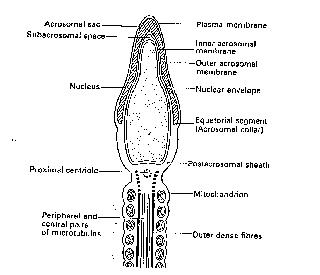
Figure 1. Outline of a spermatozoon.
Conception is the consequence of a series of complex events which include final maturation of the spermatozoon and oocyte, transport of the gametes in the female genital tract (sperm must travel from the vagina, through the cervix and up the Fallopian tube, until they reach the ampulla where the ovulated ovum should be waiting) and the establishment of a diploid (2n) number of chromosomes (2n=46 chromosomes) referred to as syngamy (the coming together of chromosomes, one set each of 23 chromosomes from the egg and from the spermatozoon).
- All mature eggs (ova) contain 23 chromosomes.
- Mature sperm also contain 23 chromosomes.
- One of the 23 chromosomes in all ova is a sex chromosome (ova only carry X chromosomes).
- Sperm also carry a sex chromosome. However, unlike the female ovum, sperm contain either an X or a Y chromosome!
- When a sperm fertilises an egg, the resulting embryo contains 46 chromosomes, of which 2 will be sex chromosomes. The sex combinations will therefore be either XX (female) or XY (male).
- Therefore it is the sperm which determines whether the sex of a child will be boy or girl.
Sex of an embryo
is determined by sperm.
The mature sperm containing a haploid (n=23 chromosomes) number of chromosomes (n=22 + a male chromosome-Y or a female chromosome-X) is a few microns long only, but considering its relatively small size, must travel an epic distance of 30-40cm in the female genital tract to fertilise the oocyte. Sperm have a complex structure which consists mainly of the nucleus (genetic package consisting mainly of chromosomes). The nuclear portion of the sperm is capped by the acrosome (an enzyme containing portion of the sperm head which is vital for the penetration of the outer membrane of the oocyte-the zona pellucida), a midpiece (the energy source) and a tail (provides motility and propulsion) (see Figure 1).
Figure 1. Outline of a spermatozoon.
- In humans, most cells contain 46 chromosomes. These cells are diploid in state.
- Meiosis produces mature egg and sperm, which contain just 23 chromosomes, which are described as haploid.
- A diploid primary spermatocyte , which is an immature sperm, undergoes meiosis, producing 4 haploid mature sperm.
- A diploid primary oocyte, an immature egg, undergoes meiosis, producing only one haploid mature ovum (containing one polar body).
During the process of production, sperm pass through the seminiferous tubules to the rete testis, onto the vasa efferentia, the head of the epididymis and thence 12 days later to the tail of the epididymis. Transport of the mature sperm is via muscular activity within the epididymis and vas. During this time, the sperm acquires motility and undergoes the final biochemical changes that confer them with the ability to fertilise the ovum following ejaculation. Seminal fluid coagulates at ejaculation (it is made up from the secretions of a number of glands, such as the bulbo-urethral, seminal vesicles and the prostate, all of which add to the half a millilitre or so of epididymal fluid), but under normal circumstances, then liquefies within 20 minutes, its basic pH protecting the sperm in the acidic vagina. Sometimes, when certain enzymes are missing the coagulation persists and can bring about infertility by restricting the sperm from gaining access to the cervix). Within minutes after ejaculation, sperm may be found in the cervix (see figure 2) and are released constantly over a period of up to 72 hours, during which time the sperm will move with great speed and direction to the ampulla
where fertilision of the ovum (mature ovulated oocyte) will occur. To achieve this, the sperm must undergo capacitation (attaining fertilising capability; oestrogen dependent) and activation (readiness to undergo acrosome reaction, a change in movement pattern also occurs, inner membrane underneath acrosome cap becomes primed for fusion with the inner membrane of the ovum-oolemma; calcium dependent) (Figure 3).

Figure 2. Sperm moving through
cervical mucus
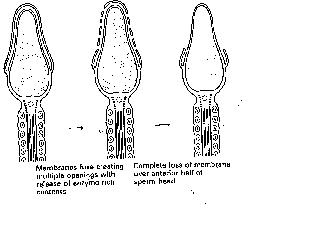
Figure 3. The
acrosome reaction exposes the inner membrane
of sperm. This portion will fuse
with the oolemma of the ovum.
Sperm are produced at the onset of puberty in boys. Thereafter the seminiferous tubules of the testis will produce sperm on a daily basis until well into an average person`s late middle age (up to 60).Meiosis will be occuring at all hours of the day, during which time one primary spermatocyte will produce four spermatids (which will subsequently become four mature sperm). In the ovary, primordial follicles, comprised of primordial germ cell (immature diploid egg-oogonia) surrounded by mesenchymal cells (cells derived from endodermal tissue, which have the ability to migrate) will all have arrived at and arrested (stopped) at the first stage of meiotic division at birth and thereby become primary oocytes. This process being in stark contrast to that in the testis, in which sperm are made continuously throughout adult life. This arrested state (at Prophase I of the meiotic cycle) is maintained until puberty when a few follicles at a time recommence growth on a daily basis, the largest being 2-5mm in the late luteal phase. It is from these that the follicle destined for the next ovulation will be selected (see Figure 4), but it is important to note that in each cycle only one egg is usually ovulated. Antral follicles are surrounded by granulosa (inner) and theca (outer) cell layers (derived from mesenchyme), cells which synthesise androgens (theca cells) and oestrogens (granulosa cells) (Figure 5).
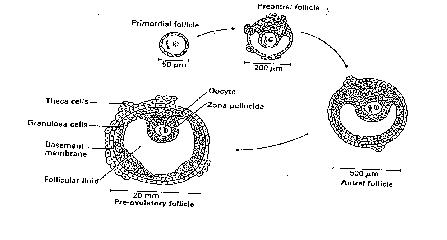
Figure 4. Development of
a follicle in a menstrual cycle.
Meiosis is not resumed until
after the LH surge.
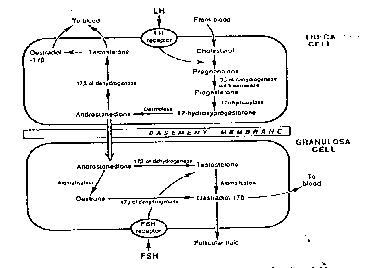
Figure 5. The biochemistry of hormone production in the ovary.
Complex mechanisms ensure that only one follicle (the so-called dominant follicle) becomes preovulatory (the other follicles undergoing atresia within the first week of the follicular phase (growth period prior to ovulation). Other biochemical interactions occur within the developing follicle between the oocyte and granulosa and thecal cells as seen in Figure 5, culminating in high oestrogen levels bringing about the preovulatory surge (the LH surge), which then initiates the resumption of meiosis, ending with the extrusion of the first polar body whence the oocyte, now described as an ovum has acquired the capability to be fertilised (Metaphase II of the meiotic cycle, see figure 6).

Figure 6. Mature ovum (Metaphase
II). The extruded
polar body may be seen at 8 o clock.
At ovulation, the ovum covered in cumulus cells (jelly like protective coating surrounding the egg, see figure 7) is picked up by the fimbria of the Fallopian tube and then swept by ciliary action towards the ampulla where fertilisation occurs.

Figure 7. Human ovum surrounded by cumulus (jelly).
The sperm penetrates the cumulus, corona radiata (a ring of cells surrounding the egg-like a halo) and zona pellucida (outer coating around the egg). Binding to the zona pellucida leads to the sperm undergoing the acrosome reaction, after which penetration and fusion with the oolemma (membrane of the egg) will occur, leading to completion of the fertilisation process. Fertilisation is completed within 20 hours resulting in a return to a diploid (i.e. 2n, a cell containing 46 chromosomes) genetic constitution of the embryo as evidenced by the appearance of two pronuclei at this time in the centre of the fertilised egg (see figure 8).
Figure 8. Fertilised egg showing
two pronuclei.
One pronucleus is derived from the egg (containing 23 chromosomes). The other is derived from the sperm (containing 23 chromosomes). This fertilised egg is diploid containing 46 chromosomes in total.The coming together of the two haploid sets of chromosomes called syngamy, is the final phase of fertilisation. Soon after, anaphase and telophase are completed, the one cell zygote becomes a two cell embryo (see figure 9).

Figure 9. A two cell embryo is
formed after syngamy.
IMPLANTATION
A well developed five day old embryo will have achieved the blastocyst stage (see figure 10). For a pregnancy to establish, the embryo must hatch (escape from the zona pellucida, the outer covering) and then begin to burrow into the womb lining (the endometrium) by sending out invading tissue (implantation). To prepare for this, the endometrium undergoes complex cyclical changes, in particular, extensive proliferation occurs under the influence of oestrogen during the follicular phase (prior to ovulation).
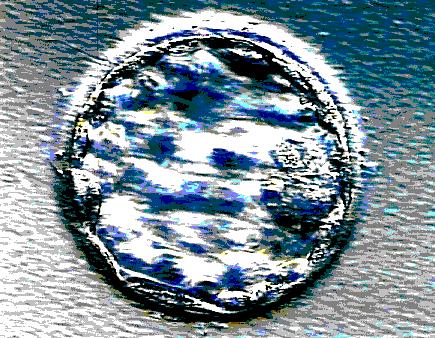
Figure 10. A blastocyst prior to
hatching and implantation.
Figure 11. The endometrium grows
under the
influence of oestrogen and progesterone.
Following ovulation (the luteal phase), now under the additional influence of progesterone (produced by the corpus luteum) as well as oestrogen, the endometrium and its glands undergo rapid morphological changes, demonstrating secretory activity (term which indicates receptivity regarding the potential for implantation) see figure 11. However, the function of these structural changes remains unclear; factors other than gonadal steroids influence uterine receptivity leading to a hypothesis that the uterine epithelium acts as a barrier to the embryo, only becoming receptive for implantation when appropriately conditioned by oestrogen and progesterone.Additionally, other local endocrine events may be involved such as a variety of interactions between growth factors including epidermal growth factor (EGP), insulin-like growth factor-1 (IGF-1) and its binding protein (1GF-BP-I), prostaglandin (notably PGE2), plasminogen activators and possibly leukaemia inhibiting factor (LIF).
The embryo after remaining in the fallopian tube for 3-4 days until reaching morula stage (late 8-32 cell stage), proceeds via the tubal isthmus to the uterine cavity where it will float freely for up to a further 72 hours. The embryonic disc (sometime flippantly referred to as the embryo on a compact disc by tabloid journalists!) is thought to orientate itself towards the endometrium and from the sixth day, will begin to penetrate the endometrium which responds by increased membrane activity (see below), producing extracellular matrix tissue (ECM) by de novo synthesis, which is thought to enhance the chances of implantation. This process facilitates one of the earliest embryo-maternal interactions such as secretion of human chorionic gonadotrophin by the trophoblastic cells of the embryo into the circulation leading to the maternal recognition of pregnancy (there appear to be many changes which involve modulation of the immune responses within the womb).
The binding to ECM (glycoproteins) within basement membranes (laminin and fibronectin) through receptors (integrins-recognition site which often initiates a cellular response to the stimulant that binds to it); activation of proteases and pericellular degradation of matrix components (matrix metallo proteinases); migration of cells into the matrix, i.e. cells progressing through the endometrial epithelium towards the basement membrane, recognising laminin and collagen type IV through their integrins. All these changes allow the invading cytotrophoblast cells of the embryo easier access to the endometrium than they would have in the absence of the complex changes described above. Indeed, de novo expression of the gene for gelatinases A & B is activated, causing breakdown of the basement membrane permitting cytotrophoblastic cells to make contact with the extracellular matrix (sub-epithelial layer of the endometrium) through their fibronectin receptors (see figure 12).
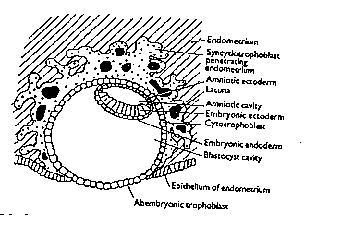
Figure 12. Implanting conceptus showing bilaminar disc.
Endometrial cytokines modulate cytotrophoblastic proteolytic activity to control the depth of invasion. By 7-8 days after fertilisation, the embryo is fully embedded within the endometrial stroma, the trophoblast having already differentiated into cytotrophoblastic and invasive syncytotrophoblast. The syncytial mass (the developing embryo) at nine days is 500-600um in diameter, with predecidual cells surrounding the embryonic mass (about 100um in diameter). Increased epithelial vasculature at the implantation site is observed due to oedema and localised hyperaemia, at 11-12 days, the implantation site being seen as a 1mm red spot on the mucosa due to maternal blood in lacunar spaces. By 14-21 days, the trophoblastic structure at the periphery of the embryo resembles the villi of the mature placenta as the inner cell mass has begun to undergo embryogenesis.
The conceptus at implantation, which occurs 6-8 days after fertilisation, has reached the blastocyst stage (Figure 10) and weighs less than 1 ug. Subsequent differentiation occurs rapidly from the trilaminar disc at 16 days (see Table I).Embryogenesis describes the time period from conception until eight weeks, the embryo being evident ultrasonically from about 30 days gestation by observation of the secondary yolk sac and 42-49 days by the pulsation in the pericardial cavity, a bulge below the developing facial and pharyngeal regions (Figure 12).
Figure 12. A beating heart may be observed at about 6-7 weeks.
The rate of growth of the embryo is rapid being 1mm per day until the end of embryogenesis (crown rump length 30mm) increasing to 1.5mm/day in the early fetal period. The major organ systems develop rapidly, a recognisable human form being evident by 49 days (Fig 13).
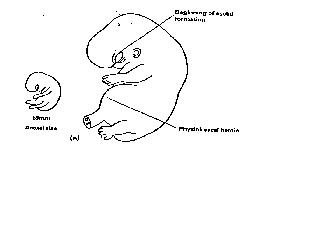
Figure 13. Organogenesis complete.
- Before implantation may take place, a number of events must take place
- Embryos must synthesise integrins (cell adhesion molecules; CAMs)
- The womb lining must synthesise fibronection and other extracellular matrix (ECM) components
- The abovementionned activities are likely to be mediated by growth factors and cytokines. Both the womb lining and the embryo are likely to make and release these factors
- The embryo must also successfully produce cytotrophoblastic and synctiotrophoblastic tissue in order to successfully infiltrate the endometrial stroma (the ECM)
- Implantation is a complex process
- The womb is not a privileged immunological site, contrary to public myth
- The embryo and fetus are foreign to the mother (host)
- Trophoblastic cells have been found to exhibit few antigens
- It is now thought that cytokines are involved in initiating a localised immunosuppression, which would allow the developing embryo to avoid a rejection-like response from the womb
- Leukocyte infiltration of the implantation site has been observed. Most of these seem to be T-suppressor cells.
The fetal-maternal allograft paradox even perplexed Nobel Prize winner Peter Medawar (Clinical Immunologist-Transplant Pioneer)
