Effects of Rock Temperature on Limpet Densities
E. T. Kosman
Abstract:
Temperature plays an important role in the zonation of the intertidal community. To date, very few studies have examined the role of substratum temperature in zonation. Rock temperature and species density was recorded to determine if there was such an effect. This study found no daily effect of substratum temperature on zonation, but found a possible seasonal effect on zonation.
Introduction:
Zonation in the intertidal has been attributed by two causes, physical stresses and predation. Tolerance to physical stress is believed to set the upper limit of the distribution of a species. These physical stresses are desiccation, osmotic stress, oxygen deprivation and temperature stress (Somero, 2002).
In particular, resistance to temperature stress has been shown to be correlated with an increasing tidal height range (Somero, 2002). Temperature stress has often been linked with the growth rate of mollusks, showing an important sub-lethal impact on species (Kenny, 1983). While studies have been conducted on lethal temperatures of intertidal species (Miller, 2005) and the roles that temperature plays in zonation (Bertness et al, 1999), these studies have used ambient temperatures in their calculations. Few studies have focused on the role that microhabitat temperatures may play in the distribution of intertidal invertebrates.
Helmuth (2002) stated that the color of the shell and how much contact it had with the substratum help dictate the animal’s internal temperature. Thus, the temperature of the microhabitat may play an important but relatively undocumented role in species distribution. In a study done on barnacles, it was noted that by artificially shading the rocks the temperature of the substratum significantly increased survivorship and decreased the temperature of the rock (Bertness et al, 1999).
Another set of
intertidal species that have a high surface area in contact with the substratum
are limpets. Limpets are found in all
intertidal zones and have known lethal temperatures which determine their
placement in the intertidal (Wollcott, 1973).
Previous work on limpets has shown that the internal body temperature of
the animal in equal to that of its substratum (Wollcott, 1973). Four species of limpets common off the coast
of
The purpose of this study is to determine if there is a difference in the densities of limpet species based on the temperature of the substratum. By examining the effects of the rock temperature on limpet density, I hope to determine the importance of the microhabitat in zonation studies.
Methods:
Two sites were
selected, one at
Adults were identified as one of four limpet species (M. scabra, L. digitalis, L. limatula, or L. pelta) and counted. Sub-adults and other species were counted, but not identified. Limpets were counted on two separate days; September 18 and November 13 in 2005. Limpets were counted when mean low tide was less than +0.4, and no earlier in the day than 12:30 p.m.(Pacific Standard Time). Rocks temperature was measured within the quadrat using a Raytek ST Pro Non-Contact Thermometer.
One-way ANOVAs were performed to determine if total limpet densities varied by date or by site. One-way ANOVAs were also used to determine if there was a significant interaction between temperature and date or site. A two-way ANOVA was used to determine if there was any significant interaction between site and the total densities of each limpet species. As number of rock sampled for each date was different, a GLM was performed to determine if there was an interaction between the total density of each limpet species and sampling date. A regression analysis was performed for each species to determine if temperature of the rock affected the density of the species.
Results:
No L. pelta were sampled at either site, so
this species was excluded from the analysis. There was no significant
difference in the total number of limpets sampled by date (Fig. 1). However, there was a significant difference
in total numbers of limpets found between sites (Fig. 2), with more limpets
found at Wylder Annex than at
Fig. 1: There was no difference in
the number of limpets found, regardless of species, between the two
sampling dates (P=0.37, F=0.83, DF= 63). 10.15 is September 15th 11.13 is November 13th
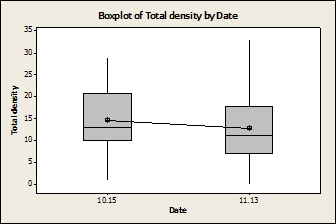
Fig. 2: There was a significant
difference in the number of limpets found, regardless of species, between
the two sampling sites (P=0.016, F=2.61, DF=63). 1 is Wylder Annex 2 is
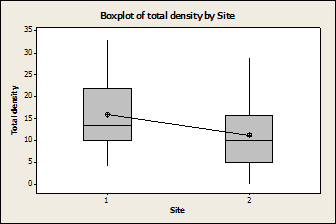
There was a significant effect on temperature due to site and date, with Wylder Annex having a higher temperature than Cabrillo (Fig. 3). This could be due to the fact that Cabrillo was sampled second, when the temperatures had begun to decline.
Fig. 3:
Temperature significantly different by site (P<0.001, F=16.33, DF=63). 1 is Wylder Annex 2 is
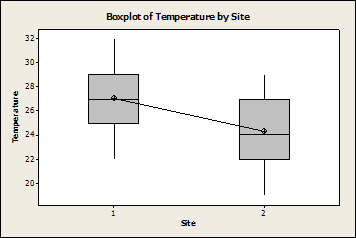
When the effect of site on temperature
was removed there was a significant effect date on temperature for Wylder Annex
(Fig. 4) with the temperature lower during November than in the September
sampling period, but no effect of temperature by date for
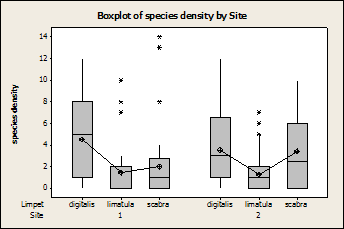
There was no
effect of site on the densities of each individual limpet species (Fig. 5),
even though total limpet densities were found to be lower at
|
|
|
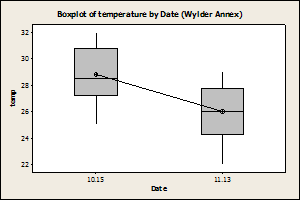
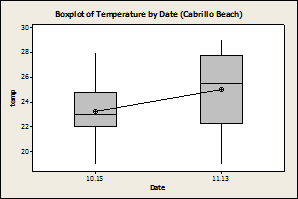
Fig. 4: Wylder
Annex showed a significant difference in rock temperature due to date (P=0.001,
F=15, DF=31), whereas Cabrillo beach did not (P=0.105, F=2.8, DF=31).
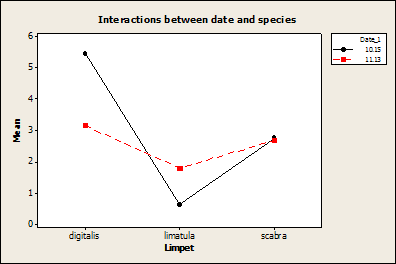
There was a significant effect of sampling date on individual species density. Less L. digitalis and more L. limatula were counted on November 13 as opposed to September 15 (Fig. 6).
Fig. 5:
Box plots showing the densities for each species by site. There was no significant difference
between the sites (P=0.076, F=2.61, DF=2). * are outliers 1 is Wylder Annex 2 is
Since there was a significant effect of temperature on sampling day and no effect of species density by site, samples were combined by site and not date for regression analysis. Regression analysis showed that there was no significant correlation between temperature of the rock and limpet species for any of the three limpet species for both days (Fig.7).
Fig. 6:
Shows the mean number of L. digitalis
found per rock decreased, while the mean number of L. limatula increased on November 13 sampling date (P=0.007,
F=5.10, DF=2).
|
C A A |
D B |
||||||||||
|
|
|
||||||||||
|
E |
F |
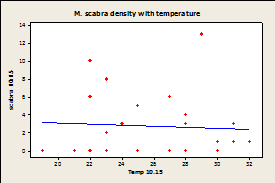
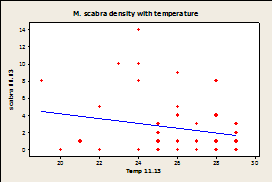
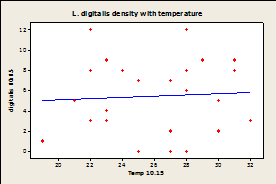
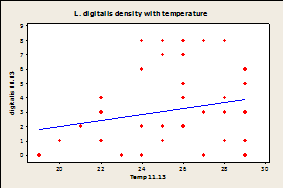
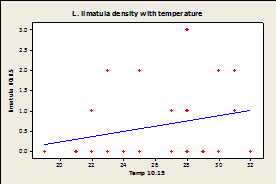
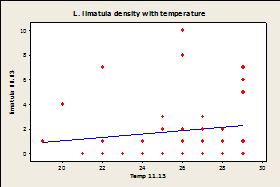
Fig.
7A-E: Scatter plot of
rock temperature and individual species density, with a line of best
fits. Regression analysis showed no
significant effect of rock temperature on species density after taking
sampling date into account (P>0.05).
R-squared values= 0.004 (A), 0.039 (B), 0.004 (C), 0.036 (D), 0.064
(E), 0.020 (F).
Discussion:
Although there was
no discernable effect of rock temperature on the distribution of limpet
species, this study shows a possible effect of seasonal temperatures on limpet
distribution. Daily rock temperature
seems to fluctuate considerably based on the weather conditions of the site, as
evidenced by
This cooling
trend may allow for migration upwards intertidally of the limpet species. Seasonal migrations in the intertidal have
been recorded for chitons (Harper and Williams, 2001) and limpets (Liu, 1994)
in
Breen (1972) noted that L. digitalis was able to move five meters or more away from its initial starting point in a month while migrating. Takada (1999) observed that mobile gastropods were able to evaluate their environment and move into shady areas within two weeks. Thus, it is very probable that L. limatula and L. digitalis were able to move from the study area within the time frame of the two sampling periods. Harper and Williams (2001) noted that chitons who were able to conform better to their crevice did not exhibit a migration pattern, as they were able to use their crevice as a refuge from heat. This may explain why there was no difference found in the densities of M. scabra, which resides in a home scar (Sept, 2002). While daily rock temperatures may not affect limpet densities, monthly and seasonal temperatures may.
Further studies are needed to validate this hypothesis. A semi-permanent transect across the different zones of the intertidal and monthly measurements of limpet densities by species may accomplish this. In addition, marking limpets and observing individual limpet movements would help us understand rates and direction of migration seasonally.
Literature Citied:
Bertness, M.D., G.H. Leonard, J.M.
Levine, J.F Bruno. 1999. Climate-driven interactions among rocky intertidal
organisms caught between a rock and a hot place. Oecologia. 120:446-450
Breen, P.A. 1972. Seasonal
migration and population regulation in the limpet Acmaea (Callisella) digitalis.
Veliger. 15:133-141
Harper, K.D., G.A. Williams. 2001. Variation in abundance and distribution
of the chiton
Acanthopleura japonica and associated molluscs on a seasonal, tropical,
rocky shore. Zoological Society of
Helmuth, B. 2002. How do we measure
the environment? Linking intertidal thermal
physiology and ecology through biophysics. Integrative and Comparative Biology.
42:837-845
Kenny, R. 1983. Growth
characteristics of intertidal limpets in relation to temperature trends.
Pacific Science. 37:37-44
Liu, J.H. 1994. The ecology of the
Miller, L.P. Effects of high
temperature and desiccation stress on the survival of the giant owl limpet, Lottia gigantea. Conference Society for
Integrative and Comparative Biology.
Somero, G.N. 2002. Thermal physiology
and vertical zonation of intertidal animals: optima, limits, and costs of living.
Integrative and Comparative Biology. 42:780-789
Sept, J.D. The Beachcomber’s Guide to Seashore life of
California. Harbor Publishing.
Takada, Y. 1999. Influence of shade and number of boulder layers on mobile organisms on a warm temperate boulder shore. Marine Ecology Progress Series. 189:171-179
Wolcott T.G. 1973. Physiological ecology and intertidal zonation in limpets (Acmaea): a critical look at "limiting factors". Biological Bulletin. 145:389-422