INTRODUZIONE
 Distribuiti in gran parte della regione Olartica, i Branchiobdellidi sono un gruppo abbastanza uniforme di circa 150 specie. Vivono come ectosimbionti sul corpo di crostacei d'acqua dolce, in particolare gamberi, ma anche granchi e isopodi.
Gli adulti sono lunghi al massimo 10 mm, mentre le specie pi� piccole possono avere dimensioni inferiori al millimetro. La forma � a bacchetta, a pera o a fiasco, pi� o meno allungata. Il corpo pu� essere appiattito ventralmente o dorsoventralmente.
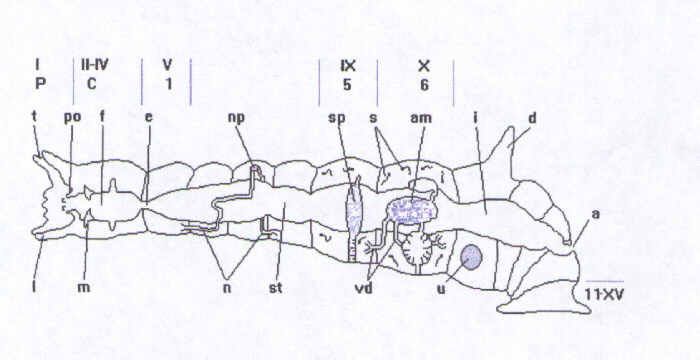
Il numero di segmenti � costante: i primi quattro costituiscono il capo, con un organo adesivo orale formato dal primo segmento (peristomio); il corpo � costituito da undici segmenti di cui l'ultimo o gli ultimi due, formano l'organo adesivo posteriore. Ogni segmento del corpo risulta essere diviso da un solco in un anello anteriore ed uno posteriore pi� piccolo. Sull'anello pi� grande, in alcune specie, si sviluppano proiezioni dorsali digitiformi o espanse. Il peristomio � costituito da due labbra, una dorsale e una ventrale. Quella superiore pu� essere dotata di lobi o tentacoli.
La bocca � normalmente circondata da papille. Il canale alimentare inizia con la faringe muscolosa nella cui parte anteriore sono situate due mascelle, una superiore e una inferiore, formate dall'ispessimento di pieghe della cuticola faringea. L'esofago � un breve tubo cui segue lo stomaco parzialmente diviso in pi� camere da setti. Una costrizione separa lo stomaco dall'intestino anteriore, anch'esso suddiviso in camere. Un altro setto segna il passaggio all'intestino posteriore che si restringe a formare il retto. L'ano si apre nel decimo segmento in posizione dorsale mediana.
Distribuiti in gran parte della regione Olartica, i Branchiobdellidi sono un gruppo abbastanza uniforme di circa 150 specie. Vivono come ectosimbionti sul corpo di crostacei d'acqua dolce, in particolare gamberi, ma anche granchi e isopodi.
Gli adulti sono lunghi al massimo 10 mm, mentre le specie pi� piccole possono avere dimensioni inferiori al millimetro. La forma � a bacchetta, a pera o a fiasco, pi� o meno allungata. Il corpo pu� essere appiattito ventralmente o dorsoventralmente.
Il numero di segmenti � costante: i primi quattro costituiscono il capo, con un organo adesivo orale formato dal primo segmento (peristomio); il corpo � costituito da undici segmenti di cui l'ultimo o gli ultimi due, formano l'organo adesivo posteriore. Ogni segmento del corpo risulta essere diviso da un solco in un anello anteriore ed uno posteriore pi� piccolo. Sull'anello pi� grande, in alcune specie, si sviluppano proiezioni dorsali digitiformi o espanse. Il peristomio � costituito da due labbra, una dorsale e una ventrale. Quella superiore pu� essere dotata di lobi o tentacoli.
La bocca � normalmente circondata da papille. Il canale alimentare inizia con la faringe muscolosa nella cui parte anteriore sono situate due mascelle, una superiore e una inferiore, formate dall'ispessimento di pieghe della cuticola faringea. L'esofago � un breve tubo cui segue lo stomaco parzialmente diviso in pi� camere da setti. Una costrizione separa lo stomaco dall'intestino anteriore, anch'esso suddiviso in camere. Un altro setto segna il passaggio all'intestino posteriore che si restringe a formare il retto. L'ano si apre nel decimo segmento in posizione dorsale mediana.
L'apparato riproduttore � costituito da due paia di testicoli, situati nei segmenti 5 e 6, e da due ovari nel segmento 7. Gli spermatozoi, liberati nel celoma, sono raccolti da un paio di imbuti ciliati, che si aprono nella cavit� di ciascun segmento contenente i testicoli, comunicanti con due dotti per ciascun segmento i quali si uniscono a formare i vasi deferenti e confluiscono nel dotto spermatico ghiandolare (segmento 6). Da questa ghiandola prende origine un breve atrio muscolare (dotto eiaculatore) che termina col pene, il quale pu� essere eversibile o protrusibile.
Il sistema vascolare consiste di due vasi principali, uno dorsale contrattile e uno ventrale (posto tra intestino e cordone nervoso), connessi da quattro paia di vasi laterali nel capo e da un singolo paio in ciascuno dei segmenti 1, 7 e 11 del corpo.
Il sistema nervoso � costituito da un cervello posto dorsalmente unito a due cordoni nervosi ventrali per mezzo di connettivi circumesofagei. Il capo contiene quattro paia di gangli, ciascun segmento del corpo ne contiene un paio, mentre ve ne sono tre paia nel segmento 11. I gangli di ciascun segmento sono collegati da commessure ed uniti a quelli degli altri segmenti da connettivi.
Due nefridi, situati asimmetricamente nella parte anteriore del corpo, si aprono per mezzo di un poro dorsale nel terzo segmento. Posteriormente, nel segmento 8 si trovano altri due nefridi che sboccano laterodorsalmente nel segmento 9.
La muscolatura della parete corporea consiste di uno strato circolare esterno ed uno longitudinale interno. Le fibre dello strato circolare si trovano immediatamente sotto l'epitelio. Le fibre longitudinali, situate sotto quelle circolari, decorrono da un setto all'altro.
La posizione sistematica dei Branchiobdellidi ha costituito un problema per i tassonomi. A lungo considerati delle sanguisughe, sono stati anche collocati nella classe degli Oligocheti. Nel 1965 Holt propose di fare dei Branchiobdellidi un ordine dei Clitellati. Malgrado la quasi unanimit� rispetto a questa classificazione, vi � ancora chi considera i Branchiobdellidi una sottoclasse di Irudinei (Sawyer, 1986) o di Oligocheti (Timm, 1991). Secondo Gelder e Brinkhurst (1989) ci� che permette di associare i tre taxa � la comune presenza di clitello, mentre pochissimi dati supporterebbero la tesi di Sawyer. I caratteri comuni tra Irudinei e Branchiobdellidi (numero costante di segmenti, assenza di peristomio, assenza di setole, riduzione della cavit� celomatica, ventose, faringe muscolare, mascelle) sarebbero dovuti a fenomeni di convergenza evolutiva (Brinkhurst & Gelder, 1989). Confrontando questi caratteri si pu� per� osservare che sono pi� le divergenze che le similarit� tra Irudinei e Branchiobdellidi. Il numero di segmenti � di 34 nei primi e solo di 15 nei secondi. La riduzione della cavit� celomatica � parziale nei Branchiobdellidi, mentre � notevole nelle sanguisughe, e le mascelle sono rispettivamente 2 e 3, tra l'altro posizionate in maniera differente. Inoltre, mentre nelle sanguisughe le ventose sono strutture muscolari, nei Branchiobdellidi sono presenti solo dischi di attacco muniti di ghiandole che producono secrezioni adesive; la muscolatura della ventosa posteriore servirebbe solo per la locomozione, ma non per aderire all'ospite. Lo studio comparato dell'embriogenesi di diversi anellidi conferma la teoria di Holt in quanto, accanto ad alcune caratteristiche comuni alle altre classi, i Branchiobdellidi posseggono caratteri peculiari, quali le uova con minor quantit� di tuorlo. Alcune caratteristiche, come le modalit� di segmentazione simili a quelle osservabili in Tubifex, avvicinano i Branchiobdellidi agli Oligocheti, ma altre, come la gastrulazione simile a Piscicola, li accomunano agli Irudinei (Anderson, 1973). Anche la filogenesi del gruppo pone alcuni problemi. Holt (1968 a), partendo da un ipotetico protobranchiobdellide con caratteristiche primitive (due nefridiopori anteriori, due paia di testicoli, spermateca bifida, pene eversibile), ha ricostruito l'albero filogenetico dei Branchiobdellidi viventi, basandosi sulle caratteristiche dell'apparato riproduttore e sulla loro distribuzione.
L'Europa � abitata da specie appartenenti al genere Branchiobdella con caratteri primitivi ed alcune spiccate divergenze. Specie di Branchiobdella sono presenti anche in Estremo Oriente (il genere avrebbe raggiunto l'Europa partendo dall'Asia) dove domina il genere Cirrodrilus che appare strettamente correlato all'americano Ankyrodrilus.
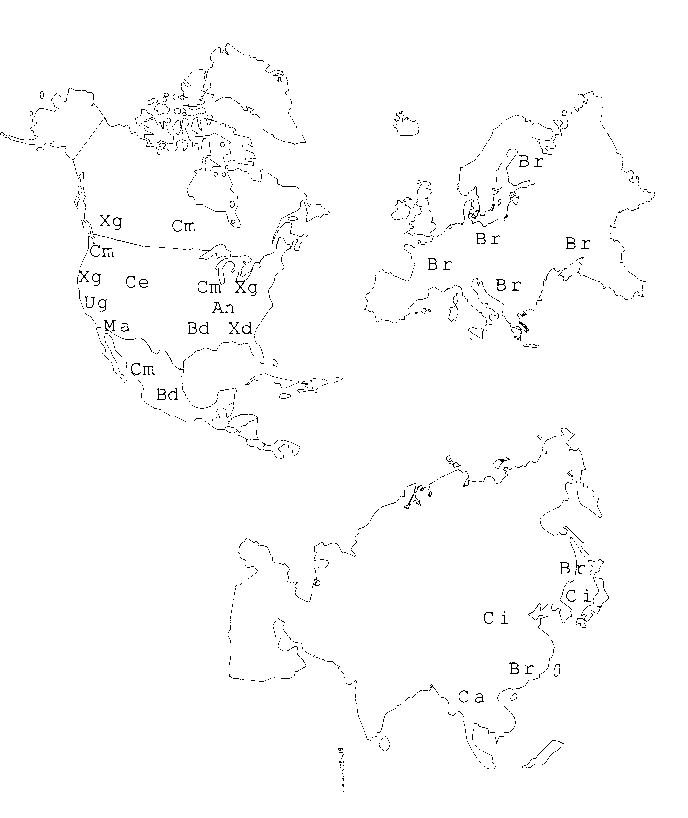
Fig.1- Distribuzione di alcuni generi di Branchiobdellidi in Nord America, Europa ed Asia: An: Ankyrodrilus; Bd: Bdellodrilus; Br: Branchiobdella; Ca: Caridinophila; Ce: Ceratodrilus; Ci: Cirrodrilus; Cm Cambarincola; Ma: Magmatodrilus; Ug: Uglukodrilus; Xd: Xironodrilus; Xg: Xironogiton.
Meno chiare sono le relazioni tra i generi viventi nel continente Nord Americano ( Holt, 1968 a; B�n�rescu, 1990). In Australia, la nicchia ecologica dei Branchiobdellidi � parzialmente occupata da Temnocefalidi (Platelminti: Turbellari) presenti anche in Sud America dove la presenza dei Branchiobdellidi non � ancora ben definita (Holdich & Lowery, 1988).
Vista la continua scoperta di nuove specie (pi� di ottanta dal 1950) Holt (1986) ritiene pi� che giustificata la separazione dei Branchiobdellidi sia dagli Irudinei che dagli Oligocheti. Holt considera anche la relazione esistente tra i Branchiobdellidi ed i loro ospiti. Non ritiene possibile una comune evoluzione dei vermi e dei crostacei, ma piuttosto suppone che i Branchiobdellidi inizialmente vivessero simbionti di animali d'acqua dolce per poi, in un secondo tempo, "colonizzare" gli ospiti attuali. Infatti i Branchiobdellidi devono essersi originati in acque dolci in quanto non sono in grado di tollerare una elevata salinit�, mentre i loro principali ospiti, gli Astacoidei, hanno avuto sicuramente un'origine marina. Il numero di Branchiobdellidi presente su di un unico ospite pu� essere molto elevato. Da studi compiuti su gamberi nord americani ed europei (Hobbs et al., 1967; Vey, 1981) risulta che questi crostacei possono trasportare sul proprio esoscheletro centinaia di Branchiobdellidi. Hobbs et al. sottolineano poi il fatto che non esiste specificit� di associazione tra Branchiobdellidi ed i loro ospiti, anche se i primi devono necessariamente vivere sul corpo dei crostacei, dai quali dipendono per la propria sopravvivenza. In particolare alcune ricerche (Young, 1965) sembrano dimostrare la necessit� dei Branchiobdellidi di deporre i propri cocoons sui crostacei, tant'� che le uova rimosse dall'ospite o lasciate sull'exuvia dopo la muta degenerano in poche ore. La mancanza di specificit� fa s� che si possano trovare specie differenti di questi vermi su un unico gambero, ciascuna per� occupa un microhabitat particolare, per esempio Ankyrodrilus koronaeus si trova esclusivamente sui chelipedi, Pterodrilus alcicornus � distribuito sulla superficie ventrale del gambero, Cambarincola fallax occupa i segmenti basali di alcune appendici quali le antenne, mentre Bdellodrilus illuminatus vive sulle branchie dell'ospite.
Le notizie riguardanti le modalit� con cui i Branchiobdellidi infestano i gamberi sono piuttosto scarse: probabilmente � sufficiente il contatto con un animale gi� infestato. Una volta occupato l'ospite i vermi non vengono mai persi neppure durante la muta quando il gambero abbandona il vecchio esoscheletro. Nella maggior parte dei casi i Branchiobdellidi non sono parassiti, ma sono da considerare come epibionti. L'analisi del loro contenuto intestinale dimostra che la quasi totalit� delle specie di Branchiobdellidi si nutre di ci� che trova sull'esoscheletro del proprio ospite: alghe, protozoi, larve di insetti ed eventuali detriti. Soprattutto tra le specie appartenenti al genere Cambarincola, pare sia diffuso il cannibalismo (Hobbs et al., 1967; Holt, 1968 a). I Branchiobdellidi possono anche nutrirsi delle uova dell' ospite, ma pare che si limitino a quelle che comunque non giungerebbero alla schiusa (Vey, 1981). L' elevato numero di Branchiobdellidi presenti su di un unico ospite pu� in alcuni casi debilitare il crostaceo. Questo succede soprattutto se fra le specie di Branchiobdellidi vi sono quelle che si nutrono dei filamenti branchiali, provocando perdite di sangue continue. L'eventuale morte � comunque provocata solo indirettamente dai vermi, che con la loro attivit� possono aprire la via ad agenti patogeni quali batteri o protozoi (Holdich & Lowery, 1988; Vey, 1981). Numerose sono le prove che i Branchiobdellidi possano sopravvivere senza ospite, senza apparenti effetti negativi (Penn, 1959). Studi pi� recenti (Young, 1965) confermano questi dati. Numerosi esemplari stabulati di Cambarincola vitrea sarebbero sopravvissuti nutrendosi di alghe e altri microorganismi presenti nell'acqua.
Negli ultimi anni, per contribuire a far chiarezza sulla sistematica dei Branchiobdellidi, oltre ai tradizionali caratteri morfologici, alcuni autori hanno preso in considerazione le caratteristiche degli spermatozoi (Ferraguti & Gelder, 1990), della parete muscolare del corpo e dell' ultrastruttura delle fibre muscolari (Valvassori et al., 1994; de Eguileor & Ferraguti, 1980). La struttura degli spermatozoi dei Branchiobdellidi appare simile a quella degli Oligocheti per ci� che riguarda il nucleo e la parte intermedia, ma � pi� vicina al modello degli Irudinei se si considerano il flagello ed il complesso acrosomale (Ferraguti & Lanzavecchia, 1977). Questi caratteri con alcuni autapomorfismi tipici del taxon, quali la riduzione delle fibre marginali e la concavit� della sommit� del nucleo, possono fornire preziose indicazioni per stabilire la filogenesi del gruppo (Ferraguti et al., 1986; Ferraguti & Gelder, 1990). Anche la morfologia delle fibre muscolari varia nelle diverse specie di Branchiobdellidi. L' analisi comparata della muscolatura della parete del corpo di alcune specie appartenenti a famiglie diverse mette in risalto come lo stile di vita possa avere influenzato lo sviluppo di questi tessuti. Specie con vita sedentaria, come Bdellodrilus illuminatus , mostrano una riduzione delle fibre muscolari ed un aumento degli spazi intermuscolari rispetto a specie pi� attive quali Xironogiton instabilis, Cambarincola fallax, Ankyrodrilus legaeus o Branchiobdella kozarovi (Valvassori et al., 1994). Anche la presenza di un sistema T nelle fibre circomiarie cilindriche di Branchiobdella pentodonta sembra avere un significato adattativo. Permetterebbe una migliore contrazione della fibra e quindi assicurerebbe all'animale la possibilit� di movimenti rapidi e scattanti (de Eguileor & Ferraguti, 1980). Le caratteristiche delle fibre muscolari hanno permesso di confermare la classificazione gi� proposta da Gelder per i Branchiobdellidi appartenenti ad alcune specie. La muscolatura longitudinale � costituita solo da fibre circomiarie cilindriche in Ankyrodrilus legaeus, Branchiobdella kozarovi e Xironogiton instabilis, appartenenti alla famiglia Branchiobdellidae, mentre i generi Bdellodrilus illuminatus e Cambarincola fallax, appartenenti alle famiglie Bdellodrilidae e Cambarincolidae, presentano, accanto a poche fibre circomiarie cilindriche, un consistente numero di fibre "poliplatimiarie". In questo studio abbiamo preso in considerazione la morfologia e l'ultrastruttura delle fibre della parete muscolare di altri dieci branchiobdellidi, appartenenti ai taxa Branchiobdellida e Bdellodrilida, allo scopo di ampliare le scarse conoscenze di questo gruppo e di correlare i diversi adattamenti della muscolatura della parete corporea con il diverso stile di vita e la diversa posizione sistematica.
BDELLODRILIDAE
CAMBARINCOLIDAE
BRANCHIOBDELLIDAE
XIRONODRILIDAE
CONCLUSIONI