
|
|
Neurological dysfunction is a manifestation of several neurodegenerative and autoimmune diseases of the nervous system, such as Alzheimer's disease, Parkinson�s disease, multiple sclerosis (MS) and in HIV-1 associated encephalopathy. The pathophysiological mechanisms underlying the changes in neurological function are as yet unresolved, but may include neuroinflammatory responses to infectious and/or environmental, as well as endogenous factors. I am interested in identifying and characterizing the molecular and cellular pathways involved in the neuroinflammatory mechanisms and in advancing the understanding of neuroinflammation in the initiation and expansion of cellular injury and death in the neurodegeneration of the central nervous system (CNS) diseases. | References: [1] [2] [3] [3] [4] |
Viral Pathogenesis: The mouse hepatitis virus (MHV)induced demyelination mimics some of the pathological hallmarks of human disease multiple sclerosis (MS). My work in this area focused on genetic and molecular approaches in understanding the molecular basis of MHV induced neurotropism.� I sequenced several demyelinating and non demyelinating strains of MHV and mapped several possible genomic regions involved in demyelinating properties (Das Sarma et al., 2001 [1], Das Sarma et al., 2001 [2]).�The genetic approach using targeted RNA recombination demonstrated for first time that demyelination determinants map to the spike (S) gene of MHV (Das Sarma et al., 2000 [3]). The recombinant viruses containing the S gene derived from MHV-2 (non encephalitic, non demyelinating strain) in MHV-A59(encephalitic and demyelinating strain) background (Penn 98-1/2) exhibited acute stage encephalitis and chronic stage persistence in the brain and spinal cord although they were unable to induce demyelination. The recombination studies clearly demonstrated that spike gene is one of the major determinants of demyelination but the viral persistence and the encephalitogenic properties of MHV are controlled by other genes or in combination of genes. Additionally, persistence of viral RNA and encephalitis are insufficient to induce chronic stage demyelination, although it may be a prerequisite for the development of demyelination. This finding is important as it provides fundamental insights into indirect cellular mechanism at play. | 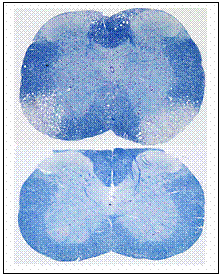 |
The alteration of virus spike protein influences the host interaction. Our understanding on how it happens is limited.� It is evident in the MHV induced disease model that replication and spread of viral antigen in the central nervous system (CNS) is a prerequisite for the development of chronic demyelination which develops after the resolution of encephalitis and clearance of the virus. It is unknown whether the spread is associated with the ability of the MHV strain to cause demyelination.� Comparing the spread of the viral antigen in the CNS between demyelinating strain and non demyelinating strain sheds some light in the understanding of the role of viral spread in the demyelination processes. To monitor the viral spread, I engineered both demyelinating and non demyelinating enhanced green fluorescent (EGFP) tagged MHV strains by targeted RNA recombination which are isogenic except for the S gene, and exhibiting pathogenic properties similar to their parental viruses. These EGFP expressing viruses are powerful tools to follow the viral spread using fluorescence over time without terminating the infection by fixation in order to allow detection of viral antigen by immunofluorescence (Das Sarma et al, 2002 [4]). | 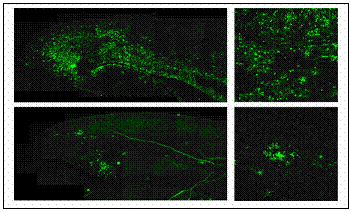 |
| My preliminary data suggests, that after intracerebral or intranasal inoculation, virus is first observed in the brain followed by spread into the spinal cord.� Viral antigen in the spinal cord is first detected in the gray matter, and then in white matter soon after. But very little is known about the mechanism of virus spread from gray matter into white matter and about the relationship between early viral replication and later induction of demyelination. It is possible that neuroglial communication orchestrates the pathological and cellular mechanisms of MHV induced chronic demyelination. Neurons and glial cells are known to communicate by direct coupling through gap junction, which provides pathways for direct intercellular communication.�Recent studies suggested that expression of gap junction protein is altered by CNS tissues in response to inflammation. | 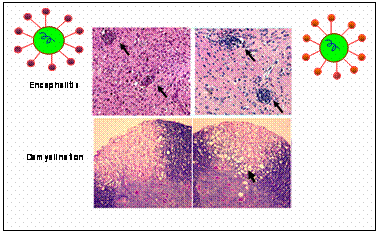 |

| Last updated on November 27, 2004. Jayasri Das Sarma, © Copy Rights Reserved. |