| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | SEARCH RESULT |
J. Biol. Chem., Vol. 275, Issue 50, 39125-39129, December 15,
2000
|
| |||||||
|
|

From the Karolinska Institute, Division of Clinical Virology, Huddinge University Hospital, S-141 86 Stockholm, Sweden
Received for publication, July 13, 2000, and in revised form, September 12, 2000
| |
ABSTRACT |
|---|
Nucleoside kinases from several species are investigated as "suicide genes"
for treatment of malignant tumors by combined gene/chemotherapy. We
have recently cloned a multisubstrate deoxyribonucleoside kinase of
Drosophila melanogaster (Dm-dNK), and we have shown
that the enzyme phosphorylates cytotoxic pyrimidine and purine
nucleoside analogs. The broad substrate specificity of the enzyme,
as well as its very high catalytic rate, makes it a unique member
of the nucleoside kinase enzyme family. In the present study,
we evaluated Dm-dNK as a suicide gene by constructing a
replication-deficient retroviral vector that expresses the enzyme.
The human pancreatic adenocarcinoma cell line MIA PaCa-2 and a
thymidine kinase-deficient osteosarcoma cell line were transduced
with the recombinant virus. We showed that Dm-dNK can be
expressed in human cells, that the enzyme retained its enzymatic
activity, and that it is localized in the cell nuclei due to a
nuclear localization signal in its C-terminal region. The cells
expressing Dm-dNK exhibited increased sensitivity to several
cytotoxic nucleoside analogs, such as 1- The transfer of the gene encoding herpes simplex virus type-1 thymidine
kinase (HSV-1 TK)1 into malignant cells and subsequent treatment with
ganciclovir is one of the most commonly studied strategies of suicide
gene therapy (1-5).
The HSV-1 TK gene is introduced into cancer cells using either viral
or non-viral vectors. The nucleoside kinase phosphorylates nucleoside
analogs such as the guanosine analog ganciclovir to their
monophosphate derivatives, and cellular enzymes further phosphorylate
the compounds to their triphosphate forms. Ganciclovir triphosphate
interferes with DNA replication (6)
and induces cell death, probably by apoptosis (7,
8).
In addition to affecting the cells expressing HSV-1 TK, adjacent
untransduced cancer cells are killed by the transfer of
phosphorylated nucleoside analog between cells via gap junctions (7,
9).
This phenomenon, known as the "bystander effect," results in killing
of a larger portion of cells than those transduced with the suicide
gene. One limiting factor for the efficiency of suicide gene therapy is the kinetic
properties of the "suicide" nucleoside kinase. Genetically engineered
mutants of HSV-1 TK with improved kinetic properties for nucleoside
analog phosphorylation show increased efficiencies as suicide genes
(10-12).
Nucleoside kinases from different members of the herpesvirus family,
such as herpes simplex virus type 2 and varicella zoster virus,
have also been studied for possible use as suicide genes (13-16).
The human nucleoside kinases deoxycytidine kinase and deoxyguanosine
kinase enhance sensitivity to cytotoxic nucleoside analogs and are
also candidate genes for gene therapy (17-19).
Munch-Petersen and co-workers (20)
have purified a nucleoside kinase from Drosophila melanogaster that
catalyzed the phosphorylation of all the natural pyrimidine and
purine deoxyribonucleosides. In addition to its broad substrate
specificity, the enzyme also exhibited a high catalytic rate that is
10-100-fold higher than reported for the previously studied
nucleoside kinases. The broad substrate specificity of this enzyme,
together with its high catalytic rate, makes it a unique member of
the human and viral nucleoside kinase enzyme family. We have recently
cloned the cDNA of this multisubstrate enzyme, named
D. melanogaster deoxyribonucleoside kinase
(Dm-dNK), and shown that it also efficiently phosphorylates
several anti-viral and anti-cancer nucleoside analogs (21).
In the present study, we decided to evaluate the possible use
of Dm-dNK as a suicide gene by expressing the enzyme in human
cancer cell lines. In summary, we have shown that Dm-dNK can
be expressed in human cells with retained enzymatic activity and
that it increases the sensitivity of the cells to several cytotoxic
nucleoside analogs. Construction of a Retrovirus Vector Expressing Dm-dNK-- We used a
retrovirus vector based on the Moloney murine leukemia virus to generate a
replication-deficient recombinant retrovirus containing the cDNA of
Dm-dNK. Oligonucleotide primers containing engineered
EcoRI and XhoI restriction enzyme sites were designed
flanking the open reading frame of Dm-dNK cDNA
(5'-AAGAATTCGGACTGATGGCGGAGGCAGCATCC and
5'-TTCTCGAGTGGTTATCTGGCGACCCTCTGGC). The primers were used in a
polymerase chain reaction, and the DNA fragment was cloned into the
EcoRI-XhoI site of the pLXSN plasmid vector (CLONTECH). The
plasmid was purified using the NucleoBond plasmid purification kit
(CLONTECH). The DNA
sequence of the constructed plasmid was verified by DNA sequence
determination using an ABI310 automated DNA sequencer (PerkinElmer
Life Sciences).
RetroPack PT67 packaging cells (CLONTECH) were cultured at
37 °C in Dulbecco's modified Eagle's medium supplemented with 10%
(v/v) fetal calf serum (Life Technologies, Inc.), 100 units/ml
penicillin, and 0.1 mg/ml streptomycin. The constructed pLXSN
plasmid vector was transfected into the packaging cells using
LipofectAMINE (Life Technology. Inc.) according to the protocol
provided by the supplier. The medium from the transfected cells
was collected 48 h after transfection, filtered through a
0.45-µm filter, and diluted 2-fold with fresh medium. The
virus-containing medium was subsequently used to transduce the cancer
cell lines as described below.
Cell Culture and Retroviral Transduction-- TK-deficient osteosarcoma
cells was a kind gift from Prof. J. Balzarini, Rega Institute, Leuven,
Belgium. MIA PaCa-2 human pancreatic adenocarcinoma cells were
purchased from the American Type Culture Collection. All cells were
cultured in Dulbecco's modified Eagle's medium supplemented with 10%
(v/v) fetal calf serum (Life Technologies, Inc.), 100 units/ml
penicillin, and 0.1 mg/ml streptomycin. Cells were grown at
37 °C in a humidified incubator with a gas phase of 5%
CO2. The cells lines were transduced with the retrovirus
containing medium mixed with 4 µg/ml Polybrene. The cells were
incubated for 48 h and then cultured continuously for
3 weeks in the presence of 1.0 mg/ml geneticin (Life
Technologies, Inc.).
Enzyme Assays-- Cell protein extracts were prepared as described (22).
The assays were performed in 50 mM Tris-HCl, pH
7.6, 5 mM MgCl2,
5 mM ATP, 2 mM
dithiothreitol, 15 mM NaF, 100 mM KCl, 0.5 mg/ml bovine serum albumin, and
0.6 µg of protein extract in a total volume of 35 µl.
2.5 µM [methyl-3H]dThd (Moravek
Biochem), 3 µM [5-3H]-1- Autoradiography-- The cells were cultured on poly-L-lysine-coated chamber slides (Nunc, Inc.) for 24 h and
subsequently labeled with [methyl-3H]dThd (Moravek Biochem)
for 12 h. The slides were rinsed twice with phosphate-buffered
saline, fixed 10 min in methanol:acetic acid (3:1), and washed
three times with ice-cold 10% trichloroacetic acid, once with water,
and once with methanol. The slides were coated with Hypercoat
photoemulsion (Amersham Pharmacia Biotech) and exposed 1-3 weeks at
4 °C. The autoradiograms were developed using D-11 developer
(Eastman Kodak Co.).
Subcellular Localization of Dm-dNK-- We used the pEGFP-N1 plasmid
vector (CLONTECH) to
express the Dm-dNK cDNA in fusion with the green fluorescent protein
(dNK-GFP). Oligonucleotide primers containing engineered EcoRI
and SalI restriction enzyme sites were used to clone the open
reading frame of Dm-dNK cDNA
(5'-AAGAATTCGGACTGATGGCGGAGGCAGCATCC and 5'-ATGTCGACCTGGCGACCCTCTGGCGCTTGC)
into the EcoRI-SalI site of the pEGFP-N1 vector. The
C-terminal mutations ( Cell Proliferation Assays-- 9- Expression of Dm-dNK in Mammalian Cells-- We used a retrovirus vector
based on the Moloney murine leukemia virus to create replication-deficient
recombinant retroviridae with (dNK-pLXSN) and without (pLXSN) the
Dm-dNK cDNA (Fig. 1).
A TK1-deficient human osteosarcoma cell line and an MIA PaCa-2
human pancreatic adenocarcinoma cell line were transduced with
the retroviridae, and polyclonal populations of stably transfected
cells were selected.
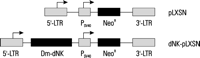
We used autoradiography to visualize in situ incorporation of
[3H]dThd into DNA of the thymidine kinase 1-deficient
osteosarcoma cells (Fig. 2).
The wild-type untransduced cells and the cells transduced with the
pLXSN vector alone showed a faint dotted autoradiography pattern
distributed throughout the cells, indicating phosphorylation of dThd
by mitochondrial thymidine kinase 2 and its subsequent
incorporation into mitochondrial DNA (23-24).
In contrast, the cells expressing Dm-dNK exhibited dark
staining of the cell nucleus indicating incorporation of
[3H]dThd into nuclear DNA. Approximately 90% of the cells
incorporated dThd in nuclear DNA, indicating that the majority of the
transduced cancer cells expressed active Dm-dNK.
To quantify the total level of nucleoside kinase activity in the cells, we
determined the phosphorylation of dThd, CdA, and araC in cell
protein extracts (Fig. 3).
Untransfected osteosarcoma cells deficient in cytosolic thymidine
kinase 1 activity showed low levels of dThd phosphorylation,
probably catalyzed by mitochondrial thymidine kinase 2 (23).
The cells transduced with the pLXSN retroviral vector alone showed
similar levels of dThd phosphorylation as the wild-type cells,
whereas the cells transfected with dNK-pLXSN exhibited
Expression of Dm-dNK Increased Sensitivity to Nucleoside Analogs-- We
determined the sensitivity of the untransduced cells and the cells transduced
with either the retroviral vector alone or the dNK-pLXSN vector for
several cytotoxic nucleoside analogs (Table I).
The cytotoxicity (IC50) was determined after 4 days
of drug exposure as described under "Experimental Procedures."
The difference in sensitivity between the wild-type cell lines
and the cells transduced with the pLXSN vector without Dm-dNK
was less than 3-fold for all investigated compounds. Both the
osteosarcoma cells and the pancreatic cancer cells that expressed
Dm-dNK showed an increase in sensitivity to several of the
nucleoside analogs. The highest increase in sensitivity for the
osteosarcoma cell was detected for BVDU, CdA, dFdC, and
FdUrd with a 100-400-fold decrease in IC50 compared with
the untransduced cells. araC and araT showed
The Dm-dNK Is Localized in the Nucleus of Human Cells-- In order to
determine the subcellular location of the Dm-dNK when overexpressed in
human cells, we constructed a fusion of Dm-dNK and GFP to
visualize the protein in vivo (dNK-GFP) (Fig. 4).
The cells transfected with a plasmid encoding the fusion protein
exhibited green fluorescence predominantly in the nucleus of the
cells. The nuclear location was verified by contra-staining the
cells with DAPI. Based on sequence analysis Munch-Petersen and
co-workers (25)
suggested two possible nuclear localization signals starting at
residue 95 (PTNKKLK) and 242 (PSKRQRV). We decided to test
the role of the putative nuclear localization signal in the C
terminus of the protein in an experiment using site-directed
mutagenesis. The putative nuclear localization signal was mutated by
either replacing Arg-247 with a serine residue (
Here we have shown that the multisubstrate nucleoside kinase of
D. melanogaster retains its enzymatic activity when expressed
in human cells and that the activity results in an enhanced
sensitivity to several cytotoxic pyrimidine and purine nucleoside
analogs. These results suggest that Dm-dNK may be used as a
suicide gene in combined gene/chemotherapy of cancer. Two of the most
promising candidate nucleoside analogs to use in conjunction with
Dm-dNK gene transfer are the pyrimidine analogs dFdC and
BVDU. dFdC is currently used in the treatment of several
types of solid tumors such as pancreatic adenocarcinoma and non-small
cell lung cancer (26).
It may be possible to increase the efficiency of this therapy by
transduction of tumor cells in vivo with the Dm-dNK
gene. BVDU, the second candidate nucleoside analog, was initially
developed as an anti-herpetic agent, and the compound has been
used to treat patients with varicella zoster infections (27).
BVDU shows low toxicity to untransduced cells, but the cells
expressing Dm-dNK are highly sensitive to the compound. The
concurrent use of both a nucleoside analog that is restricted to
Dm-dNK for activation, such as BVDU, and a nucleoside
analog that already has shown clinical efficacy in treatment of
malignant tumors, such as dFdC, is one possible approach to
develop new treatment modalities for tumors with combined gene and
chemotherapy. Combining the therapy with other anti-metabolites to
achieve synergistic effects may be another approach to enhance the
efficiency of nucleoside kinase suicide gene therapy. Recent studies
on HSV-1 TK-transduced cancer cells suggest that synergistic
cytotoxicity and enhanced bystander effects may be achieved when
combining ganciclovir with the ribonucleotide reductase inhibitor
hydroxyurea (28).
Inhibition of ribonucleotide reductase results in decreased de
novo dNTP synthesis, which is favorable for the incorporation of
nucleoside analogs into DNA. Furthermore, a decrease of the dNTP pool
may also increase the phosphorylation of nucleoside analogs because
several nucleoside kinases are feedback-inhibited by dNTPs. The
nucleoside analogs dFdC and CdA are both potent inhibitors of
ribonucleotide reductase once phosphorylated to their diphosphate
forms. Because Dm-dNK phosphorylates both of these compounds,
it is possible that they will inhibit ribonucleotide reductase and
thereby enhance the phosphorylation of BVDU or other nucleoside
analogs that are activated by Dm-dNK. We are presently
initiating studies on tumor models to investigate further the
possible use of Dm-dNK as a suicide gene in vivo and to
evaluate the possible benefits of combining multiple nucleoside
analogs to improve this therapeutic regime. The unique features of Dm-dNK as a suicide gene compared with HSV-1 TK
and other nucleoside kinases are its broad substrate specificity and
high catalytic rate. Pyrimidine deoxyribonucleosides have in
vitro higher affinity to the enzyme compared with purine
deoxyribonucleosides (20,
21,
25).
The difference in affinity between purine and pyrimidine nucleosides
is also observed for some, but not all, nucleoside analogs. Enzymatic
assays performed with recombinant Dm-dNK show that pyrimidine
nucleoside analogs such as BVDU, FdUrd, araC, and araT
compete more efficiently with the natural pyrimidine substrates dThd
and dCyd compared with most of the investigated purine nucleoside
analogs such as araG and araA (21).
However, one of the exceptions is the clinically important purine
nucleoside analog CdA, which efficiently competes with both dThd
and dCyd for phosphorylation by Dm-dNK. This compound did also
show a marked increase in cytotoxicity in the Dm-dNK-transfected
cells. Studies on HSV-1 TK-transfected cells suggest that the
purine phosphorylating activity in vivo may be increased by
mutations that decreases the enzymes affinity to pyrimidine
nucleoside analogs (29).
We are currently initiating studies to investigate if a similar
approach can be used to enhance purine nucleoside analog
phosphorylation catalyzed by Dm-dNK. Dm-dNK is sequence related to the human nucleoside kinases
deoxycytidine kinase, deoxyguanosine kinase, and thymidine kinase
2 (21).
The human enzymes differ in their subcellular location by being
located in either the nucleus, cytosol, or mitochondria (23,
30-32).
In the present study we have shown that Dm-dNK is targeted to
the cell nucleus when expressed in human cell lines. Two putative
nuclear localization signal sequences have been identified in the
Dm-dNK sequence (25).
We showed that two independent mutants of the putative nuclear
localization signal located in the C terminus of the protein
completely abolished its nuclear translocation. These experiments
showed that this signal is the physiologically important signal
mediating the nuclear localization. We do not yet know whether the
nuclear location of the enzyme is restricted to the mammalian cells
used in this study or whether the enzyme is also located in the
nucleus of D. melanogaster cells as well. Interestingly,
HSV-1 TK is also located in the cell nucleus when expressed in human
cells (33).
The physiological importance of the nuclear location of nucleoside
kinases is, however, yet unclear, since studies suggest that
phosphorylated nucleoside analogs may freely traverse the nuclear
envelope, and no differences in nucleoside analogs sensitivity have
been detected when nucleoside analogs are phosphorylated in either of
the two subcellular compartments (30).
The success of suicide gene therapy is dependent on efficient gene delivery
systems and the selective expression of the suicide gene in tumor
cells. Most of the in vivo studies on nucleoside kinase
suicide gene therapy have been performed using first generation
retro- or adenovirus vector systems (2-5).
These systems show insufficient vector distribution and low
transduction efficiencies. Recently, significant achievements in
vector developments have been made for both viral and non-viral
systems (34).
Progress has also been made in achieving selective and targeted
expression of the suicide gene in tumor cells. Several
tissue-specific promoters have been shown to direct transgene
expression to a certain tissue or cell type. These promoters include
the tyrosinase promoter for transgene expression in melanocytes that
may be used for targeting melanoma cells (35),
the prostate-specific antigen promoter for selective gene expression
in prostate and prostate cancer tissue (36),
and the glial fibrillary acidic protein promoter for gene expression
in brain tumors of glial origin (37).
Several promoters of tumor marker proteins, such as the
carcinoembryonic antigen (38)
and the 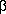 -D-arabinofuranosylcytosine, 1--D-arabinofuranosylthymine,
(E)-5-(2-bromovinyl)-2'-deoxyuridine,
2-chloro-2'-deoxyadenosine, and 2',2'-difluorodeoxycytidine. These
findings suggest that Dm-dNK may be used as a suicide gene in
combined gene/chemotherapy of cancer.
-D-arabinofuranosylcytosine, 1--D-arabinofuranosylthymine,
(E)-5-(2-bromovinyl)-2'-deoxyuridine,
2-chloro-2'-deoxyadenosine, and 2',2'-difluorodeoxycytidine. These
findings suggest that Dm-dNK may be used as a suicide gene in
combined gene/chemotherapy of cancer.
![]()
INTRODUCTION
![]() TOP
TOP ![]() ABSTRACT
ABSTRACT ![]() INTRODUCTION
INTRODUCTION ![]() EXPERIMENTAL PROCEDURES
EXPERIMENTAL PROCEDURES ![]() RESULTS
RESULTS ![]() DISCUSSION
DISCUSSION ![]() REFERENCES
REFERENCES
![]()
EXPERIMENTAL PROCEDURES
![]() TOP
TOP ![]() ABSTRACT
ABSTRACT ![]() INTRODUCTION
INTRODUCTION ![]() EXPERIMENTAL
PROCEDURES
EXPERIMENTAL
PROCEDURES ![]() RESULTS
RESULTS ![]() DISCUSSION
DISCUSSION ![]() REFERENCES -D-arabinofuranosylcytosine (araC) (Moravek Biochem), or
3 µM
[8-3H]-2-chloro-2'-deoxyadenosine (CdA) (Moravek Biochem) were
used in the assays and mixed with equivalent amounts of unlabeled
substrates. The araC phosphorylation assay mixtures were
supplemented with 50 µM tetrahydrouridine
to inhibit deamination of araC by cytidine deaminase. Aliquots of
the reaction mixture were spotted on Whatman DE-81 filters after
10-, 20-, and 30-min incubation at 37 °C. The filters
were washed three times in 5 mM ammonium formate.
The nucleoside monophosphates were eluted from the filter with
0.5 M KCl, and the radioactivity was determined
by scintillation counting.
REFERENCES -D-arabinofuranosylcytosine (araC) (Moravek Biochem), or
3 µM
[8-3H]-2-chloro-2'-deoxyadenosine (CdA) (Moravek Biochem) were
used in the assays and mixed with equivalent amounts of unlabeled
substrates. The araC phosphorylation assay mixtures were
supplemented with 50 µM tetrahydrouridine
to inhibit deamination of araC by cytidine deaminase. Aliquots of
the reaction mixture were spotted on Whatman DE-81 filters after
10-, 20-, and 30-min incubation at 37 °C. The filters
were washed three times in 5 mM ammonium formate.
The nucleoside monophosphates were eluted from the filter with
0.5 M KCl, and the radioactivity was determined
by scintillation counting.
 247/dNK-GFP and 244-245/dNK-GFP) were
introduced by using two extended 3'-oligonucleotide primers
(5'-ATGTCGACCTGGCGACGCTCTGGCGCTTGCTGGGC and
5'-ATGTCGACCTGCGACCCTCTGGCTCTCGCTGGGCGACAC). The plasmids were
transfected into the cell lines using LipofectAMINE (Life
Technologies, Inc.) as described above. The nuclei of the cells
were counterstained with 4',6'-diamidino-2-phenylindole (DAPI).
GFP and DAPI fluorescence was observed in a Nikon Eclipse
E600 microscope equipped with a SPOT RT digital camera.
-D-Arabinofuranosyladenine
(araA), araC, 1--D-arabinofuranosylthymine
(araT), ganciclovir, and CdA were obtained from Sigma.
2',2'-Difluorodeoxycytidine (dFdC) was obtained from Lilly.
9--D-Arabinofuranosylguanine (araG),
(E)-5-(2-bromovinyl)-2'-deoxyuridine (BVDU), and
5-fluoro-2'-deoxyuridine (FdUrd) were gifts from Prof.
J. Balzarini, Rega Institute, Leuven, Belgium. The cells were
plated at
247/dNK-GFP and 244-245/dNK-GFP) were
introduced by using two extended 3'-oligonucleotide primers
(5'-ATGTCGACCTGGCGACGCTCTGGCGCTTGCTGGGC and
5'-ATGTCGACCTGCGACCCTCTGGCTCTCGCTGGGCGACAC). The plasmids were
transfected into the cell lines using LipofectAMINE (Life
Technologies, Inc.) as described above. The nuclei of the cells
were counterstained with 4',6'-diamidino-2-phenylindole (DAPI).
GFP and DAPI fluorescence was observed in a Nikon Eclipse
E600 microscope equipped with a SPOT RT digital camera.
-D-Arabinofuranosyladenine
(araA), araC, 1--D-arabinofuranosylthymine
(araT), ganciclovir, and CdA were obtained from Sigma.
2',2'-Difluorodeoxycytidine (dFdC) was obtained from Lilly.
9--D-Arabinofuranosylguanine (araG),
(E)-5-(2-bromovinyl)-2'-deoxyuridine (BVDU), and
5-fluoro-2'-deoxyuridine (FdUrd) were gifts from Prof.
J. Balzarini, Rega Institute, Leuven, Belgium. The cells were
plated at 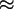 2000 cells/well in 96-well plates. Nucleoside
analogs were added after 24 h, and the medium containing the
nucleoside analogs was changed daily. Cell survival was assayed by a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
(Roche Molecular Biochemicals) after 4 days of drug exposure.
Each experiment was performed in triplicate. The IC50
value of the investigated compounds was calculated as the mean
value of these experiments.
2000 cells/well in 96-well plates. Nucleoside
analogs were added after 24 h, and the medium containing the
nucleoside analogs was changed daily. Cell survival was assayed by a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
(Roche Molecular Biochemicals) after 4 days of drug exposure.
Each experiment was performed in triplicate. The IC50
value of the investigated compounds was calculated as the mean
value of these experiments.
![]()
RESULTS
![]() TOP
TOP ![]() ABSTRACT
ABSTRACT ![]() INTRODUCTION
INTRODUCTION ![]() EXPERIMENTAL PROCEDURES
EXPERIMENTAL PROCEDURES ![]() RESULTS
RESULTS ![]() DISCUSSION
DISCUSSION ![]() REFERENCES
REFERENCES
View larger version
(8K):
[in a new window]
Fig. 1.
Retroviral vector (pLXSN) used to insert the Dm-dNK
cDNA (dNK-pLXSN). LTR, long terminal repeat;
PSV40, SV40 large T-antigen promoter;
NeoR, neomycin resistance gene.
View larger version (87K):
[in a new window]
Fig. 2.
[3H]dThd autoradiography of TK-deficient osteosarcoma
cells transduced with pLXSN (A) or dNK-pLXSN (B).
100-fold higher
enzymatic activity than the parent cell line. Untransfected MIA
PaCa-2 cells exhibited a higher basal level of dThd phosphorylating
activity compared with the osteosarcoma cells due to the retained
expression of TK1 in the pancreatic cells. However, the Dm-dNK
expression increased the dThd kinase activity 35-fold in the MIA PaCa-2 cells as
well. The phosphorylation of CdA and araC in the cell protein
extracts were also increased 4-15-fold in the
Dm-dNK-transduced osteosarcoma and pancreatic adenocarcinoma
cells compared with the untransfected parent cell lines and the cells
transfected with pLXSN vector alone (Fig. 3).
In summary, these experiments showed that human cancer cells
transduced with the dNK-pLXSN retroviral vector expressed
enzymatically active Dm-dNK and that the expression resulted
in an increase of nucleoside and nucleoside analog phosphorylation.
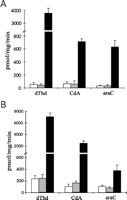
View larger version (13K):
[in a new window]
Fig. 3.
[3H]dThd, [3H]CdA, and
[3H]araC phosphorylating activity in
cell protein extracts. A, osteosarcoma cells; B,
MIA PaCa-2 pancreatic adenocarcinoma cells. Wild-type cells (open
bars), pLXSN-transduced cells (gray bars), and
dNK-pLXSN-transduced cells (black bars). 50-fold reduction in
IC50, whereas araA and ganciclovir showed between
3- and 9-fold reduction in IC50 compared with the
untransduced cells. araG was not toxic to either cell line at
the investigated concentrations. The highest increase in sensitivity
for the MIA PaCa-2 cells was observed for BVDU with a
>6400-fold increased sensitivity. araT and dFdC increased
the sensitivity 30- and 75-fold, respectively. CdA showed a
6-fold increase in sensitivity in the Dm-dNK-expressing cells,
whereas the sensitivity of araA, araC, FdUrd, and ganciclovir
was not enhanced by Dm-dNK expression in this cell line.
Sensitivity (IC50) of
osteosarcoma and MIA PaCa-2 pancreatic adenocarcinoma cell lines to
nucleoside analogs 247/dNK-GFP) or by replacing
Lys-244 and Arg-245 with glutamic acid and serine amino acid
residues, respectively (244-245/dNK-GFP). When expressed in
the osteosarcoma cell line, 247/dNK-GFP and 244-245/dNK-GFP were both retained
in the cytosol and did not translocate to the cell nucleus (Fig. 4).
In summary, these experiments showed that the nuclear localization of
Dm-dNK in human cells is mediated by the C-terminal signal
sequence.
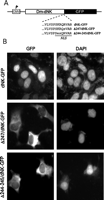
View larger version (52K):
[in a new window]
Fig. 4.
Subcellular localization of Dm-dNK fused to
GFP. A, constructs used to express
Dm-dNK in fusion with GFP (dNK-GFP) and the two
mutants ( 247/dNK-GFP and 244-245/dNK-GFP). Mutated amino
acid are shown in lowercase letters. B, fluorescence
microscopy of cells transfected with the plasmids. GFP
fluorescence and DAPI nuclear contra-staining showed that the
wild-type dNK-GFP was located in the nucleus and that the mutants
were located in the cytosol. CMV, cytomegalovirus;
NLS, nuclear localization signal.
![]()
DISCUSSION
![]() TOP
TOP ![]() ABSTRACT
ABSTRACT ![]() INTRODUCTION
INTRODUCTION ![]() EXPERIMENTAL PROCEDURES
EXPERIMENTAL PROCEDURES ![]() RESULTS
RESULTS ![]() DISCUSSION
DISCUSSION ![]() REFERENCES
REFERENCES  -fetoprotein (39),
can also be used to control selective transgene expression in tumor
cells. Novel vector systems also utilizes receptors specific for
tumor cells or other phenotypic alterations of the cancer cells to
restrict vector transduction or replication to tumor cells (34).
The recently developed oncolytic adenovirus ONYX-015 utilizes
the loss of p53 function in many types of human tumors for
selectivity. The ONYX-015 adenovirus cannot replicate in cells with
intact p53 (40),
and the virus thus only kills cells deficient in p53 expression,
whereas normal cells are not killed by the virus. Initial clinical
studies using the ONYX-015 virus for local treatment of tumors show
promising results (41),
and it has been suggested that the efficacy may be further enhanced
by the concomitant use of a suicide gene expressed by the virus (41-42).
In the future, it will be important to evaluate these novel vectors
and expression system using the optimal prodrug/suicide gene
combination to achieve the most efficient killing of transduced tumor
cells as well as killing of untransduced tumor via bystander
effects.
-fetoprotein (39),
can also be used to control selective transgene expression in tumor
cells. Novel vector systems also utilizes receptors specific for
tumor cells or other phenotypic alterations of the cancer cells to
restrict vector transduction or replication to tumor cells (34).
The recently developed oncolytic adenovirus ONYX-015 utilizes
the loss of p53 function in many types of human tumors for
selectivity. The ONYX-015 adenovirus cannot replicate in cells with
intact p53 (40),
and the virus thus only kills cells deficient in p53 expression,
whereas normal cells are not killed by the virus. Initial clinical
studies using the ONYX-015 virus for local treatment of tumors show
promising results (41),
and it has been suggested that the efficacy may be further enhanced
by the concomitant use of a suicide gene expressed by the virus (41-42).
In the future, it will be important to evaluate these novel vectors
and expression system using the optimal prodrug/suicide gene
combination to achieve the most efficient killing of transduced tumor
cells as well as killing of untransduced tumor via bystander
effects.
| |
FOOTNOTES |
|---|
* This work was supported by grants from the Swedish Medical Research Council, the Swedish Cancer Foundation, the Swedish Foundation of Strategic Research, and the European Commission.The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
.gif) To whom
correspondence should be addressed. Tel.: 46-8-58587932; Fax 46-8-58587933;
E-mail: [email protected].
To whom
correspondence should be addressed. Tel.: 46-8-58587932; Fax 46-8-58587933;
E-mail: [email protected].
Published, JBC Papers in Press, September 18, 2000, DOI 10.1074/jbc.M006212200
| |
ABBREVIATIONS |
|---|
The abbreviations used are: HSV-1 TK, herpes simplex virus type-1 thymidine
kinase; araA, 9--D-arabinofuranosyladenine;
araC, 1--D-arabinofuranosylcytosine; araG, 9--D-arabinofuranosylguanine; araT, 1--D-arabinofuranosylthymine; BVDU,
(E)-5-(2-bromovinyl)-2'-deoxyuridine; CdA, 2-chloro-2'-deoxyadenosine;
DAPI, 4',6'-diamidino-2-phenylindole; dFdC, 2',2'-difluorodeoxycytidine;
Dm-dNK, Drosophila melanogaster deoxyribonucleoside kinase; GFP,
green fluorescent protein.
| |
REFERENCES |
|---|
| 1. | Moolten, F. L. (1986) Cancer Res. 46,
5276-5281 |
| 2. | Moolten, F. L., and Wells, J. M. (1990) J. Natl. Cancer
Inst. 82, 297-300 |
| 3. | Culver, K. W., Ram, Z., Wallbridge, S., Ishii, H.,
Oldfield, E. H., and Blaese, R. M. (1992) Science 256,
1550-1552 |
| 4. | Ram, Z., Culver, K. W., Oshiro, E. M., Viola, J. J.,
DeVroom, H. L., Otto, E., Long, Z., Chiang, Y., McGarrity, G. J., Muul, L.
M., Katz, D., Blaese, R. M., and Oldfield, E. H. (1997) Nat. Med.
3, 1354-1361 |
| 5. | Klatzmann, D., Valéry, C. A., Bensimon, G., Marro, B.,
Boyer, O., Mokhtari, K., Diquet, B., Salzmann, J.-L., Philippon, J., and
the Study Group on Gene Therapy for Glioblastoma. (1998) Hum. Gene
Ther. 9, 2595-2604 |
| 6. | Reardon, J. E. (1989) J. Biol. Chem. 264,
19039-19044 |
| 7. | Freeman, S. M., Abboud, C. N., Whartenby, K. A., Packman,
C. H., Koeplin, D. S., Moolten, F. L., and Abraham, G. N. (1993) Cancer
Res. 53, 5274-5283 |
| 8. | Beltinger, C., Fulda, S., Kammertoens, T., Meyer, E.,
Uckert, W., and Debatin, K.-M. (1999) Proc. Natl. Acad. Sci.
U. S. A. 96, 8699-8704 |
| 9. | Mesnil, M., Piccoli, C., Tiraby, G., Willecke, K., and
Yamasaki, H. (1996) Proc. Natl. Acad. Sci. U. S. A.
93, 1831-1835 |
| 10. | Black, M. E., Newcomb, T. G., Wilson, H.-M. P., and Loeb,
L. A. (1996) Proc. Natl. Acad. Sci. U. S. A. 93,
3525-3529 |
| 11. | Christians, F. C., Scapozza, L., Crameri, A., Folkers, G.,
and Stemmer, W. P. C. (1999) Nat. Biotechnol. 17,
259-264 |
| 12. | Kokoris, M. S., Sabo, P., Adman, E. T., and Black, M. E.
(1999) Gene Ther. 6, 1415-1426 |
| 13. | Balzarini, J., Bohman, C., and De Clercq, E. (1993) J.
Biol. Chem. 268, 6332-6337 |
| 14. | Degrève, B., Andrei, G., Izquierdo, M., Piette, J., Morin,
K., Knaus, E. E., Wiebe, L. I., Basrah, I., Walker, R. T., De Clercq, E.,
and Balzarini, J. (1997) Gene Ther. 4, 1107-1114 |
| 15. | Grignet-Debrus, C., and Calberg-Bacq, C.-M. (1997) Gene
Ther. 4, 560-569 |
| 16. | Loubière, L., Tiraby, M., Cazaux, C., Brisson, E., Grisoni,
M., Zhao-Emonet, J. C., Tiraby, G., and Klatzmann, D. (1999) Gene
Ther. 6, 1638-1642 |
| 17. | Hapke, D. M., Stegmann, A. P. A., and Mitchell, B. S.
(1996) Cancer Res. 56, 2343-2347 |
| 18. | Manome, Y., Wen, P. Y., Dong, Y., Tanaka, T., Mitchell, B.
S., Kufe, D. W., and Fine, H. A. (1996) Nat. Med. 2,
567-573 |
| 19. | Zhu, C., Johansson, M., Permert, J., and Karlsson, A.
(1998) J. Biol. Chem. 273, 14707-14711 |
| 20. | Munch-Petersen, B., Piskur, J., and Sondergaard, L. (1998)
J. Biol. Chem. 273, 3926-3931 |
| 21. | Johansson, M., Van Rompay, A. R., Degrève, B., Balzarini,
J., and Karlsson, A. (1999) J. Biol. Chem. 274,
23814-23819 |
| 22. | Arnér, E. S. J., Spasokoukotskaja, T., and Eriksson,
S. (1992) Biochem. Biophys. Res. Commun. 188,
712-718 |
| 23. | Berk, A. J., and Clayton, D. A. (1973) J. Biol.
Chem. 248, 2722-2729 |
| 24. | Davis, A. F., and Clayton, D. A. (1996) J. Cell
Biol. 135, 883-893 |
| 25. | Munch-Petersen, B., Knecht, W., Lenz, C., Sondergaard, L.,
and Piskur, J. (2000) J. Biol. Chem. 275, 6673-6679 |
| 26. | Hui, Y. F., and Reitz, J. (1997) Am.
J. Health-Syst. Pharm. 54, 162-170 |
| 27. | Wildiers, J., and De Clercq, E. (1984) Eur.
J. Cancer Clin. Oncol. 20, 471-476 |
| 28. | Boucher, P. D., Ostruszka, L. J., and Shewach, D. S. (2000)
Cancer Res. 60, 1631-1636 |
| 29. | Kokoris, M. S., Sabo, P., Adman, E. T., and Black, M. E.
(1999) Gene Ther. 6, 1415-1426 |
| 30. | Johansson, M., Brismar, S., and Karlsson, A. (1997)
Proc. Natl. Acad. Sci. U. S. A. 94,
11941-11945 |
| 31. | Söderlund, J. C. F., and Arnér, E. S. J. (1995)
Adv. Exp. Med. Biol. 370, 201-204 |
| 32. | Wettin, K., Johansson, M., Zheng, X., Zhu, C., and Karlsson, A.
(1999) FEBS Lett. 460, 103-106 |
| 33. | Degrève, B., Johansson, M., De Clercq, E., Karlsson, A.,
and Balzarini, J. (1998) J. Virol. 72, 9535-9543 |
| 34. | Romano, G., Michell, P., Pacilio, C., and Giordano, A. D.
(2000) Stem Cells 18, 19-39 |
| 35. | Siders, W. M., Halloran, P. J., and Fenton, R. G. (1998)
Cancer Gene Ther. 5, 281-291 |
| 36. | Latham, J. P., Searle, P. F., Mautner, V., and James, N. D.
(2000) Cancer Res. 60, 334-341 |
| 37. | Chen, J., Bezdek, T., Chang, J., Kherzai, A. W.,
Willingham, T., Azzara, M., and Nisen, P. D. (1998) Cancer Res.
58, 3504-3507 |
| 38. | Cao, G., Kuriyama, S., Gao, J., Kikukawa, M., Cui, L.,
Nakatani, T., Zhang, X., Tsujinoue, H., Pan, X., Fukui, H., and Qi, Z.
(1999) Gene Ther. 6, 83-90 |
| 39. | Mawatari, F., Tsuruta, S., Ido, A., Ueki, T., Nakao, K.,
Kato, Y., Tamaoki, T., Ishii, N., and Nakata, K. (1998) Cancer Gene
Ther. 5, 301-306 |
| 40. | Bischoff, J. R., Kirn, D. H., Williams, A., Heise, C.,
Horn, S., Muna, M., Ng, L., Nye, J. A., Sampson-Johannes, A., Fattaey, A.,
and McCormick, F. (1996) Science 274, 373-376 |
| 41. | Khuri, F. R., Nemunaitis, J., Ganly, I., Arseneau, J.,
Tannock, I. F., Romel, L., Gore, M., Ironside, J., MacDougall, R. H.,
Heise, C., Randlev, B., Gillenwater, A. M., Bruso, P., Kaye, S. B., Hong,
W. K., and Kirn, D. H. (2000) Nat. Med. 6, 879-885 |
| 42. | Wildner, O., Blaese, R. M., and Morris, J. C. (1999)
Cancer Res. 59, 410-413 |
|
| |||||||
|
|
| HOME | HELP | FEEDBACK | SUBSCRIPTIONS | ARCHIVE | SEARCH | SEARCH RESULT |