|
|
Vol. 60, Issue 2, 262-266, August
2001
Division of Clinical Virology, Karolinska Institute, Huddinge University Hospital, Stockholm, Sweden
| |
Abstract |
|---|
The multisubstrate deoxyribonucleoside kinase of Drosophila melanogaster (Dm-dNK) can be expressed in human cells with retained enzymatic activity. The cells expressing Dm-dNK exhibit increased sensitivity to several cytotoxic nucleoside analogs. In this study, we further evaluated Dm-dNK as a potential novel suicide gene in combination with (E)-5-(2-bromovinyl)-2'-deoxyuridine (BVDU) as the prodrug. We used two human cancer cell lines transduced with a retrovirus encoding the Dm-dNK cDNA and investigated whether the cells expressing the enzyme can induce cell death of untransduced cells, a phenomenon known as the "bystander effect". A bystander effect was observed in a thymidine kinase-deficient human osteosarcoma cell line but not in the MIA PaCa-2 human pancreatic adenocarcinoma cell line. The cytotoxicity of BVDU increased in both cell lines when the compound was used in combination with subtoxic concentrations of hydroxyurea. Hydroxyurea also enhanced the bystander effect in the osteosarcoma cells, but not in the MIA PaCa-2 cells, treated with BVDU. These findings indicate that BVDU phosphorylated by Dm-dNK in transduced cancer cells may also induce bystander cell death in certain cell lines.
| |
Introduction |
|---|
The principle of suicide gene therapy is the transduction of cells with a
gene that encodes an enzyme that can convert an inactive prodrug into
a cytotoxic metabolite (Lal et al., 2000![]() ). A commonly used
suicide gene approach involves the transfer of the herpes simplex
virus type-1 thymidine kinase (HSV-1 TK) gene into malignant cells
and subsequent treatment with ganciclovir (GCV) (Balzarini et al.,
1985
). A commonly used
suicide gene approach involves the transfer of the herpes simplex
virus type-1 thymidine kinase (HSV-1 TK) gene into malignant cells
and subsequent treatment with ganciclovir (GCV) (Balzarini et al.,
1985![]() ; Moolten, 1986
; Moolten, 1986![]() ; Moolten and Wells,
1990
; Moolten and Wells,
1990![]() ; Culver et al., 1992
; Culver et al., 1992![]() ; Ram et al., 1997
; Ram et al., 1997![]() ; Klatzmann et al.,
1998
; Klatzmann et al.,
1998![]() ). The HSV-1 TK-expressing
tumor cells phosphorylate GCV to its cytotoxic triphosphate
derivative, which interferes with DNA replication (Reardon, 1989
). The HSV-1 TK-expressing
tumor cells phosphorylate GCV to its cytotoxic triphosphate
derivative, which interferes with DNA replication (Reardon, 1989![]() ) and induces cell death,
probably by apoptosis (Freeman et al., 1993
) and induces cell death,
probably by apoptosis (Freeman et al., 1993![]() ; Beltinger et al., 1999
; Beltinger et al., 1999![]() ). In addition to
affecting cells expressing HSV-1 TK, adjacent untransduced cancer
cells are killed by the transfer of the phosphorylated nucleoside
analog between cells (Freeman et al., 1993
). In addition to
affecting cells expressing HSV-1 TK, adjacent untransduced cancer
cells are killed by the transfer of the phosphorylated nucleoside
analog between cells (Freeman et al., 1993![]() ; Mesnil et al.,
1996
; Mesnil et al.,
1996![]() ). This phenomenon, known as
the "bystander effect", results in the killing of a larger portion of
cells than is transduced with the suicide gene. The mechanism of the
bystander effect is not fully understood. The uptake of apoptotic
vesicles released from dying cells by adjacent nontransduced cells
(Freeman et al., 1993
). This phenomenon, known as
the "bystander effect", results in the killing of a larger portion of
cells than is transduced with the suicide gene. The mechanism of the
bystander effect is not fully understood. The uptake of apoptotic
vesicles released from dying cells by adjacent nontransduced cells
(Freeman et al., 1993![]() ) and the passage of
phosphorylated GCV via gap junctions (Rubsam et al., 1999
) and the passage of
phosphorylated GCV via gap junctions (Rubsam et al., 1999![]() ; Boucher et al., 2000
; Boucher et al., 2000![]() ) have been proposed
as possible mechanisms.
) have been proposed
as possible mechanisms.
In contrast to the family of deoxyribonucleoside kinases characterized from
several species, Drosophila melanogaster contains a single
highly efficient deoxyribonucleoside kinase (Dm-dNK) that is
able to phosphorylate all four natural deoxyribonucleosides as well
as several clinically important antiviral and anticancer nucleoside
analogs (Munch-Petersen et al., 1998![]() ; Johansson et al.,
1999
; Johansson et al.,
1999![]() ). We recently evaluated
Dm-dNK as a suicide gene by expressing the enzyme in human
cancer cell lines using a replication-deficient retroviral vector
(Zheng et al., 2000
). We recently evaluated
Dm-dNK as a suicide gene by expressing the enzyme in human
cancer cell lines using a replication-deficient retroviral vector
(Zheng et al., 2000![]() ). We have shown that
Dm-dNK can be expressed in human cells and that the enzyme
retains its enzymatic activity. The cells expressing Dm-dNK
exhibited an increased sensitivity to several cytotoxic nucleoside
analogs, among which (E)-5-(2-bromovinyl)-2'-deoxyuridine
(BVDU) was one of the most efficient prodrug candidates.
). We have shown that
Dm-dNK can be expressed in human cells and that the enzyme
retains its enzymatic activity. The cells expressing Dm-dNK
exhibited an increased sensitivity to several cytotoxic nucleoside
analogs, among which (E)-5-(2-bromovinyl)-2'-deoxyuridine
(BVDU) was one of the most efficient prodrug candidates.
A major limitation of gene therapy for cancer at present is the inability to
transduce all the cancer cells in vivo (Moolten, 1986![]() ; Moolten and Wells, 1990
; Moolten and Wells, 1990![]() ; Culver et al., 1992
; Culver et al., 1992![]() ; Freeman et al.,
1993
; Freeman et al.,
1993![]() ; Vile et al., 1994
; Vile et al., 1994![]() ; Smythe et al., 1995
; Smythe et al., 1995![]() ; Roth and Cristiano,
1997
; Roth and Cristiano,
1997![]() ). Bystander killing is thus
critical for the eradication of tumors (Roth and Cristiano, 1997
). Bystander killing is thus
critical for the eradication of tumors (Roth and Cristiano, 1997![]() ). However, suicide genes
are also used in clinical protocols of allogenic bone marrow
transplantation (Link et al., 2000
). However, suicide genes
are also used in clinical protocols of allogenic bone marrow
transplantation (Link et al., 2000![]() ). The HSV-1 TK gene can be
transfected ex vivo into donor T lymphocytes before their infusion
into patients. GCV can subsequently be administered to destroy the
allogenic T lymphocytes if graft-versus-host disease occurs. In this
case, bystander effect is probably unimportant and should even
be avoided.
). The HSV-1 TK gene can be
transfected ex vivo into donor T lymphocytes before their infusion
into patients. GCV can subsequently be administered to destroy the
allogenic T lymphocytes if graft-versus-host disease occurs. In this
case, bystander effect is probably unimportant and should even
be avoided.
In the present study, we investigated the bystander effect of BVDU in a Dm-dNK-transduced thymidine kinase-deficient osteosarcoma cell line and the MIA PaCa-2 human pancreatic adenocarcinoma cell line. We also studied the ability of hydroxyurea, a ribonucleotide reductase inhibitor, to enhance the cytotoxicity and the efficiency of bystander killing of Dm-dNK-transduced cells treated with BVDU.
| |
Materials and Methods |
|---|
Cell Culture and Retroviral Transduction. TK-deficient osteosarcoma
cells was a kind gift from Professor J. Balzarini (Rega Institute, Leuven,
Belgium). MIA PaCa-2 human pancreatic adenocarcinoma cells were
purchased from the American Type Culture Collection (Manassas, VA).
All cells were cultured in Dulbecco's modified Eagle's medium
supplemented with 10% (v/v) fetal calf serum (Invitrogen, Carlsbad,
CA), 100 U/ml penicillin, and 0.1 mg/ml streptomycin. The
cells were cultured at 37°C in a humidified incubator with a gas
phase of 5% CO2. The production of recombinant
replication-deficient retroviral vectors has been described in detail
previously (Zheng et al., 2000![]() ). The cells were transduced
with the retrovirus-containing medium mixed with 4 µg/ml
polybrene. The cells were incubated for 48 h and then cultured
continuously for 3 weeks in the presence of 1.0 mg/ml
Geneticin (Invitrogen).
). The cells were transduced
with the retrovirus-containing medium mixed with 4 µg/ml
polybrene. The cells were incubated for 48 h and then cultured
continuously for 3 weeks in the presence of 1.0 mg/ml
Geneticin (Invitrogen).
Generation of Mouse Polyclonal Antibodies and Western Blot Analysis.
Dm-dNK was expressed in the BL21 Escherichia coli strain with an
N-terminal polyhistidine tag and was purified by TALON resin affinity
chromatography (CLONTECH, Palo Alto, CA). Then, 2.0 µg of fusion
protein in 300 µl of phosphate-buffered saline was subcutaneously
injected into three 4-week-old female BALB/c mice with an equal
volume of Freund's complete adjuvant (Sigma, St. Louis, MO). This
was followed by a booster injection 10 days later of the same
amount of fusion protein in Freund's incomplete adjuvant (Sigma),
which was injected in the same manner. Two weeks after the booster
injection, 3 ml of blood was retrieved and allowed to clot. The
serum was collected and stored at  20°C.
20°C.
Cell Proliferation Assays. (E)-5-(2-Bromovinyl)-2'-deoxyuridine
(BVDU) was a gift from Professor J. Balzarini. The cells were plated at
 2000
cells/well in 96-well plates. BVDU with and without hydroxyurea
(120 µM) was added after 24 h, and the medium containing
the BVDU (with and without hydroxyurea) was changed once during the
experiment. Cell survival was assayed by a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
(Roche Molecular Biochemicals, Summerville, NJ) after 2 to
3 days of drug exposure. Each experiment was performed in
triplicate. The IC50 value of the investigated compounds was
calculated as the mean value of the experiments.
2000
cells/well in 96-well plates. BVDU with and without hydroxyurea
(120 µM) was added after 24 h, and the medium containing
the BVDU (with and without hydroxyurea) was changed once during the
experiment. Cell survival was assayed by a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay
(Roche Molecular Biochemicals, Summerville, NJ) after 2 to
3 days of drug exposure. Each experiment was performed in
triplicate. The IC50 value of the investigated compounds was
calculated as the mean value of the experiments.
Bystander Effect. The protocol used for the bystander experiments is
similar to protocols described previously (Denning and Pitts, 1997![]() ; Qiao et al.,
2000
; Qiao et al.,
2000![]() ). Tumor cells expressing
Dm-dNK were mixed at different ratios with their respective
parental cell lines. To promote cell contacts, the mixed cells were
plated in 24-well plates at 3 ×105 cells/well. The following
day, confluent cells were treated with BVDU ranging from
0.001 to 100 µM. After another 24-h incubation, cells were
trypsinized and a 1:100 dilution of the cells was distributed into
96-well plates in five replicates. Cells were cultured subsequently
in the presence of BVDU for 2 to 3 days until cells without
BVDU reached confluency. The proliferation of the cells was
measured by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
assay. We calculated the inhibition of bystander cells
proliferation using a method described previously (Qiao et al.,
2000
). Tumor cells expressing
Dm-dNK were mixed at different ratios with their respective
parental cell lines. To promote cell contacts, the mixed cells were
plated in 24-well plates at 3 ×105 cells/well. The following
day, confluent cells were treated with BVDU ranging from
0.001 to 100 µM. After another 24-h incubation, cells were
trypsinized and a 1:100 dilution of the cells was distributed into
96-well plates in five replicates. Cells were cultured subsequently
in the presence of BVDU for 2 to 3 days until cells without
BVDU reached confluency. The proliferation of the cells was
measured by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
assay. We calculated the inhibition of bystander cells
proliferation using a method described previously (Qiao et al.,
2000![]() ). In the cultures
containing A% dNK-expressing cells, the relative cell
proliferation of Dm-dNK-expressing cells (Dm-dNK cell
proliferation × A) and untransfected cells
[(untransfected cell proliferation × (100 A)] was subtracted
from the relative proliferation of the cultures containing mixtures
of Dm-dNK-expressing cells and untransfected cells [cell
mixture proliferation A × (Dm-dNK cell
proliferation) (100 A) × (untransfected cell proliferation)].
The inhibition of bystander cell proliferation was expressed as
a percentage relative to cells cultured without BVDU; i.e., 50% reflects a 50%
decrease in proliferation of bystander cells compared with the
inhibition of proliferation detected in the cells in which no
bystander effect occurred.
). In the cultures
containing A% dNK-expressing cells, the relative cell
proliferation of Dm-dNK-expressing cells (Dm-dNK cell
proliferation × A) and untransfected cells
[(untransfected cell proliferation × (100 A)] was subtracted
from the relative proliferation of the cultures containing mixtures
of Dm-dNK-expressing cells and untransfected cells [cell
mixture proliferation A × (Dm-dNK cell
proliferation) (100 A) × (untransfected cell proliferation)].
The inhibition of bystander cell proliferation was expressed as
a percentage relative to cells cultured without BVDU; i.e., 50% reflects a 50%
decrease in proliferation of bystander cells compared with the
inhibition of proliferation detected in the cells in which no
bystander effect occurred.
| |
Results |
|---|
Expression of Dm-dNK in Cancer Cells. A thymidine kinase-1
(TK1)-deficient human osteosarcoma cell line and a MIA PaCa-2 human pancreatic
adenocarcinoma cell line were transduced with replication-deficient
recombinant retrovirus with and without the Dm-dNK cDNA. A
polyclonal population of stably transduced cells was obtained by
Geneticin selection. Western blot analysis with mouse polyclonal
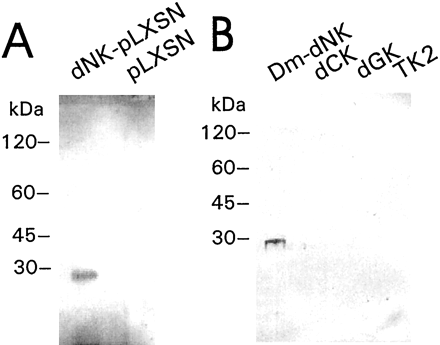
anti-Dm-dNK antibodies detected a band of ~28 kDa in the cells
transduced with Dm-dNK, but not in the cells transduced with
the control vector (Fig. 1A).
The specificity of the antibodies was verified using a Western
blot analysis on recombinant Dm-dNK and the sequence-related
human deoxyribonucleoside kinases deoxycytidine kinase,
deoxyguanosine kinase, and thymidine kinase-2 (Fig. 1B).
The anti-Dm-dNK antibodies detected the Dm-dNK protein
and did not cross-react with the human nucleoside kinases. To verify
that the Dm-dNK retained its enzymatic activity when expressed
in human cells, we determined the activity of thymidine
phosphorylation in crude cell protein extracts. Compared with their
parent untransduced cell line, the human osteosarcoma cells and the
pancreatic adenocarcinoma cells expressing Dm-dNK showed 100-fold and 30-fold increases in
thymidine kinase activity, respectively (Zheng et al., 2000![]() ).
).
|
Bystander Effect of BVDU in Dm-dNK Expressing Cells. Cells
expressing Dm-dNK (Dm-dNK+) were mixed at indicated
ratios with cells from their respective parental cell line
(Dm-dNK.gif) ). To
promote cell contacts, the mixed cells were initially plated in
24-well plates. The next day, BVDU was added to the confluent cells
at concentrations ranging from 0.001 to 100 µM. After another
24-h incubation, the cells were trypsinized and a dilution of
the cells was distributed into 96-well plates; exposure to BVDU
was continued for another 2 to 3 days. A bystander effect
was found in the osteosarcoma cell line (Fig. 2A),
but not in the MIA PaCa-2 cells (Fig. 2B).
At BVDU concentrations of 1 µM or higher, a bystander effect was
seen at all investigated different mixtures of
Dm-dNK+ and Dm-dNK
osteosarcoma cells, whereas no bystander effect was seen at lower
concentrations of BVDU.
). To
promote cell contacts, the mixed cells were initially plated in
24-well plates. The next day, BVDU was added to the confluent cells
at concentrations ranging from 0.001 to 100 µM. After another
24-h incubation, the cells were trypsinized and a dilution of
the cells was distributed into 96-well plates; exposure to BVDU
was continued for another 2 to 3 days. A bystander effect
was found in the osteosarcoma cell line (Fig. 2A),
but not in the MIA PaCa-2 cells (Fig. 2B).
At BVDU concentrations of 1 µM or higher, a bystander effect was
seen at all investigated different mixtures of
Dm-dNK+ and Dm-dNK
osteosarcoma cells, whereas no bystander effect was seen at lower
concentrations of BVDU.
|
Effects of Hydroxyurea on the Cytotoxicity of BVDU. The ribonucleotide
reductase inhibitor hydroxyurea has been shown to enhance cell killing and
bystander effects in cells transduced with HSV-1 TK and treated with
GCV (Boucher et al., 2000![]() ). To determine
whether hydroxyurea had an effect on the BVDU-mediated cell death, we
measured the sensitivity of cells expressing Dm-dNK in both
TK1-deficient osteosarcoma and wild-type MIA PaCa-2 human pancreatic
adenocarcinoma cell lines. Hydroxyurea alone was not toxic to the
cells up to concentrations of 150 µM (Fig. 3),
and we therefore decided to use 120 µM hydroxyurea in the
combination experiments. The IC50 value was 150-fold lower for the
Dm-dNK-expressing osteosarcoma cells incubated with BVDU in
the presence of 120 µM hydroxyurea than for cells incubated with
BVDU alone (Table 1). In
the MIA PaCa-2 cell line, the IC50 value for BVDU was
50-fold lower in the presence of hydroxyurea.
). To determine
whether hydroxyurea had an effect on the BVDU-mediated cell death, we
measured the sensitivity of cells expressing Dm-dNK in both
TK1-deficient osteosarcoma and wild-type MIA PaCa-2 human pancreatic
adenocarcinoma cell lines. Hydroxyurea alone was not toxic to the
cells up to concentrations of 150 µM (Fig. 3),
and we therefore decided to use 120 µM hydroxyurea in the
combination experiments. The IC50 value was 150-fold lower for the
Dm-dNK-expressing osteosarcoma cells incubated with BVDU in
the presence of 120 µM hydroxyurea than for cells incubated with
BVDU alone (Table 1). In
the MIA PaCa-2 cell line, the IC50 value for BVDU was
50-fold lower in the presence of hydroxyurea.
|
|
Effect of Hydroxyurea on the Bystander Effect of BVDU. Mixtures of
Dm-dNK- and
Dm-dNK+-transduced cells, with and without 120 µM
hydroxyurea, were cultured with different concentrations of BVDU to
study the bystander effect. The osteosarcoma cells showed a bystander
effect that gradually increased with higher doses of BVDU. The
bystander effect was more pronounced in the presence of hydroxyurea,
with a highly significant difference compared with the bystander
effect of BVDU in cell cultures without hydroxyurea (Table 2). We
found that 10% Dm-dNK-expressing cells induced cell death in
the presence of hydroxyurea to a degree similar to that obtained
obtained with 50% Dm-dNK-expressing cells without hydroxyurea.
For the MIA PaCa-2 cell line, there was no bystander effect in either
the absence or the presence of hydroxyurea (data not shown).
|
| |
Discussion |
|---|
In this study, we investigated the cytotoxicity and the bystander effect of a
novel suicide gene/prodrug combination: Dm-dNK and BVDU. Our
data demonstrate a BVDU-mediated bystander cell killing in
Dm-dNK-transduced osteosarcoma cells that could be enhanced by
subtoxic concentrations of hydroxyurea. In a Dm-dNK-transduced
human pancreatic adenocarcinoma cell line, MIA PaCa-2, we showed
that hydroxyurea increased the cytotoxicity of BVDU, but a bystander
effect was not detected in these cells. In a previous study, we
demonstrated that BVDU was an efficient substrate for both HSV-1
TK and Dm-dNK (Johansson et al., 1999![]() ), and the compound has
previously been investigated as a prodrug in HSV-1 TK-transduced
cells. Although BVDU did not show any bystander effect in HSV-1
TK-transduced osteosarcoma cells (Degreve et al., 1999
), and the compound has
previously been investigated as a prodrug in HSV-1 TK-transduced
cells. Although BVDU did not show any bystander effect in HSV-1
TK-transduced osteosarcoma cells (Degreve et al., 1999![]() ), we detected a clear
bystander effect in the Dm-dNK-transduced osteosarcoma cells
in the present study. The high catalytic rate of Dm-dNK is one
possible mechanism to explain the differences in bystander effects
for BVDU when activated by different enzymes in the same cell
line. However, the bystander effect of BVDU in our study is not as
pronounced as the bystander effect of GCV in HSV-1 TK-transduced
cells, and GCV remains unique among the nucleoside analog prodrugs
regarding its high efficiency in bystander cell killing. No bystander
killing was observed for BVDU in the MIA PaCa-2 pancreatic
adenocarcinoma cell line transduced with Dm-dNK. This cell
line has previously been shown to exhibit poor bystander killing when
using HSV-1 TK/GCV. The MIA PaCa-2 cell line has also shown very low
levels of expression of mRNA for both connexin-43 and connexin-26,
which are the main proteins of gap junctions (Yang et al., 1998
), we detected a clear
bystander effect in the Dm-dNK-transduced osteosarcoma cells
in the present study. The high catalytic rate of Dm-dNK is one
possible mechanism to explain the differences in bystander effects
for BVDU when activated by different enzymes in the same cell
line. However, the bystander effect of BVDU in our study is not as
pronounced as the bystander effect of GCV in HSV-1 TK-transduced
cells, and GCV remains unique among the nucleoside analog prodrugs
regarding its high efficiency in bystander cell killing. No bystander
killing was observed for BVDU in the MIA PaCa-2 pancreatic
adenocarcinoma cell line transduced with Dm-dNK. This cell
line has previously been shown to exhibit poor bystander killing when
using HSV-1 TK/GCV. The MIA PaCa-2 cell line has also shown very low
levels of expression of mRNA for both connexin-43 and connexin-26,
which are the main proteins of gap junctions (Yang et al., 1998![]() ).
).
Several mechanisms may be responsible for bystander effects in vivo, and at
least two mechanisms have been shown to mediate bystander killing in
HSV-1 TK-transduced cells in vitro (Aghi et al., 2000![]() ). These include the transfer
of phosphorylated GCV through intercellular gap junctions and the
phagocytosis of apoptotic vesicles containing GCV metabolites from
dying HSV-1 TK-transduced cells (Bi et al., 1993
). These include the transfer
of phosphorylated GCV through intercellular gap junctions and the
phagocytosis of apoptotic vesicles containing GCV metabolites from
dying HSV-1 TK-transduced cells (Bi et al., 1993![]() ; Freeman et al., 1993
; Freeman et al., 1993![]() ; Seachrist, 1994
; Seachrist, 1994![]() ; Fick et al.,
1995
; Fick et al.,
1995![]() ). The importance of gap
junctions is supported by studies demonstrating that the bystander
effect correlates with the extent of gap junctions between cells
(Fick et al., 1995
). The importance of gap
junctions is supported by studies demonstrating that the bystander
effect correlates with the extent of gap junctions between cells
(Fick et al., 1995![]() ) and the fact that
neuroblastoma and pheochromocytoma cells that lack endogenous
junctional conductance show no bystander effect unless they are
transfected with connexin genes (Vrionis et al., 1997
) and the fact that
neuroblastoma and pheochromocytoma cells that lack endogenous
junctional conductance show no bystander effect unless they are
transfected with connexin genes (Vrionis et al., 1997![]() ). Also, drugs that up-regulate
gap junctions, such as retinoic acid (Park et al., 1997
). Also, drugs that up-regulate
gap junctions, such as retinoic acid (Park et al., 1997![]() ), augment the HSV-1 TK/GCV
bystander effect. Dieldrin, which decreases gap junctions, reduces
this effect (Touraine et al., 1998
), augment the HSV-1 TK/GCV
bystander effect. Dieldrin, which decreases gap junctions, reduces
this effect (Touraine et al., 1998![]() ). The role of gap junctions
for the in vivo bystander effect has been verified through the
transfection of rat glioma cells with connexin-43 cDNA. The
experimental tumors could be completely eliminated when only 25% of
the cells expressed HSV-1 TK, whereas tumor cells expressing low
endogenous levels of connexin-43 could not be eliminated even when
50% of the cells expressed HSV-1 TK (Dilber et al., 1997
). The role of gap junctions
for the in vivo bystander effect has been verified through the
transfection of rat glioma cells with connexin-43 cDNA. The
experimental tumors could be completely eliminated when only 25% of
the cells expressed HSV-1 TK, whereas tumor cells expressing low
endogenous levels of connexin-43 could not be eliminated even when
50% of the cells expressed HSV-1 TK (Dilber et al., 1997![]() ).
).
Hydroxyurea increases the GCV sensitivity of cells expressing HSV-1 TK
(Boucher et al., 2000![]() ). Hydroxyurea reduces the
intracellular dGTP level, and the enhancement of GCV-mediated
toxicity was suggested to be the result of an increased GCV-TP/dGTP
ratio. We found a marked increase in cytotoxicity of BVDU, similar to
that of GCV, when the compound was administered in combination with
subtoxic concentrations of hydroxyurea. Although the total dTTP pool
increases in cells incubated with hydroxyurea (Collins and Oates,
1987
). Hydroxyurea reduces the
intracellular dGTP level, and the enhancement of GCV-mediated
toxicity was suggested to be the result of an increased GCV-TP/dGTP
ratio. We found a marked increase in cytotoxicity of BVDU, similar to
that of GCV, when the compound was administered in combination with
subtoxic concentrations of hydroxyurea. Although the total dTTP pool
increases in cells incubated with hydroxyurea (Collins and Oates,
1987![]() ), the enhanced
sensitivity to pyrimidine analogs is probably a result of a decrease
in the de novo dTTP synthesis, which could favor the salvage of
deoxyribonucleosides and nucleoside analogs. It is presently not
known how the expression of Dm-dNK influences the dNTP pools
in cells. This enzyme is unique in its efficient phosphorylation of
all four natural deoxyribonucleosides, and future studies should
reveal how such a highly active enzyme affects the dNTP pools and the
supply of DNA precursors.
), the enhanced
sensitivity to pyrimidine analogs is probably a result of a decrease
in the de novo dTTP synthesis, which could favor the salvage of
deoxyribonucleosides and nucleoside analogs. It is presently not
known how the expression of Dm-dNK influences the dNTP pools
in cells. This enzyme is unique in its efficient phosphorylation of
all four natural deoxyribonucleosides, and future studies should
reveal how such a highly active enzyme affects the dNTP pools and the
supply of DNA precursors.
Dm-dNK has a broad substrate specificity, and several cytotoxic compounds have been identified as substrates for the enzyme. Our study suggests that the Dm-dNK/BVDU combination should be further evaluated in applications in which a bystander effect is not desired (i.e., in allogenic bone marrow transplantation and in other cell therapy protocols). For these applications, the inability of Dm-dNK to activate GCV or acyclovir should be an advantage because these patients often have herpes virus infections. For the treatment of solid tumors, other substrates for Dm-dNK should be evaluated that may have more efficient bystander cell killing. It is important to realize that studies of bystander effects performed in vitro may be of limited value to predict the in vivo situation. However, they are an important step toward a better understanding of the basic mechanisms of prodrug activation by suicide genes and for the development of novel therapeutic suicide gene/prodrug combinations.
| |
Footnotes |
|---|
Received December 21, 2000; Accepted May 1, 2001
This work was supported by grants from the Swedish Medical Research Council, the Swedish Cancer Foundation, and the European Community.
Anna Karlsson, Division of Clinical Virology, F68, Karolinska Institute, Huddinge University Hospital, S-141 86 Stockholm, Sweden. E-mail: [email protected]
| |
Abbreviations |
|---|
BVDU, (E)-5-(2-bromovinyl)-2'-deoxyuridine; Dm-dNK, Drosophila melanogaster deoxyribonucleoside kinase; HSV-1 TK, herpes simplex virus type-1 thymidine kinase; GCV, ganciclovir; TBS, Tris-buffered saline.
| |
References |
|---|
|
|
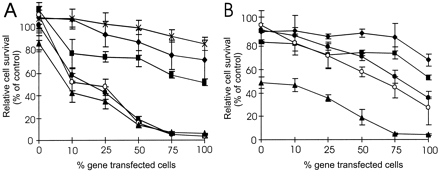
 ),
0.1 (
),
0.1 ( ), 1 (
), 1 ( ),
10 (
),
10 ( ), and 100 µM
(
), and 100 µM
( ). Data are expressed as a proliferation percentage
relative to the proliferation of cells in the absence of BVDU and
are the average of five values ± S.D.
). Data are expressed as a proliferation percentage
relative to the proliferation of cells in the absence of BVDU and
are the average of five values ± S.D. 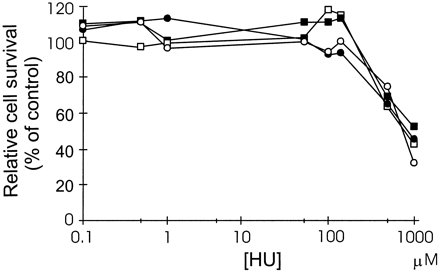
 ).
).