, there are two aspects to this concept; firstly an irreducibly complex system cannot be made any less simpler, i.e. it cannot have less components and still fulfil the same function, secondly, it is impossible to imagine a Darwinian step by step improvement from something simpler. There are problems with both aspects of this definition: it is very hard to be sure systems are irreducibly complex and it is always possible with enough ingenuity to come up with physical precursors for it.Take the mousetrap example again. Behe claims that it is irreducibly complex and needs a minimum of five components (the hold down bar, the hammer, the platform, the spring and the catch), remove any one of these and the system will cease to function. Yet as John McDonald showed [11] the mousetrap can function with 4, 3, 2 or even only one part. Firstly, he removed the catch and bent the hold down bar a little. This hold down bar is placed at the end of the hammer that protrudes out from the spring. The cheese (bait) is attached to the end of this hold down bar. If a mouse tries to get to the cheese, the disturbance will remove the hold down bar from the protrusion of the hammer and, kapaam!!, down comes the hammer on the mouse. Viola, you have a fully functioning mousetrap with only four components! With ingenuity, McDonald was able to reduce the mousetrap to a single spring.
Thus a major example used by the supreme master of recognizing irreducible complexity is conclusively shown to be not irreducibly complex. This serves as a cautionary tale-recognizing systems as irreducibly complex is not easy.
The second facet of his definition, that it is impossible to think of a step by step improvement to this system, had also been shown to be wrong.
Don Stoner, had shown how the Behe mousetrap could have evolved in Darwinian step-by-step fashion from a simple box and stick trap.[12] There are nine steps to this:
- Step One: First we have a box and stick. The stick is proping up the box. If a mouse past through and disturbs the stick, there is a possibility that the box will fall on the mouse trapping it.
- Step Two: Next we have a piece of wood, perhaps from the box itself, that falls on the floor and forms a wooden base. This base, initially small, is still a minor improvement, since it keeps the mouse from burrowing out of that particular part of the box. The bigger the wooden base, the less area the mouse can try to burrow out. This advantage continues until the base covers the bottom of the box.
- Step Three: Now ridges are formed at the edge of the box which touches the base when the trap is armed. This ridge serve to centralise the box, eventually the ridges can start to "squeeze" onto the box on both sides such that an indentation is formed. This becomes the hinge. This makes the box movement more predictable, thus it can catch more mice.
- Step Four: Next a spring is added. This can initially be just a whicker. Even this whicker will help make the box close more quickly. The spring's strength can be increased slowly. Each increase helps the box close faster, making it more efficient at catching mice.
- Step Five: Now the stick can get longer. As it gets longer it holds the box further away from the base, fulfilling two tasks. The box now looks less threatening and should scare away less mice. It now becomes possible to trap the mouse even without the mouse being fully under the box, as the spring force drops at a rate fast enough to alow it to crush the mouse.
- Step Six: With the box now functioning like a hammer, it will catch more mice by crushing it, the top of the box now becomes more of a hindrance as it captures air and slows down the snap. Thus if the box begins to lose the top, it will encounter less air resistance and will snap down even faster, catching even more mice. Eventually the box becomes a rectangular frame. It's now looking more like Behe's "irreducibly complex" mousetrap!
- Step Seven: Like earlier, a hinge can now be formed at the top end of the stick to the frame/hammer. This makes it more stable and less erratic (it is unlikely to come loose and snap suddenly). Again the improvement should help catch more mice.
- Step Eight: Now a bait can be added to the base of the stick, the connection at the bottom of the stick is slowly improved until a trigger is formed. This can be in the shape of a small "mound" on the wooden base which eventually changes into a full fledged trigger. This will improve the trap's effectiveness further.
- Step Nine: At this stage the stick (with the trigger at the bottom) is already able to push the hammer almost all the way down. Now if the hinge is formed at the base (rather than on the hammer), the stick would be able to push the hammer all the way back. This will result in less force on the trigger, making it easier to activate. Now, finally, we have the "irreducible complex" mousetrap formed through a step-by-step improvement in function, the very thing that was supposed to be impossible.
Even with a relatively simple system like the mousetrap, Behe can be shown to be mistaken in the application of his concept. How much confidence can we have when he pronounce biochemical systems "irreducibly complex"? As we shall see below, not much!
Back to the top
Intelligent Design is Untestable
Behe's main thesis of intelligent design is fundamentally untestable. Why? Well because he gave himself a big escape hatch. Look:
| Some features of the cell appear to be the result of natural processes, other probably so. Still other features were almost certainly designed. And with some features, we can be confident that they were designed as that anything was...The production of some biological improvements by mutation and natural selection-by evolution-is quite compatible with intelligent design theory. [13] |
(I have italicised, for emphasis, some portion of the quote above.) Can you see the escape hatch here? If any of his six examples is shown to be derived from earlier precursors, he always have the cop-out- "Yes, I made a mistake here in calling this particular process irreducibly complex, afer all I did say some processes could have evolved. But, hey check out this new one I found." Thus he can always move on to other processes that have yet to be studied deeply and pronounce those "irreducibly complex".
In other words, because there will always be biochemical pathways to study (that's what makes science exciting and vibrant) there will always be frontiers of knowledge where more research is required where some of the evolutionary precursors are unknown, there will always be gaps in our knowledge for the proponents of the ID theory to thrive on. Thus ID and irreducible complexity is just a modern dressed up version of the old god of the gaps argument. Because we do not know what makes the sun shine, God must have done it; because we do not know how human beings arose, God must have created them. But gaps in scientific fields have a habit of closing very fast and (as we shall see below), Behe and his crew will have to find new frontiers of ignorance[g] to slip their Intelligent Designer in.
Back to the top
False Dichotomy
Behe's summary dismissal of other possibilities (apart from Darwinian gradualism and intelligent design) hides the fact that other ideas are making impressive headway. It is certainly possible that the origin of life and/or some new species may have been caused by other laws than Darwinian natural selection. Some examples:
- Complexity Theory: The complexity theories of Stuart Kaufman, Per Bak and Brian Goodwin, apply an interdisciplinary approach (physics, mathematics and biology) to the emergence of order (complexity) from chaotic systems. Far from this being "fact free science" (as Behe claims [14]), David Ussery, an associate professor in the Institute of Biotechnology at the Danish Technical University is currently using these ideas to model protein structures. [15]
- Punctuated Equilibrium: The theory first introduced by Niles Eldridge and Stephen Jay Gould-that evolution happens in short creative burst followed by long periods of stasis. As David Ussery showed, we have evidence of this kind of evolution in the (well documented) evolution of domestic wheat.
- Scaffolding theories: These types of hypothesis are mainly used in origin of life scenarios. A.G Cairns-Smith had suggested that the origins of life may not be carbon based but silicon based.[16] His argument is that the origin need not be made of the same material as the final product. For instance in making the stone arch. Every piece of the arch is in compression and each piece is required to "hold" everything in place. It is impossible to have a step-by-step approach to building this. So how are stone arches built? By scaffolding. The scaffolding is removed after the arch is in place. Note also that the scaffolding material (mainly wood for making the stone arch) need not be the same as the final product. To Cairns-Smith, the crystals of clay, could have played a major role in providing the scaffolding for life to develop.
Of course, non of the above, at the moment, can fully explain the origns of the some of the complex biochemical processes Behe presented. However they still form viable alternatives to Darwinian gradualism and intelligent design. Further, because the intelligent design explanation is possible only after all other possibilities are exhausted-it is, after all, an argument from ignorance- these ideas (including Darwinism) are inherently more plausible.
Thus by presenting Darwinian gradualism and Intelligent Design as an either-or situation, Behe had set up a false dichotomy.[17]
Back to the top
Biochemical Evidence for Darwinian Evolution
Reading Behe's book, one get the distinct impression that biochemistry as a whole is problematic for Darwinian evolution. Thus his statement (see above) about biochemistry being a "Lilliputian challenge" to Darwin and that the complex world revealed by this field "must be explained". Furthermore he mentioned that biochemistry was not "invited" to the meetings that developed the Neo-Darwinian synthesis in the first half of the last century (the discipline wasn't born yet at that time). Thus the complex world that biochemistry discovered was never, and could never be, resolved by Darwinism. [18] This impression, whether it is intended or not, is false.For from disproving Darwinian evolution, biochemistry supplied further evidence of the relatedness of all organism and had even showed the detailed mechanism which allowed for evolution. Some of the evidence from biochemistry include:
- DNA Comparison: We share 98% of the DNA sequence with chimpanzees and 97% with the gorillas. [19]
- Cytochrome-c Comparison: Cytochrome-c is a protein found in all living things and is used in oxidative processes. It has been found that only around 30 or so of the 100 amino acids in the protein is required for it's function. Human cytochrome-c is completely identical with that of the chimpanzee. These two (human and chimpanzee) differs from the cytochrome-c of other animals by at least 10 amino acids. They differ from the yeast Candida krusei by 51 amino acids. In fact the family tree of cytochrome sequence resembles very closely the evolutionary tree suggested by scientists before these sequence were known.[20]
- Pseudogenes: Pseudogenes are DNA sequences that are not functional but are closely related to some other functional genes. Finding the same pseudogene in the same chromosomal location in two different species would give strong evidence of evolutionary descent. There are many examples of such shared pseudogenes in humans and primates.
Biochemistry has also showed how mutations are formed, the various kinds of mutation (e.g. gene duplication, point mutation etc) and how a single point mutation in the DNA sequence could make a different amino acid- hence a slightly different protein.
We end this section with a quote from evolutionary biologist H. Allen Orr in his review of Behe's book:
| The real irony is that...Behe...think[s that] the molecular world poses the most profound challenge to Darwinism. But if one had to point out the one field of evolutionary biology that has been most thoroughly mined, it would likely be molecular evolution. Molecular evolution has dominated-indeed virtually monopolised-the study of evolution for the last forty years. And it has been overwhelmingly successful...Indeed there is a growing sense that many of the deep questions that first tempted evolutionists to think hard about molecules are on the verge of getting wrapped up.[21] |
Back to the top
A Step-by-step Evolutionary Model of a Biochemical Cascade
Behe claims that biochemical cascades, such as blood clotting, could not have been produced in a Darwinian fashion:
| Because of the nature of the cascade, a new protein would immediately have to be regulated. From the beginning, a new step in the cascade would require both a proenzyme at the correct time and place. Since each step necessarily requires several parts, not only is the entire blood-clotting system irreducibly complex, but so is each step of the pathway.[22] |
[Again I have italicised some portions of the above quote for emphasis]
Let us look at the cascade for blood clotting :[23]
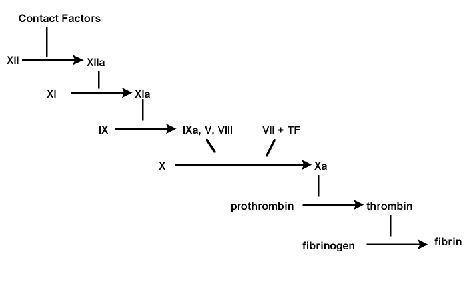 The process of blood clotting consists mainly of the "cleaving" of proteins (or protease reactions [h] ) You will note that each step seems to depend on the previous one. Thus Behe's claim - that these could not evolve step by step. Yet it is relatively easy to construct a model of how a cascade like this could have evolved. I am following the example given by Keith Rodison [24]
The process of blood clotting consists mainly of the "cleaving" of proteins (or protease reactions [h] ) You will note that each step seems to depend on the previous one. Thus Behe's claim - that these could not evolve step by step. Yet it is relatively easy to construct a model of how a cascade like this could have evolved. I am following the example given by Keith Rodison [24]
First let us note a few things. Many of the processes of blood clotting are auto-catalytic; the more the material is made the more it tends to make more of itself. For evolution to happen in a Darwinian fashion, there has to be three factors: a way for the organism to replicate via DNA(and within a population, i.e. we are not talking about a single organism but one that is "typical" of a population), a way for mistakes to enter into the replication (various types of mutation) and a way for advantageous (or at least non-disadvantages mutations to survive in the population) mutations to be selected in a step by step fashion.
Let us see how one step in the cascade may develop:
 | Step One: This is just a simple autocatalytic reaction, where a stimulus reacts with enzyme X changing it into an active state X* which interacts with the target. |
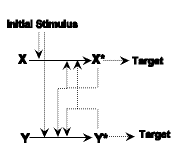 | Step Two: Here there is a mutation called gene duplication. This is a common (and observed) phenomenon where a portion of a gene is copied into another portion of the chromosome. Since both genes are active, they both made the same type of enzyme. I have labelled the "new" duplicate as Y and Y* although they are exactly identical to X and X*. Since these two perform the same function as before there is neither a selective advantage nor disadvantage. Thus we would expect the mutation to linger around in the gene pool, i.e. it would not be immediately eliminated by natural selection. |
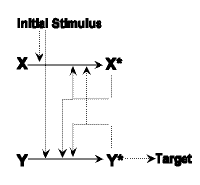 | Step Three: Again mutation happens to some individuals in the population. Now a point mutation happens that affects the ability of the original activated protein, X*, to interact with the target. Point mutations are mutations in one of the DNA base pairs which result in the change of one amino acid in the protein. Again this is a well known and understood phenomenon. However since the process still interact with the target (via Y*) it is not deleterious to the organism and will not be selected against. |
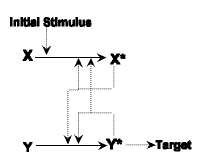 | Step Four: Again mutation happens to some individuals in the population. A point mutation occurs which affects the ability of the the enzyme, Y, to interact with the stimulus. Now Y becomes completely dependent upon the enzyme X* to activate it. A system like this is potentially advantageous since it gives rise ot the possibility of controlling the process and also of amplification. This final cascade is already irreducibly complex, in other words there is no way the target could be interacted with unless it goes through the two stages. Both the Y and the X enzymes now cannot do this alone. |
The model above also makes certain predictions about cascading systems. It says that we would expect to find homologous enzymes in these systems. In other words, since one enzyme (in this case Y) came from gene duplication of X and both subsequently have point mutations we would expect many of their amino acids and their general sequences to be quite similar, or homologous, to each other.
Do we find this in the enzymes involved in the blood clotting cascade? Yes we do! As George Acton noted:
| The five proteases [involved in the blood clotting cascade] are each composed of a single peptide chain which is almost exactly the same length in all, and the point at which the chain is cut in the activation process is in exactly the same relative position in all 5. Eight defined domains or motifs, e.g. the serine protease domain, are located in exactly the same position in 4 of the 5 protease factors. In other words, at this low level of detail all four are 100% homologous. [25] |
Back to the top
Possible Evolutionary Precursors for Behe's Examples
Looking at the six examples Behe presented shows us that not only are there possible pathways for these systems, it throws doubt on how the idea of irreducible complexity can be used in any meaningful way in biochemistry:
-
The bacterial flagellum:
Ian Musgrave has suggested a very interesting evolutionary pathway for the bacterial flagellum. [26] First he casually points out that the major components of the flagellum, the filament, the hook, the rod and the motor are hollow. He then noted that there are major structural similarities between the major components of the bacterial flagellum and a type of bacterial protein secretion system called the type III secretory systems. The type III secretory system has a rivet identical to the rod of the flagellum. The switching/torque generation system also has homologs in type III secretary systems. Some type III secretory system even have tubular structures attached to the rod which resembles the filament of the flagellar system! In type III secretary system proteins are exported through the hollow motor ring and rod! Homology studies have also shown the proteins of the type III secretary system and the flagella are related, in fact in some bacteria these proteins are identical to each other!Given these clues, he then suggested how the flagellum could have evolved. First the SMC ("Motor") rod pore is evolved forming the simplest secretory system. Then the proto-flagellum arose as part of the protein secretion structure. Finally an ion pump which initially was used for something else (such as keeping ionic cell balance) was incorporated into this structure which gives it perhaps some weak motility. This then is a case where a structure originally meant for some other purpose (protein secretion) evolved to a point where it could be used for another entirely different purpose (motility).
Now see how ludicrous the application of the concept of irreducible complexity is. Viewed as a flagellum, yes, you can't get any simpler than the motor, rod, hook and filament. Yet viewed as a secretory system most of the parts can be dispensed away with. [Update-April 2004: A good recent article on the bacterial flagellum is The Flagellum Unspun: The Collapse of "Irreducible Complexity" by biologist Kenneth Miller.]
- The Cilium
The major components of the cilium all have functions within other parts of the cell which could have been adapted for it's use later. Microtubules have various uses within the cell, including chromosome movements during mitosis. There are also similar versions of the "motor" dynein within the cell performing oher functions. Thus the components of the cilia were not created ex-nihilo purely for providing movement. Clare Stevens, noting that cilia have varying lengths, had suggested that they were initially used to help the cell lie on a surface film and later co-opted for motility. [27] - Blood Clotting
We have already treated blood clotting in some detail above. We just add a note here that recent studies have shown that some of the processes in blood clotting (such as Factor XII-which was presented in Behe by it's complex earlier names-Hegeman, prekallikrein and HMK) are now excluded from the basic process since their deficiences do not lead to any malfunction of the system. [28] Thus apart the possibility of step-by-step evolution of the blood-clotting cascade, we see that, the process, as it is presented by Behe is not even irreducibly complex, since Factor XII can be done away with. - Cellular Transport Systems
Here we have a truly complicated system. And here too we see flaws in Behe's presentation. The system, protein transport, is not irreducibly complex, since, as Dave Ussery pointed out, some studies have shown that some proteins direct their own secretions, i.e they do not use any transport proteins. These systems could easily have been the precursor to the vesicular transport system. Similarly, there are bacterial transport systems that are much simpler than the ones Behe pointed to in his book which could easily have been the precursor.[29] - The Immune System
Behe's example (given above) is far from irreducibly complex. As Keith Rodison noted:[S]ome moths produce a protein call hemolin, which binds (in an apparently generic manner) to bacteria and assists in removing the bacteria from the hemolyphs (the blood-like substance in insects). The protein sequence of hemolin reveals it to be a relative of vertebrate antibodies.[30] Thus in one fell swoop we have a precursor to the mammalian antibody which works in only one step. This without needing any of the complicated gene combination that was explained above
Again we see in nature a gradation of the antibody diversity system. Studies of primitive jawless fish shows that there exist gradiations of the antibody diversity generating systems. The hagfish was tested and showed no adaptive immune response when a foreign substance was injected into it.[i] However when the same test is done on the lamprey (another jawless fish) it exhibited "a feeble, but nonetheless definite, adaptive immune response to certain antigens." When the same test is done on a "higher" fish-such as the shark-it's shows a higher concentration of specific antibody production than the lamprey. [31]
In fact the antibody genome of sharks have been studied. Instead of the complicated gene splicing found in human antibody generation, the shark has various genomes already assembled, e.g. the genome would look something like this, V5-D5-J5-C; V6-D6-J6-C and so on, in long strings of DNA. This is a simpler system which functions well enough for the shark. Thus even here the gene-splicing mechanism is not required for the antibody diversity. [32] So much for irreducible complexity in the immune system!
- Metabolic Pathways
There are literally thousands of biochemical metabolic pathways. Yet Behe had purposely chosen an example whose evolutionary origins is not that well studied, this ostensibly to bolster his claim that "There has never been a meeting, or a book, or a paper on details of the evolution of complex biochemical systems." [33]Yet many other metabolic pathways have been studied in detail. One such cycle is the well-known Kreb-cycle (also known as citric acid cycle or tricarboxylic cycle-depending on who you want to impress). The Kreb-cycle is the second stage of the metabolism of glucose and synthesizes ATP. It requires nine different enzymes to work, remove anyone and the system cease to function. Irreducibly complex, right? Wrong! As Keith Rodison pointed out, citing a paper in the Journal of Molecular Evolution published about the same time as Behe's book, the evolution of the complex biochemical pathway in a step-by-step manner is being explored. The evolution of other biochemical pathways, such as pentose phosphate pathway, glycogen biosynthesis and amino acid synthesis, are all being studied and papers are published. [34]
Furthermore recently [j] a novel metabolic pathway, that degrades the toxic chemical pentachlorophenol (PCP), has been discovered in a bacteria called Sphingomonas chlorophenolica. This pathway provides the only source of carbon for the bacteria. This metabolic degradation requires three enzymes. The removal of any one of these enzymes will kill the bacteria, as it no longer has any carbon source. This is thus an irreducibly complex process. However PCP is a man-made chemical that is not known to exist naturally and, get this, PCP was first introduced into the environment only in 1936. Thus the bacteria must have evolved this irreducibly complex metabolic pathway less than 65 years ago! Shelley D. Copley [35] had suggested that this pathway evolved through the co-option of enzymes already used in other pre-existing pathways.
Thus Behe's claim, that no one has studied the evolution of these pathways, is shown to be false.
Behe's Super Bacteria
As you can see above Behe's "speculative" scenario for the origins of genes for all the irreducibly complex systems we see today is that the designer makes the first cell already containing the genes of irreducibly complex systems Thus the blood-clotting gene is already present in this first "SuperAdamite Bacteria" but was not "turned on". Let us see this quote from Behe again:
| Suppose that nearly four billion years ago the designer made the first cell, already containing all the irreducibly complex biochemical systems discussed here and many other. (one can postulate that the designs for systems that were to be used later, such as blood clotting, were present but not "turned on."...) [10] |
I let Behe refute his own scenario:
| [I]f a protein appeared in one step with nothing to do, then mutation and natural selection would tend to eliminate it. Since it is doing nothing critical, it's loss would not be detrimental, and production of the gene and protein would cost energy that other animals aren't spending.[36] |
Delicious! Behe is saying here that genes that are not used tend to be eliminated over time. Yet his "speculative scenario" postulates just such genes! Blood clotting would not be required for billions of years (for added affect say this in a Carl Saganish baritone!) What keeps the genes from being mutated beyond recognition and unusable? Could it be that he would postulate the designer would somehow prevents its loss? Perhaps with the "shield of righteousness"?
There are other problems with this scenario, of course. As H. Allen Orr asks:
| Although some genes do get killed and silenced over time (producing non-functional "pseudogenes"), how come we only carry pseudogenes that are wrecked copies of our real genes? In other words, why don't I carry pseudogenes for chlorophyll or flower structure? Why don't azaleas carry pseudogenes for brain cells?[37] |
I think there is a reason why Behe called his suggestion "speculative", he knows it is very weak. Yet, if these irreducibly complex systems did not evolve and, as we have seen, did not appear with the first bacteria, how did they arise? The only other alternative I can think of for his postulate of intelligent design is for the designer to periodically interfere in the evolution of living things (say whenever a major transition were to take place) by inserting the newly required irreducibly complex genes into their genomes! This periodic intelligent design means that an ad hoc miraculous intervention is required every few million years.
Back to the top
Conclusions on Irreducible Complexity and Intelligent Design
Our conclusions are many:
- Irreducible complexity may not be a meaningless term, certainly some structure (both macro and micro structures) are irreducibly complex in the sense that there may be no simpler way to fulfil the current function. Yes as we have seen above it is not a useful tool in biochemistry or in the study of evolution. Firstly it is hard to be completely sure if a structure is irreducibly complex. We have seen examples of these in the mousetrap and in some of Behe's biochemical examples where structures he thought were irreducibly complex turn out to be not so.
- Behe's idea is not refutable in principle, since he admits that some systems may have evolved. Thus upon being shown one biochemical example is not irreducibly complex, he would simply shrugged his shoulders admit that it is part of an evolved system and go on his merry way pointing out other irreducibly complex system.
- Like the creationists, Behe presents the case between Darwinian gradualism and Intelligent design as though these are the only two alternatives available. And refuting one automatically proves the other. This, as we have seen above, is a false dichotomy.
- Biochemistry, far from being a "challenge" to Darwinism, had actually strengthened it by providing even more proofs of evolutionary relatedness and by providing concrete explanations of how mutations can occur.
- It is possible for irreducibly complex biochemical cascades to evolve in a step-by-step Darwinian fashion.
- His six examples are already being shown to be shaky and eventually all these gaps in scientific knowledge will close.
- Perhaps the main weakness of his postulate is in postulating just exactly how non-evolving irreducibly complex genes get embedded into the genome. Behe demands that evolutionsts explain how irreducibly complex biochemical systems evolve step by step, yet he can't even begin to come up with a believable scenario for this intelligent design idea.
To all these I will add one more objection: despite what he claimed in his book [38], his Intelligent Design is still subject to the same critique Hume made more than two hundred years ago.
In short, irreducible complexity and intelligent design, fail to provide any evidence for a designer.
Back to the top
Notes
| a. | Cytoplasm is the base material of the cell. This is kinda like your face is the base which includes parts such as your nose, your eyes etc. Anything outside the cell nucleus is in the cytoplasm. |
| b. | The lysosome is the garbage collector/scavenger in the cell. |
| c. | mRNA stands for messenger RNA. RNA, or ribonucleic acid, is the information required to build the protein. mRNA is copied from the DNA (deoxyribonucleic acid)which is the main "blueprint" of the cell. |
| d. | The other three bases are thymine (T), guanine (G) and cytosine (C). These four bases, A, T, G and C are all that is required to code for all the proteins living things require. There is another base, uracil (U) which is found only in RNA (taking the place of thymine). |
| e. | This point will be crucial later on. Although Behe's calls his suggestion "speculative" (i.e. "don't associate my ideas too close with this!"), there is actually little else that can work in this scheme of things. The only other alternative is for the "designer" (let's call a spade a spade-Behe's God) to periodically intervene in evolution and add irreducibly complex systems from time to time into living genomes. |
| f. | Eucaryotic cells are cells with a nucleus, the cells of all animals and plants are of this type. Procaryotic cells belongs to organisms such as bacteria. These types of cells have no nucleus, no internal membranes and non of the sophisticated organelles (such as mitochondria) we find in eucaryotic cells. |
| g. | Keith Robison's phrase, describing Behe's book. |
| h. | Proteases are a class of enzymes that can cut peptide bonds between amino acids in a protein. Think of a biochemical "chopper". |
| i. | This does not mean that the hagfish has no immunity, just that it did not react to the antigen (the injected substance) in a specific way. |
| j. | Thanks to Douglas Theobald, from the Department of Chemistry and Biochemistry in the University of Colorado at Boulder, for pointing this out to me and for providing me with a copy of the paper from TIBS! |
Back to the top
References
Back to the top