The effect of the amphipod Caprella californica on the forging behaviors and distribution of the nudibranch Hermissenda crassicornis in the mudflats of Bodega Bay and the role of size differences in the interspecific competition for space.
By
1Ellen Kosman
Bodega Marine Laboratory
University of California
Bodega Bay, California 94923
Environmental Science and Policy 124
Marine Coastal and Field Ecology
Summer 2002
Email address: 1[email protected]
Abstract:
Both Caprella californica and Hermissenda crassicornis occupy the eelgrass (Zostera marina) on the mudflats of Bodega Bay, CA. C. californica was found to have a significant impact on the feeding patterns of H. crassicornis. However, there is no literature on the interaction between these to species to date. My aim was to figure out how these to species interacted.
When C. californica was present in the field, H. crassicornis lost weight at a greater rate, than when C. californica was not present. In the laboratory, the presence of C. californica was linked with a longer transit time for H. crassicornis to get to Obelia a hydriod used as food. An increase in the size of C. californica was also linked with a longer transit time for H. crassicornis.
Introduction:
Both Caprella californica and Hermissenda crassicornis occupy the eelgrass (Zostera marina) on the mudflats of Bodega Bay, CA. C. californica can also be found in high densities on the hydroid Obelia sp. (pers. obs.), a food source for H. crassicornis (Hamiltion, 2001). However, in areas of high C. californica density, H. crassicornis are rare.
H. crassicornis is an aeolid nudibranch characterized by dorsal papillae (Morris et al., 1980). These papillae contain nematocysts and can secrete mucus as a defense mechanism (Thompson, 1960). Thompson (1960), Trowbridge (1994) and Lindgreen (2000) observed that when threatened H. crassicornis will wave the papillae and retract or hide all of their vulnerable parts, including the head and tail.
C. californica is an amphipod distinguished from other caprellids by a head spine (Morris et al., 1980) that may be used in defense (pers. obs.). They can cling to many structures, both man-made and natural, including eelgrass, docks, and Obelia sp. (Caine, 1979 & 1987). C. californica can move between different types of substrates, but prefer to live on the type of substrate on which they were born (Keith, 1971).
There is no literature on the interaction between these to species, despite the fact that they inhabitat the same areas. Few studies have been done on the ecology of caprellids. My aim is to further the knowledge of these two species by studying their interactions in different settings.
The following questions were posed for this research project: Does C. californica forcibly exclude H. crassicornis in areas of high caprellid density? Does the presence of C. californica on Obelia or eelgrass, affect H. crassicornis’ feeding behavior? Do different sizes of C. californica affect the foraging behavior of H. crassicornis?
Methods and materials:
All H. crassicornis were collected during low tide from the Bodega mudflats near channel marker 21 (Area 1), an area of high H. crassicornis density. C. californica were collected from three different sites: Spud Point Marina (Bodega Harbor), Area 1, and by channel marker 21 off the Westside Park campgrounds (Area 2), an area of low H. crassicornis density. Obelia sp. was collected from Spud Point Marina for use in laboratory experiments.
Field enclosures: effects of C. crassicornis on H. crassicornis growth
Forty-eight H. crassicornis were weighed and separated into 12 plastic containers, measuring 21 cm x 15.3 cm x 9.5 cm. The sides and top of the container were cut out and a fine mesh was glued over the holes using hot glue (fig. 1).
Fig. 1: A field container used to manipulate
desities of C. californica during studies of interspecific
competetion.
 Poles with flags were used in the field to
mark container location. A metal twist
tie was glued to one corner for attachment to the marking poles in the field. Four H. crassicornis were placed in
each container. Then, 12 to 13 grams of
eelgrass were placed in each container.
Twenty C. californica were added to six of the twelve
containers. Six control containers held
only H. crassicornis. On July 9,
2002, six containers (three controls and three experimental) were placed in
Area 1 (fig. 2), and six were placed in Area 2.
Poles with flags were used in the field to
mark container location. A metal twist
tie was glued to one corner for attachment to the marking poles in the field. Four H. crassicornis were placed in
each container. Then, 12 to 13 grams of
eelgrass were placed in each container.
Twenty C. californica were added to six of the twelve
containers. Six control containers held
only H. crassicornis. On July 9,
2002, six containers (three controls and three experimental) were placed in
Area 1 (fig. 2), and six were placed in Area 2.
Fig. 2: A picture of Area 1 with enclosures, by
channel marker 19.

I assumed that there were no differences between these two areas with respect to food concentration, predation rates, and other factors that may account for H. crassicornis’ absence in Area 2 aside from the high concentration of C. californica. A rock was placed in each container, to ensure that the container did not float away. After four days, the containers were collected and brought back to the lab and H. crassicornis weights were recorded.
Laboratory set-up: effects of C. californica on H. crassiconis’ access to food
 Laboratory
set up consisted of a rectangular plastic tray, measuring 38 cm x 8 cm x 5
cm. A strip of eelgrass was attached
using metal binder clips, 8 cm from the end of the dish.
Laboratory
set up consisted of a rectangular plastic tray, measuring 38 cm x 8 cm x 5
cm. A strip of eelgrass was attached
using metal binder clips, 8 cm from the end of the dish.
Fig 3: Laboratory set up. From bottom to top: H. crassicornis,
eelgrass, and Obelia. The
eelgrass was placed 8cm from the end, and the Obelia 8cm from that.
Because of high densities, C. californica was removed from the Obelia sp. using a freshwater dip. Obelia sp. was then rinsed with seawater and placed 8 cm from the eelgrass (fig. 3). Five C. californica were placed on the eelgrass for experimental trials and transit time of H. crassicornis to the Obelia sp. was recorded. No C. californica were on the eelgrass in control trials.
In between trials the trays were washed with Neutrad, an equimpment cleanser. Six different trials were run with four replicates of each trial. To study the effect of size on interspecific competition, C. californica and H. crassicornis were classified as either large or small. A large C. californica was > 200 mm, a small C. californica was <200 mm. A large H. crassicornis was > 250 mm, a small H. crassicornis was < 250 mm.
Results:
Field enclosure: effects of C. californica on H. crassicornis growth
There was no difference between the
average rate of weight change in H. crassicornis in the control
containers (no C. californica) from Area 1 and Area 2. A significant statistical difference in
weight loss was found between H crassicornis from 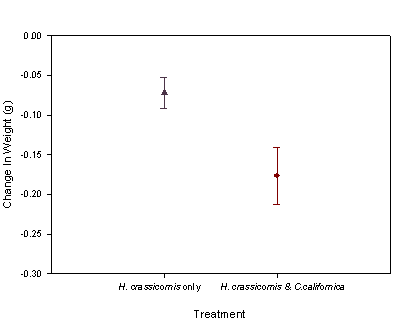 the
controls and H. crassicornis from the experimental containers (F= 6.864;
P= 0.028). When C. californica
was present, H. crassicornis lost weight an average of
0.180g. When C. californica was
not present, H. crassicornis lost weight an average of 0.072g (fig. 4).
the
controls and H. crassicornis from the experimental containers (F= 6.864;
P= 0.028). When C. californica
was present, H. crassicornis lost weight an average of
0.180g. When C. californica was
not present, H. crassicornis lost weight an average of 0.072g (fig. 4).
Fig. 4:
The average weight change of H. crassicornis with C.
californica, and without C. californica. H. crassicornis lost more wieght
when C. californica was present.
(Error bars are SD)
Laboratory set up: effects of C. californica on H. crassicornis’ access to food
I compared transit times for small H.
crassicornis to reach the Obelia when there were no C.
californica present to the times when there were large or small C.
californica present. Small H.
crassicornis had longer transit times when larger C. californica
were present (F= 2.898, P= 0.107). F
and P values were calculated from log-transformed data in order to pass
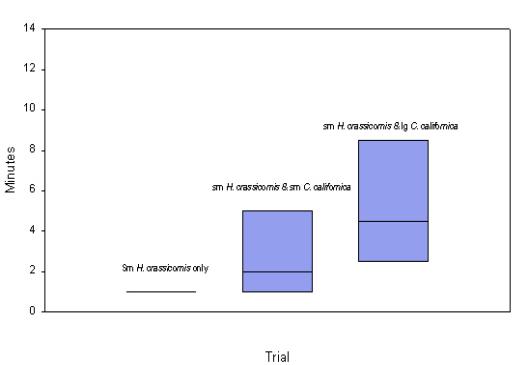
normality tests. Figure 5 shows an increase in
transit time as small H. crassicornis are exposed to small and
large C. californica.
|
Fig. 5: Shows an increase in transit time for small H. crassicornis
to reach Obelia, as size of C. californica increased. (ANOVA on
ranks) |
Three out of four small H. crassicornis were attacked by small C. californica. The small C. californica normally attacked only once, would release their hold on the eelgrass and move away from H. crassicornis. However, the female C. californica which were brooding young attacked more that once and did not move away from H. crassicornis. All small H. crassicornis were attacked when placed with large C. californica. Large C. californica attacked repeatedly and did not release their hold on the eelgrass.
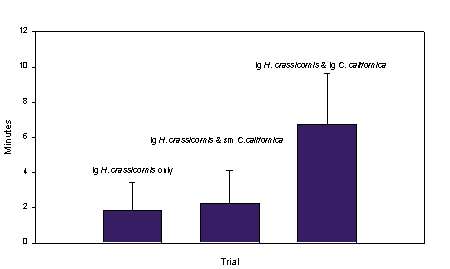
Large H. crassicornis results showed a significant difference in transit time when C. crassicornis was present (F= 6.197; P= 0.020). Large H. crassicornis had an increase in transit time when C. californica were present. (fig. 6).
|
Fig. 6: Shows
the increase in transit time for a large H. crassicornis to reach Obelia
sp. with increasing C. californica size. (Error bars are column means) |
Two out of four large H. crassicornis were attacked by small C. californica. Small C. californica tended to move away from large H. crassicornis. All the large H. crassicornis were attacked by the large C. californica. In two cases, large H. crassicornis turned towards the attacker and C. californica moved away. Otherwise, large C. californica remained on the eelgrass.
Discussion:
Mudflat sediments are able to hold water allowing for reduced physical stress to animals and thus the severe intertidal zonation characteristic of the rocky intertidal is reduced (Peterson, 1991). As a result, species interactions tend to play a larger role in the zonation of mudflat areas (Kafanov, 2001). The results of these experiments indicate that C. californica may play a role in the distribution of H. crassicornis. Control cages (containing only H. crassicornis) located in an area of high C. californica density and no H. crassicornis, did not lose any more wieght then the control cages in Area 1, were there where H. crassicornis. Conversely, in the experimental cages (containing both H. crassicornis and C. californica) H. crassicornis lost weight at a greater rate than the controls, in both areas. This suggests that C. californica may interfere interfering with H. crassicornis’ foraging behavior. While there may be other factors keeping H. crassicornis from Area 2, such as food scarcity, it is important to note that interference is occurring and may be a factor.
The experiments in the laboratory also confirmed this interference, both visually and graphically. In addition to the longer transit time, I observed numerous attacks on H. crassicornis by C. californica. The size of the C. californica seems to be an important factor in aggressive behavior. Smaller C. californica seem less aggressive based on the number and frequency of attacks, with the exception of the females brooding young. Larger C. californica were more aggressive, but would swim away from large H. crassicornis, if confronted. There are several interesting research possibilities suggested by these results and by other observations made during these experiments. I observed that during low tide, C. californica is less active than H. crassicornis. Could that be a reason why H. crassicornis is able to forage effectively in the areas where C. californica is present? Do they reproduce at different so that the bulk of their populations do not interact?
In the beginning I collected C. californica from three different sources, but towards the end I used C. californica from only one source. When C. californica from different sources were mixed together, they acted more antagonistically towards each other. They would exhibit aggressive behavior, including throwing one or another off the eelgrass. However, when C. californica from the same source was used, less of that behavior was evident. Do the C. californica recognize each other? Are they less antagonistic towards each other if they are related? Are they less antagonistic to certain kinds of C. californica, such as females with young, juveniles, or non-competitive males? The answer to these questions can only be found through further study.
C. californica is a fascinating animal that remains, for the most part, a mystery. Through future research, we will be able to answer these and many other questions. Not only about C. californica, but about intra- and interspecific interaction in species.
Acknowledgements:
I’d like to thank Vic, Janet, Julia and Dino, for all of their help and insight over the time of the course. Also, a thanks to my personal editor, Christina, and to Margaret for helping me get my cages out to the field. Thank you to all of the members of the class with cars who chauffeured this poor car-less person around. Last, but not least, a special thanks to Michelle for telling me about this class. Thanks, Michelle.
Citations:
Caine, E.A., 1987,Potential effects of floating dock communities on a South Carolina estuary, Journal of Experimental Marine Biology and Ecology, 108: 83-91
Caine, E.A., 1979, Population structures of two species of caprellid Amphipods (crustecea), Journal of Experimental Marine Biology and Ecology, 40: 103-114
Haderlie, Eugene C., Donald P. Abbot, Robert H. Morris, 1980, Intertidal Invertebrates of California, Stanford University press, Stanford
Hamilton, Sarah, 2001, Foraging behavior in the nudibranch Hermissenda crassicornis: how intra specific aggression and defense mechanisms affect prey selection, Student report: ESP 124
Kafanov, A.I., P.A. Fedotov, 1997, Relation of demographic and production indices to zonal biogeographic characteristics of coastal amphipod species in the Bay of Pos’et (Japan Sea), Zhurnal Obshchei Biologii, 58: 102-120
Keith, Donald E, 1971, Substrate selection in caprellid amphipods of southern California, with emphasis on Caprella californica Stimpson and Caprella equilibra Say (amphipoda), Pacific Science, 25: 387-394
Lindgreen, Annie, summer 2000, The defensive role of nematocysts in the aeolid nudibranch Hermissenda crassicornis, Student report, ESP 124
Peterson, Charles H, 1991, Intertidal zonation of marine invertebrates in sand and mud, American Scientist, 79 n3: 236(14)
Thompson, T.E., 1960, Defensive adaptations in opisthobrachs, Journal of Marine Biological Association, 39: 123-134
Trowbridge, Cynthia D., 1994, Defensive responses and palatability of specialist herbivores: predation on NE pacific ascoglossan gastropods, Marine Ecology Progress Series, 105: 61-70