MICROPROPAGATION OF FERNS:
Pteris vittata spores and Nephrolepis biserrata runners
(An undergraduate thesis submitted to the Institute of Biology, College of Science, University of the Philippines in partial fulfillment for the B.S. Biology degree by Aimee Julianne Anico-Parr in March 2000)
ABSTRACT
Propagation of ferns in vivo and in vitro for the ornamental plant industry and for research has been done several times using spores or vegetative parts. Micropropagation of Pteris vittata spores and Nephrolepis biserrata runners using different media formulations are the subjects of this research. These ferns are easily propagated by conventional methods. Micropropagation of these ferns can be applied to produce ferns that are hard to propagate conventionally for the benefit of the ornamental plant industry and further research.
Germination of Pteris vittata spores and elongation of resulting sporophytes are best done with � strength MS, 125 mg/l NaH2PO4, 5 mg/l thiamine-HCl, 500 mg/l myo-inositol, 20g/l sucrose, 1 mg/l BAP, 0.1 mg/l NAA solidified with 8 g/l bacto-agar. The growth of Pteris vittata spores in � strength MS with 30 g/l sucrose solidified with 8 g/l bacto-agar indicates that this fern is not very sensitive to the presence or absence of growth regulators. Spontaneous appearance of callus in culture is observed. Sporophytes both appeared from callus and gametophyte cultures. Rooting of sporophytes is best at � strength RMS with 10 mg/l sucrose solidified by 8 g/l agar. Nephrolepis biserrata runner tips embedded into the medium with � strength MS and 30 g/l sucrose solidified with 8 g/l bacto-agar produced buds and shoots. Rooted plantlet from buds using this medium was observed within 63 days. Callus formation was prolific in � strength MS, 125 mg/l NaH2PO4, 5 mg/l thiamine-HCl, 500 mg/l myo-inositol, 20 g/l sucrose, 2 mg/l BAP, 1 mg/l NAA. Rooted plantlets from callus observed within 94 days in same medium with growth regulators adjusted to 1 mg/l BAP and 0.1 mg/l NAA.
INTRODUCTION
Some ornamental plants used by a number of garden centers and nurseries for landscaping in the Philippines are collected from the wild. Depletion of these species collected from the wild is highly probable if no alternative means of propagation is commercially useful for garden centers. Nurseries using conventional methods propagate other ornamental plants. These methods include stem cuttings, seed and spore sowings, suckers, grafting and division of rhizomes, rootstocks and tuberous roots. These methods only generate few plants and take a longer time to mature so commercial availability and marketability is greatly reduced. Tissue culture of ornamental plants will generate more plants from a single mother plant in a shorter period of time (Kyte and Kleyn 1996). This method ensures more stock plants and less collection from the wild.
Ferns have been used extensively in the ornamental plant industry for landscaping and indoor. The beauty of their gently sloping leaflets makes an ideal accent in any setting. Most ferns like the shade (Steiner 1986) so it becomes an important accent to the darker portions of home and garden. Other uses of fern are for human food, handicraft raw materials, fertilizer, non-nuclear fuel, pest control and medicine (Zamora and Co 1986). Ferns are conventionally propagated from spores (Bridgers 1958; Fliflet 1961; Kleinschmidt 1952; Knobloch 1960; MacFadden 1963), from division of outgrowths from vegetative parts like runners (Beck and Caponetti 1983; Loescher and Albrecht 1979; Richards et al. 1983) and buds from leaflets (White 1968). Ferns propagated from spores undergo two stages in their life cycle that are characteristic of these plants, the sporophyte and the gametophyte stages (Zamora 2000). The gametophyte stage of ferns in terms of its simple morphology is an ideal experimental material for most research on plant growth (Miller 1968). A fern propagated from vegetative outgrowths is identical to the mother plant from which it came from.
Pteris vittata (Figure 1) belongs to the family Pteridaceae and is commonly called Chinese brake or rusty brake and has medicinal purposes (Zamora 1986). It has a short rhizome with green stipes when young that turns brown when the fern gets older. Up to 36 pairs of pinnae can be observed in this fern with the middle as the longest. The sori (Figure 2) are continuous from near the base to the apex of the pinnae. It is terrestrial in habitat and is small under exposed conditions and large under shaded conditions (Zamora 1986; Zamora 2000). They are commonly seen growing on stones (Steiner 1986). Their usual means of reproduction is from spores.
Nephrolepis biserrata (Figure 3) belongs to the sword fern family (family Oleandraceae), propagate itself by creeping stolons (Figure 4) and make beautiful clumps in a fairly short time (Riffle 1998). They are commonly referred to as the bold sword fern (Madulid 1995) or broad sword fern (Morton 1958) with a terrestrial habitat and drooping leaves. It has a short rhizome and clustered fronds. The stipes are green when young and turns brown as the fern gets older. The sterile pinnae (up to 2 cm wide) is wider than the fertile ones (1.5 cm wide) (Zamora 2000). The pinna has no ear-like appendages (Steiner 1986). The sori are at the ends of veins (Madulid 1995; Zamora 2000). There are two forms available, a fertile one and one that appears to be always sterile (Morton 1958). Their usual means of reproduction is from stolons.
Pteris vittata has been cultured in vitro to determine physiological and morphogenetic changes of fern gametophytes derived from dark cultured callus using Nitch's medium (Kato 1963). In vitro differentiation of Pteris cretica callus to gametophytic and sporophytic tissues in the presence or absence of sucrose has also been studied (Bristow 1962). Albaum (1938) used Pteris longifolia and Pteris aquilina prothallium to study the effects of growth hormones and polarity.
Nephrolepis exaltata cv. Bostoniensis runner tissue is a popular research material grown in vitro to determine effects of temperature and light (Hvoslef-Eide 1990), mother plant condition (Hvoslef-Eide 1990, 1991, 1992) and for studies of structure and organogenesis (Loescher and Albrecht 1979; Richards et al. 1983). Beck and Caponetti (1983) used runner tissues of Nephrolepis falcata forma furcans to determine effects of growth regulators on in vitro shoot multiplication.
This study aims to propagate in vitro Pteris vittata using spores and Nephrolepis biserrata using runners. Slight manipulation of the components of the culture media (hormone, basal salt and sucrose concentrations) used by the previously cited works will be done to determine the optimum media for both ferns. Results for both ferns under study can be applied to other hard to conventionally propagate but commercially desirable ferns. This will ensure that the ornamental plant industry will have another way (tissue culture) of reproducing specific ferns besides the conventional methods mentioned earlier.
REVIEW OF RELATED LITERATURE
A. Life cycle of fern
Life cycle of a fern (Zamora 2000) involves a sporophyte (the "fern" itself) where spores are produced on undersides or margins of leaves. The spores (used in sexual propagation) germinate (turn green) and produce the heart shaped "prothallus". Both the female and male organs develop from the prothallus giving rise to the gametophyte stage. Fertilization occurs when the sperm from the antheridia swims through water to the archegonia (where the eggs are). A complete fern is formed in the prothallus, which withers away. A new sporophyte is formed (Bower 1963; Nayar 1960; Riffle 1998; Zamora et al. 1993).
B. In vitro propagation
Micropropagation produces a large number of plants from small pieces of a stock plant in relatively short periods of time (Street 1977). The shoot apical meristem is actively dividing (Gunckel 1965) and will generally form a callus (Yeoman and Macleod 1977) when used for tissue culture. The advantages of this technique over conventional methods of propagation (Villareal et al. 1989) include producing unlimited plant material, year round supply of plantlets, reduced space requirement, minimal attention during propagation, faster clonal development, easier plantlet transportation and discovery of new cultivars (Mikkelsen and Hasegawa 1981; Fitchet 1990). Excision of the meristem and regeneration of plantlets on amended media free plants from viruses and other disease causing pathogens (Slack and Tufford 1995; Mikkelsen and Hasegawa 1981). One form of micropropagation, embryo culture is significant in that it overcomes seed sterility, rescuing incompatible hybrid crosses and in vitro regeneration (Hu and Zanettini 1995). Plants produced from tissue culture were proven to grow faster (Mikkelsen and Hasegawa 1981; Zamora et al. 1989) and therefore increase productivity.
Care should be taken when preparing medium and sterilizing explants. Oversterilizing the explants will kill the cells and tissues (which turn brown or white) (Gamborg 1995). Media contamination is observed as a milky white suspension in liquid media (from yeast), mycelia and round balls in liquid media (from fungus) and odor, obvious growth on agar or turbidity in liquid media (from bacteria) (Gamborg 1995). Murashige (1974) states three stages of tissue culture namely, establishment of the aseptic culture with numerous growths and no infection (Stage I), multiplication of the propagule (Stage II) and re-establishment of plants in soil (Stage III). Kyte and Kleyn (1996) refer to four stages namely explant establishment and initiation (Stage I), multiplication (Stage II), rooting (Stage III) and acclimatization or hardening off (Stage IV). Growth regulators (auxin and cytokinin) added to the MS medium will ensure shoot and root initiation depending on its concentrations (Zamora 1992). Cytokinins affect the differentiation of plant cells and influence cell proliferation (Szweykowska 1974) while auxins influence cell enlargement and root initiation (Kyte and Kleyn 1996). The optimum concentrations of each component of the media should give rise to increased productivity (or plantlet formation) (Dougall 1981). Subculturing of the culture to a fresh medium will provide fresh nutrients, preclude desiccation of agar and prevent metabolites from accumulating to toxic levels (Zamora 1992). The problem encountered in tissue culture is the low survival rate of the plantlet in the field and morphological variability (Philips et al. 1995; Mikkelsen and Hasegawa 1981). Commercial propagation of many ornamentals through tissue culture has been very successful (Mikkelsen and Hasegawa 1981).
Spores are studied (Foster 1956; Griswold 1956) and used in taxonomical purposes in ferns (Brown 1960). Spores are used in sterile culture or in more conventional methods (Bridgers 1958; Fliflet 1961; Kleinschmidt 1952; Knobloch 1960; MacFadden 1963,) of fern propagation. Growing ferns from spores involves the collection of spores from the sporophyte, sterilization of spores (Basile 1973; Bridgers 1958; Kato 1963; Kleinschmidt 1957; Knobloch 1957; Warne et al. 1986) and placing it into culture media. In vitro culture of different ferns uses different media other than MS (Murashige and Skoog 1962; Murashige 1974). Knudson C has been used extensively in fern in vitro culture (DeMaggio and Wetmore 1961; Knudson 1925; Knudson 1940; Marengo 1979; Morel and Wetmore 1951; Steeves 1952; Whittier 1981; Wetmore 1953). Other media used is Moore�s medium (Sheffield et al. 1997) and a commercial nutrient plant tablet called �Black Magic� (Marengo 1979). Fern gametophytes are quite popular research material due to its morphological simplicity of a higher plant (Miller 1968 Partenan 1955; Sheffield and Bell 1987; Swami and Raghavan 1980).
Vegetative parts of a fern have been used to propagate some species. Nephrolepis exaltata cv. Bostoniensis runner tissue is a popular research material grown in vitro to determine effects of temperature and light (Hvoslef-Eide 1990), mother plant condition (Hvoslef-Eide 1990, 1991, 1992) and for studies of structure and organogenesis (Loescher and Albrecht 1979, Richards et al. 1983). Beck and Caponetti (1983) used runner tissues of Nephrolepis falcata forma furcans to determine effects of growth regulators on in vitro shoot multiplication. Other vegetative parts of a fern that have been studied for propagation were leaf buds (White 1968). In vitro culture of leaf tissue using different media concentration has been done to study growth and development (Sussex and Steeves 1953, Steeves and Sussex 1957) and differentiation to gametophytes or sporophytes (Hirsch 1975). Oki (1981) discussed some modifications of research procedures done on Boston ferns for commercial applications, mainly in the equipment (media dispenser and culture vessels) used in the laboratory, actual culture techniques (liquid media and �chopping�) and transplanting methods.
MATERIALS AND METHODS
Source of explants
- Nephrolepis biserrata
Nephrolepis biserrata stock plant has been growing in the student�s garden for morethan two (2) years without showing any signs of fertile leaves (with spores) but plenty of runner growth. Two (2) runners measuring 10 cm long were collected and used in the experiment to determine the best part of the runner to grow in vitro. Several 3-cm long runner tips (Figure 5) were collected for other experiments. Runners were placed in plastic and submerged in water with ice cubes for transport to the laboratory.
- Pteris vittata
Pteris vittata grew on stones in the student�s garden and was transplanted in a shaded place. Fertile leaf (with brown sori) were collected and placed in white letter envelope and brought to the laboratory. The letter envelope was tapped and the brown powder was collected. Some of the sori were scraped off from under the pinnae and collected also.
Sterilization
- Nephrolepis biserrata
All the runners were cleaned by hand of any soil and debris attached to it and placed in a jar covered with gauze held in place by a rubber band. The jar was placed under running tap water for 30 minutes to 1 hour. Runners were placed in sterile jar with sterile distilled water. The rest of the sterilization process was done under the laminar flow hood (Figure 6). The results from these experiments were used for subsequent sterilizations. The following experiments were undertaken:
- Effects of different concentrations of sterilizing agents
Ten (10) pieces of 3-cm long runner tips were used to determine best sterilization procedure. Two (2) runners each were placed in jars and different concentrations of sterilizing agents were used. All the runners were dipped for 1 minute in 70% ethanol, washed with sterile water and then gently agitated in hypochlorite solution (brand Zonrox with 5.25% NaOCl content). The following hypochlorite concentrations 10%, 20% and 50% for 20 minutes and 20% and 50% for 10 minutes were used. A total of 5 sterilization treatments were made. Runners were washed at least 3 times with sterile distilled water. Runners were cut into 2 pieces. These were placed in media containing 2/3 strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.1 mg/l NAA, 20 g/l sucrose and 8 g/l agar (Kyte and Kleyn 1996) for viability and contamination studies. Four replicates per concentration were used.
- Effects of absence or presence of wetting agent
The result from the previous sterilization experiment was further studied by getting six (6) pieces of runner tips and placing two pieces each in jars. Runners were dipped for 1 minute in 70% alcohol then washed with sterile distilled water. Two runners each were placed in 20% hypochlorite solution with wetting agent [either 1 drop for every 100-ml solution of liquid detergent (brand Joy Ultra with anti-bac) or 1 drop Tween 20 for every 100-ml solution] or without wetting agent. Runners were washed at least 3 times with sterile distilled water. Runners were cut into 2 pieces of 15 mm each. These were placed into media containing 2/3 strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.1 mg/l NAA, 20 g/l sucrose and 8 g/l agar (Kyte and Kleyn 1996) for viability and contamination studies. Four replicates per wetting agent were used.
- Pteris vittata
The spores were washed with sterile distilled water and filtered through a silkscreen cloth (used stockings) to remove unwanted debris. The spores were placed in a sterile vial with sterile distilled water for at least 24 hours to synchronize opening of the spore capsules. The spores were placed in a sterile vial (inside the laminar flow hood) and 70% ethanol was added for 30 seconds and everything decanted in local filter paper. The remaining spores were washed from vial with alcohol. The spores were washed thoroughly with sterile distilled water while being filtered through the paper (local filter paper). The spores were placed back in the vial with 20% household bleach (5.25% NaOCl content) with 1 drop of tween 20 and agitated for 20 minutes (concentration of sterilizing agents from sterilization experiment shown in Table 1 was used). The spore suspension was decanted on new filter paper and washed thoroughly with sterile distilled water for removal of remaining bleach solution. The spores were placed back in vial with small (5 ml) amount of sterile distilled water.
Treatment used
Stock solutions of Murashige and Skoog medium (MS) (Murashige and Skoog 1962) and other chemicals were prepared and stored in the refrigerator while not in use. Media were prepared based on the treatments to be done and adjusted to pH 5.7. Agar at 8 g/l was added to each medium and melted over a hot water bath. Media were poured into culture vessels, 15-20 ml for 25 x 150 mm test tubes topped with cotton plugs (Figure 7), 30-40 ml for culture jars topped with aluminum foil and paper held in place by rubber bands (Figure 8) and 30-40 ml for petri dishes. The treatments A and B only used one media by Kyte and Kleyn (1996). Treatments C to E used media with different concentrations of chemicals (stated per treatment).
- Effect of runner part
The two pieces of previously sterilized (sterilization done using the results of previous experiment) 10-cm long Nephrolepis runner was cut up into 15-mm pieces under the laminar flow hood (Figure 5). Cut pieces were runner segment with node, runner segment with internode, runner segment 15 mm before the tip and the 15 mm runner segment with the tip itself. These were embedded into medium containing 2/3 strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.1 mg/l NAA, 20 g/l sucrose with pH adjusted to 5.7 then solidified with agar (Kyte and Kleyn 1996). Four parts and three replicates per part were made.
- Effect of vertical or horizontal position
The damaged portions from the sterilized 3-mm runner pieces were cut and only the tips were used for this experiment. The tips were placed in one type of media containing 2/3 strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.1 mg/l NAA, 20 g/l sucrose (Kyte and Kleyn 1996). Tips were placed in the agar in the following positions, vertical tip upwards, vertical tip downwards (Figure 9) and horizontal. Three positions and five replicates per position were made.
- Effect of different MS salt concentrations
The following media were prepared for this experiment:
- � strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.1 mg/l NAA, 20 g/l sucrose
- 2/3 strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.1 mg/l NAA, 20 g/l sucrose
- Full strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.1 mg/l NAA, 20 g/l sucrose
- Nephrolepis biserrata
The damaged portions from the 3-mm sterilized runners were cut and the hairs removed. The tips were placed in a vertical downward position into the agar medium. Three (3) different media with five replicates each were used.
- Pteris vittata
Five drops (approximately 0.25 ml) of spore suspension were inoculated into the petri dish with the solid media. Petri dishes were sealed with parafilm. Three (3) different media with three replicates each were done.
- Effect of different concentrations of growth regulators
The following media were prepared for this experiment:
- � strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 20 g/l sucrose (no growth regulators)
- � strength MS salts, 125 mg/l , NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.1 mg/l NAA, 20 g/l sucrose
- � strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 1 mg/l NAA, 20 g/l sucrose
- � strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 2 mg/l BAP, 1 mg/l NAA, 20 g/l sucrose
- � strength MS salts, 125 mg/l NaH2PO4, 500 mg/l myo-inositol, 5 mg/l thiamine-HCl, 1 mg/l BAP, 0.5 mg/l NAA, 20 g/l sucrose
- Nephrolepis biserrata
The damaged portions from the runners were cut and the hairs removed. The tips were placed in a vertical downward position into the agar medium. Five (5) different media with five replicates each were used.
Sporophytes from 91-day old cultures were subcultured to fresh new media in test tubes for measurement of frond and root length. Five (5) different media with two replicates each were used.
- Pteris vittata
Five drops (approximately 0.25 ml) of spore suspension were inoculated into the petri dish with the solid media. Petri dishes were sealed with parafilm. Five (5) different media with three (3) replicates per media were done.
Sporophytes from 77-day old gametophyte cultures were subcultured to fresh new media in test tubes for measurement of frond and root length. Five (5) different media with five replicates each were used. Sporophytes from 91-day old callus cultures were subcultured to fresh new media in test tubes for measurement of frond and root length. Five (5) different media with five replicates each were used.
- Effect of sucrose
The following media were made for this experiment.
- � strength MS salts, 10 g/l sucrose
- � strength MS salts, 30 g/l sucrose
- Nephrolepis biserrata
The damaged portions from the sterilized 3-mm runners were cut and the hairs removed. The tips were placed in a vertical downward position into the 2 media. Two (2) different media with five (5) replicates each were used.
Sporophytes from 91-day old cultures were subcultured to fresh new media in test tubes for measurement of frond and root length. Five (5) different media with five replicates each were used.
- Pteris vittata
Five drops (approximately 0.25 ml) of spore suspension were inoculated into the petri dish with the solid media. Petri dishes were sealed with parafilm. Two (2) different media with three (3) replicates per media were done.
Sporophytes from 77-day old gametophyte cultures were subcultured to fresh new media in test tubes for measurement of frond and root length. Five (5) different media with five replicates each were used. Sporophytes from 91-day old callus cultures were subcultured to fresh new media in test tubes for measurement of frond and root length. Five (5) different media with five replicates each were used.
Culture conditions and maintenance
All culture vessels were placed in a 3-tier growth chamber supplied with 3 white fluorescent lights per tier (Figure 6 and 10). Temperature in an air-conditioned room was maintained between 25-28oC with photoperiod of 12 hours in light and 12 hours in dark. Subculture of explants into fresh new medium was done every 3 weeks. Brownish calli were discarded. Clumps were divided and all parts that turned brown were cut.
Characters Measured
Nephrolepis biserrata
Cultures were observed weekly or every other week from the date of inoculation into media and all changes were noted. The following characters were measured:
- Response of runner explant
- Greening or browning of the runners: runners are typically green on the start of culture so maintenance of greening of the entire runner explant was observed with the naked eye
- Presence or absence of leafy shoots: leafy shoots were seen with a hand lens as green protrusions that resemble a fern frond about 1 mm in length.
- Presence or absence of buds: buds were observed with a hand lens as green (not as green as callus), rounded protrusions that resemble callus but with a whitish furlike surface.
- Presence or absence of callus: callus were observed with a hand lens as very green, rounded growth on the explant
- Presence or absence of roots: roots were seen with a hand lens as blackish hair like growth from the base of the sporophyte plant.
- Sporophyte development
- Frond length was measured while the sporophyte is still in the test tube (approximate) with the help of a hand lens and a ruler (in millimeter). The average was noted down as frond length.
- Root length was measured while the sporophyte is still in the test tube (approximate) with the help of a hand lens and a ruler (in millimeter). The average was noted down as root length.
- Numbers of fronds of the sporophyte inside the test tube were counted with the help of a hand lens.
- Numbers of roots of the sporophyte inside the test tube were counted with the help of a hand lens.
Pteris vittata
Cultures were observed weekly or every other week from the date of inoculation into media and all changes were noted. The following characters were measured:
- Response of spore culture
- Germination of spores as observed with a hand lens by greenish color
- Presence or absence of callus: callus was observed with a hand lens as slightly green, rounded growth on the plate
- Presence or absence of buds: buds on the callus were observed with a hand lens as green (not as green as callus), rounded protrusions that resemble callus but with a whitish furlike surface.
- Presence or absence of shoots: shoots were seen with a hand lens as green protrusions that resemble a fern frond about 1 mm in length.
- Presence or absence of roots: roots were seen with a hand lens as blackish hair like growth from the base of the sporophyte plant.
- Sporophyte development
- Frond length was measured while the sporophyte is still in the test tube (approximate) with the help of a hand lens and a ruler (in millimeter). The average was noted down as frond length.
- Root length was measured while the sporophyte is still in the test tube (approximate) with the help of a hand lens and a ruler (in millimeter). The average was noted down as root length.
- Numbers of fronds of the sporophyte inside the test tube were counted with the help of a hand lens.
- Numbers of roots of the sporophyte inside the test tube were counted with the help of a hand lens.
Microscopic examination of gametophyte development was done by getting samples during the development process and fixing onto a slide with alcohol sealed with transparent nail polish and photomicrographed at the same time.
Method of Analysis
The following data were statistically analyzed: data on percent contamination and viability of Nephrolepis runners using different hypochlorite concentrations; data on percent of leafy shoots of Nephrolepis runners using different segments and position in the agar medium; mean frond length, mean frond number, mean root length and mean root number for Nephrolepis sporophytes in different growth regulator and sucrose concentrations. Analysis of Variance (ANOVA) was done to test if the differences in the values obtained were significant. Duncan�s Multiple Range Test (DMRT) was done to check which of the values are significantly different or not. (Reyes 1996)
RESULTS
Effects of different concentrations of sterilizing agents
Contamination was lowest when the explants (after the alcohol dip) were agitated for 20 minutes in 50% hypochlorite but the percentage of the explants that died were the highest at same treatment. The best treatment was 20 minutes at 20% hypochlorite since the explants remained viable after 28 days with a 25% contamination rate only.
Effects of absence or presence of wetting agent
Tween 20 was a better wetting agent because it gave 0% contamination and the explants were 100% viable after 28 days. The liquid detergent only gave 25% contamination rate and 100% viability.
Effect of runner part
All Nephrolepis runners with node and runner tips remained green even until the 28th day of culture. Fifteen-millimeter runner tip was the best explant source for Nephrolepis biserrata in terms of emergence of leafy shoots.
Effect of vertical or horizontal position
Vertical position of the runner explant with the tip embedded into the agar (vertical tip downward) was the best in terms of greening, swelling of the tip, emergence of buds and leafy shoots.
Effect of different concentrations of MS salt
Sporophytes first emerged in � strength MS salt for both Nephrolepis and Pteris. This MS salt concentration was used throughout the rest of the experiments.
Effect of different concentrations of growth regulators
- Nephrolepis biserrata
Leafy shoots first emerged on the 21st day at the medium with 1 mg/l BAP and 0.5 mg/l NAA, while buds appeared on the same day in the control (no growth regulators). Callus were observed on the 28th day in the other medium with growth regulators (1 mg/l BAP + 0.1 g/l NAA, 1 mg/l BAP + 1 mg/l NAA and 2 mg/l BAP + 1 mg/l NAA). Buds from the callus first appeared on the 42nd day in the medium with the highest growth regulator concentrations of 2 mg/l BAP and 1 mg/l NAA. Fronds appeared on the 49th day on the same medium. Longest frond was seen in the same medium but more fronds can be seen when there were no growth regulators present. Root was longest and more numerous in the medium with 1 mg/l BAP and 0.1 mg/l NAA.
- Pteris vittata
Spores of Pteris first germinated (greening) (Figure 11) in the medium with 1 mg/l BAP and 0.1 mg/l NAA. Heart shaped prothallus (Figure 12) and gametophyte clumping (Figures 13, 14 and 15) were observed. Callus (Figures 13, 14 and 15) first emerged in the medium with 2 mg/l BAP and 0.1 mg/l NAA. . Microscopic examination of the gametophyte formation and development were observed (Figures 16 to 22). Sporophytes were first observed in the same medium. Sporophytes from gametophytes (Figure 23) were best grown in the medium with 1 mg/l BAP and 0.1 mg/l NAA in terms of frond length, frond number, root length. More roots were observed in the medium with 1 mg/l BAP and 1 mg/l NAA. Observations of sporophytes from callus were delayed by 2 weeks since there is still the organogenesis from callus to bud that was taken into consideration. Growth of sporophyte from the callus was best in the medium with 1 mg/l BAP and 0.1 mg/l NAA in terms of frond length, frond number, root length and number of roots (Table 11).
Effect of different concentrations of sucrose
Both ferns exhibited better growth in terms of first appearance of shoot and germination with a higher concentration of sucrose at 30 g/l. Pteris vittata callus was observed first in the same medium.
- Nephrolepis biserrata
Sporophytes were observed to have better frond elongation and more numerous fronds in the media with the higher concentration of sucrose. Root elongation and proliferation was best seen in the medium with lower sucrose concentration.
- Pteris vittata
Sporophytes from gametophytes in the media with the higher sucrose concentration gave the longest fronds. Frond number difference between the 2 concentrations is only minimal. Roots started out as black clumps (Figure 24) and then as hairs that developed into roots (Figure 25). Longer and more numerous roots were observed in the medium with lower sucrose concentration. Sporophytes from the callus also exhibited the same reaction in terms of frond length, frond number, root length and root number even if the observation was delayed for 2 weeks. Brown calli (Figure 26) were discarded.
DISCUSSION
Hypochlorite solutions are commonly used for sterilizing explants. The stronger the concentration of the sterilizing agent and the longer the time of exposure of the explant, the more the explants are damaged as was observed also by Gamborg (1995). In the case of Nephrolepis biserrata and Pteris vittata explants, a 30 second to one minute dip in alcohol and twenty minutes of a 20% hypochlorite (Zonrox bleach) solution with 1 drop of Tween 20 was sufficient to sterilize the explant without too much damage. Washing with sterile distilled water between sterilizing agents ensured that the explant was not exposed longer to the sterilizing agents. Care should be taken when sterilizing an explant especially a delicate tissue like the runner tip. Only a slight agitation from time to time is needed to prevent damage to the explant. A wetting agent like Tween 20 ensures that the hypochlorite solution sterilize the explant thoroughly by acting as a surfactant. It wets the surface of the explant thereby making a better contact of the hypochlorite solution and the explant. When Tween 20 is not available, a liquid detergent like �Joy Ultra with Anti-bac� (available in the local market) also gave good results.
Most of the Nephrolepis runner explants remained green during culture and maybe due to endogenous photosynthates (Loescher and Albert 1979). The 100% emergence of leafy shoots in the runner node is probably due to the presence of a shoot bud that resumes growth as was also observed by Loescher and Albrecht (1979) or the presence of a secondary stolon primordia that expands and initiate leaf primordia forming new leafy shoots (Richards et al. 1983). Runner tips were observed to be the best explant source with regards to emergence of leafy shoots. Beck and Caponetti (1983) noted similar observation. This maybe due to the presence of leaf primordia that was observed by Richards et al. (1983) in his longitudinal section of a runner. The use of the runner tip explant is most suitable because Nephrolepis runners usually initiate shoots and roots in vivo when it gets in contact with a substrate like soil and in vitro a substrate like agar. In vitro creeping stolons were observed (Figure 27) in this study and by Loescher and Albrecht (1979). Runner tips embedded in agar usually swells then show buds (Figure 28) and afterwards leafy shoots emerged (Figure 29).
Half strength MS salts is the best medium observed in this research and is consistent with existing publications of researches done in the past regarding different ferns (Fernandez et al. 1997; Kwa et al. 1995; Kyte and Kleyn 1996; Edwards and Roux 1998; Teng 1997; Teng and Teng 1997; Zamora et al. 1993). Nephrolepis leafy shoots first emerged with 1 mg/l BAP and 0.5 mg/l NAA, while buds appeared on the same day in the control (no growth regulators). More fronds (single sporophyte experiment) can be seen when there were no growth regulators present. This suggests no significant difference between absence and presence of growth regulators in the medium. Beck and Caponetti (1983), Loescher and Albrecht (1979) and Richards et al. (1983) had the same observation. Callus was observed in other medium with growth regulators. Hormones also induces the whole runner tip to form callus (Figure 30). Buds develop from this callus (Figure 31) and a sporophyte emerges. The callus was actually masses of multiple shoots and is a characteristic response of the explant to hormones according to Beck and Caponetti (1983). Root was longest and more numerous in the medium with 1 mg/l BAP and 0.1 mg/l NAA. Beck and Caponetti (1983) found out that lower hormone concentrations speeded rooting. Higher concentrations of sucrose at 30 g/l promoted longer and more numerous fronds and lower concentrations at 10 g/l promoted longer and more numerous roots. Rigby (1998) stated that the higher the level of sugar, the greater the shoot multiplication rate and the lower rhizoid production. Growth and multiplication seem to respond mostly to salt and sugar concentrations and is consistent with the research done by Beck and Caponetti (1983), Loescher and Albrecht (1979); Richards et al. (1983) and Rigby (1998)
Spores of Pteris vittata were easy to germinate (7 days) in the medium with 1 mg/l BAP and 0.1 mg/l NAA although germination also occurred in the absence of growth regulators and in higher sucrose concentration (30 g/l). Dicksonia antarctica (tree fern) spores that were collected and tried under the same culture conditions did not germinate after five months and might be due to the fact that the spores were no longer viable. Spores of Cyathea contaminans (tree fern) collected in Baguio were cultured and showed signs of germination after 30 days. The Cyathea spores were collected at a late stage in the school year so observations will not be included in this paper. The possible difference in spore coat morphology of these two tree ferns from Pteris vittata spores might be the key for the difference in germination time. The sexine of Cyathea contaminans is as thick as its nexine while in Pteris vittata the sexine is thinner than the nexine. Cyathea contaminans sexine is also thicker than Pteris vittata sexine (Joaquin 1986).
The microscopic examination of the development of the gametophyte stage is based on findings in Cheilantes dilimanensis (Zamora et al. 1993) and Athyrium esculentum (Nayar 1960). Callus was seen first in higher hormone concentrations but other media also exhibited callus formation. Spontaneous appearance of callus from gametophytes were observed in Pteris vittata (Kato 1963) and only sporophytes arise from this callus. Some ferns cultured in vitro also exhibited callus formation like in Pteridium aquilinum (Steeves and Sussex 1952), Pteris cretica (Bristow 1962) and Osmunda cinnamomea (Morel and Wetmore 1951). These sporophytes might be considered haploid since no fertilization occurred. Further study of these callus-derived sporophytes should be done. Sporophytes were also observed arising from the gametophyte (Figure 32) but were not studied further. This maybe due to the fusion of gametes or apogamously derived from the gametophyte itself (Whittier 1964). This can be another area for further research. Abnormal growth of the gametophyte (Figure 33) of Pteris vittata culture has been noted. Gametophyte outgrowths were also observed in Pteris longifolia (Albaum 1938), Pteridium aquilina (Partanen 1955, Whittier and Steeves 1960, Steeves et al. 1955). Frond length and frond number was best in the same medium for both callus-derived and gametophyte-derived sporophytes. Rooting was best in lower hormone concentrations or no hormones but lower sucrose at 10 g/l. No considerable difference in media for initiating sporophytes from spores was noted except for the rooting part, which is consistent with other findings. More sucrose (20-30 g/l) in the media promotes sporophyte growth (Wetmore 1953) and less sucrose (10 g/l) promotes root growth.
A giant fern (Angiopteris sp.) was tested and leaflets and cut pieces of young frond were used in culture. Angiopteris sp. is from the Family Marattiaceae (Bower 1963, Copeland 1947, Ogura 1972, Salgado 1990, Sporne 1968, Madulid 1995, Riffle 1998). The test was done since some ferns even produce sporophytes directly from leaves as seen in Grammitis tenella (White 1968). The leaf anatomy of ferns was discussed at length (Bower 1963, Ogura 1972). No growth was observed using the leaflets. The growth observed with the very young frond can be described as a callus (Figure 34). The meristem is actively dividing (Gunckel 1965) and will generally form a callus (Yeoman and Macleod 1977) when used for tissue culture. In this case it can be said that the very young frond is meristematic.
CONCLUSION AND RECOMMENDATIONS
This study showed that micropropagation of ferns using spores and vegetative parts can be done with � strength MS salts and variations in sucrose and growth regulator concentrations. Further study of the morphogenesis of Pteris vittata gametophytes (especially anatomical and histological observations) is recommended to emphasize from where the sporophyte was initiated. Callus cultures of Pteris vittata and the subsequent growth of sporophytes from this callus is another area open for further research. Nephrolepis biserrata in vitro techniques and media can be applied to other species of more commercially desirable Nephrolepis sp. or any fern that exhibit runner growth. In vitro spore culture techniques gathered from this research can be applied to more commercially expensive and hard to propagate ferns like Cyathea contaminans. The minimal results of Angiopteris sp. using fronds grown in vitro can be continued as further study. The Cyathea contaminans culture can also be continued. Stock plants or explants should be readily available to the researcher so that more time will be allotted for observations of in vitro techniques. The ornamental plant industry which this study aims to help can start with minimal media such as � strength MS salts with various sucrose concentrations to propagate desired ferns from spores or runners.
LITERATURE CITED
Albaum, H.G. 1938a. Inhibitions due to growth hormones in fern prothallia and sporophytes. Am. J. Bot. 25:124-132.
____________. 1938b. Normal growth, regeneration and adventitious outgrowth formation in fern prothallia. Am. J. Bot. 25(1):37-44.
Basile, D.V. 1973. A simple method of initiating axenic cultures of Pteridophytes from spores. Am. Fern J. 63(4):147-151.
Bristow, J.M. 1962. The controlled in vitro differentiation of callus derived from a fern, Pteris cretica L., into gametophytic or sporophytic tissues. Devt. Bio 4:361-375.
Beck, M.J. and Caponetti, J.D. 1983. The effects of kinetin and naphthaleneacetic acid on in vitro shoot multiplication and rooting in the fishtail fern. Am.J.Bot. 70(1):1-7.
Bower, F.O. 1963. The ferns (Filicales). V.2 The Eusporangiatae and other relatively primitive ferns. India: Jayyed Press. 344 p.
Bridgers, B.T. 1958. Techniques involving the sowing of spores of ferns for artificial culture. Am. Fern J. 48(4):161-164.
Brown, C.A. 1960. What is the role of spores in fern taxonomy. Am. Fern J. 50(1):6-14.
Copeland, E.B. 1947. Genera Filicum. Mass., USA: Chronica Botanica Company. 247 pp.
DeMaggio, A.E. and Wetmore, R.H.1961. Morphogenetic studies on the fern Todea barbara. III. Experimental embryology. Am. J. Bot. 48:551-564.
Dougall, D.K. 1981. Media factors affecting growth. En. Exp. Bot. 21:277-280.
Edwards, E.S. and Roux, S.J. 1998. Gravity and light control of the developmental polarity of regenerating protoplasts isolated from prothallial cells of the fern Ceraptoris richardii. Plant Cell Rep. 17:711-716.
Fernandez, H., Bertrand, A.M. and Tames, R.C. 1997. Gemmation in cultured gametophytes of Osmunda regalis. Plant Cell Rep. 16:358-362.
Fliflet, T. 1961. Growing ferns from spores. Am. Fern J. 51(3):113-127.
Foster, F.G. 1956. The microscopy of fern spores. Am. Fern J. 46:7-14.
Gamborg, O.L. and Phillips, G.C. 1995a. Appendix A: Basal media for plant cell and tissue culture. Plant cell, tissue and organ culture: Fundamental methods. Springer lab manual. 301-307p.
____________________________. 1995b. Media preparation and handling. Plant cell, tissue and organ culture: Fundamental methods. Springer lab manual. 21-34p.
____________________________. 1995c. Sterile Techniques. Plant cell, tissue and organ culture: Fundamental methods. Springer lab manual. 37-42p.
Griswold, M.M. 1956. Models for spore study. Am. Fern J. 46(1):1-6.
Gunckel, J.E. 1965. Factors affecting the morphogenesis of plant organs. Proc. Intl Conf. Plant Tissue Culture. 251-268.
Hvoslef-Eide, A.K. 1992(a). Effects of pre-storage conditions on storage of in vitro cultures of Nephrolepis exaltata (L.) Schott and Cordyline fruticosa (L.) A. Chev. Plant Cell and Org Cult 28:167-174.
________________. 1992(b). Influence of nitrogen fertilization to mother plants and the subsequent growth of Nephrolepis exaltata (L.) Schott and Cordyline fruticosa (L.) A. Chev. in vitro explants. Gartenbauwissenschaft 57:292-297.
_________________. 1991. Mother plant temperature effects on growth of in vitro propagated daughter plants of Nephrolepis exaltata (L.) Schott. Sci Hort 47:149-156.
_________________. 1990. The effect of irradiance and temperature on in vitro cultures of Nephrolepis exaltata (L.) Schott and Cordyline fruticosa (L.) A. Chev. Gartenbauwissenschaft 55:259-264.
_________________. 1991. The effect of temperature, daylength and irradiance on the growth of mother plants of Nephrolepis exaltata (L.) Schott and on the subsequent growth in vitro of runner tip explants. Sci. Hort. 47:137-147.
Hirsch, A.M. 1975. The effect of sucrose on the differentiation of excised fern leaf tissue into either gametophytes or sporophytes. Plant Physiol. 56:390-393.
Hu, C. and Zanettini, M.H.B. 1995. Embryo culture and embryo rescue for wide cross hybrids. Plant cell, tissue and organ culture: Fundamental methods. Springer lab manual. 129-141p.
Joaquin, C.C. 1986. Spore morphology of Philippine ferns. PhD. Thesis. UP Diliman, QC. 346p.
Kato, Y. 1963. Physiological and morphogenetic studies of fern gametophytes in aseptic culture. I. Callus tissues from dark-cultured Pteris vittata. Bot. Gaz. 124:413-416.
Kleinschmidt, W.F. 1952. Growing ferns from spores. Am. Fern J. 42(4):117-124.
_______________. 1957. A method of preparing spores for fern cultures. Am. Fern J. 47(3):95-98.
Knobloch, I.W. 1957. Gibberellic acid and ferns. Am. Fern J. 47:134-135.
____________. 1960. Some hints for the fern culturist. Am. Fern J. 54(2):73-76.
Knudson, L. 1925. Physiological study of the symbiotic germination of orchid seeds. Bot. Gaz. 79(4):345-379.
__________. 1940. Permanent changes of chloroplasts induced by x rays in the gametophyte of Polypodium aureum. Bot. Gaz. 101(4):721-758.
Kwa, S.H., Wee, Y.C., Lim, T.M. and Kumar, P.P. 1995. IAA-induced apogamy in Platycerium coronarium (Koenig) Desv. gametophytes cultured in vitro. Plant Cell Rep. 14:598-602.
Kyte, L. and Kleyn, J. 1996. Plants from test tubes: An introduction to micropropagation. Portland, OR: Timber Press. 240 pp.
Loescher, W.H. and Albrecht, C.N. 1979. Development in vitro of Nephrolepis exaltata cv. Bostoniensis runner tissues. Physiol. Plant. 47:250-254.
MacFadden, F. 1963. Remarks on the care of ferns I cultivation. Am. Fern J. 53(4):55-59.
Madulid, D. A. 1995. A pictorial cyclopedia of Philippine ornamental plants. Manila: Bookmark. 388 p.
Marengo, N.P. 1979. A simplified medium for growing fern prothallia. Am. Fern J. 69(4):122.
Mikkelsen, E.P. and Hasegawa, P.M. 1981. Report of the round-table discussion on ornamentals. Env. Exp. Bot. 21(3/4):411-413.
Miller, J.H. 1968. Fern gametophytes as experimental material. Bot. Rev. 34(4):361-426.
Morel, G. and Wetmore, R.H. 1951. Fern callus tissue culture. Am. J. Bot. 38:141-143.
Morton, C.V. 1958. Observations on cultivated ferns. V. The species and Forms of Nephrolepis. Am. Fern J. 48:18-27.
Murashige, T. 1974. Plant propagation through tissue cultures. Ann. Rev. Plant Physiol. 25:135-166.
Murashige, T. and Skoog, F. 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 15:473-497.
Nayar, B.K. 1960. The gametophyte and young sporophyte of Athyrium esculentum. Am. Fern J. 50(2):194-203.
Ogura, Y. 1972. Comparative anatomy of vegetative organs of the pteridophytes. Germany: Gebruder Borntraeger. 502 pp.
Oki, L.R. 1981. The modification of research procedures for commercial propagation of Boston ferns. Env. And Ex. Bot. 21:397-400.
Partanen, C.R., Sussex, I.M. and Steeves, T.A. 1955. Nuclear behavior in relation to abnormal growth in fern prothalli. Am. J. Bot. 42:245-256.
Phillips, G.C., Hubstenberger, J.F. and Hansen, E.E. 1995a. Adventitious shoot proliferation. Plant cell, tissue and organ culture: Fundamental methods. Springer lab manual. 56-65pp.
_______________________________________________. 1995b. Plant regeneration by organogenesis from callus and cell suspension cultures. Plant cell, tissue and organ culture: Fundamental methods. Springer lab manual. 73-79pp.
Reyes, F.C. 1996. Applied basic statistics. Quezon City: Phoenix Publ. House.408p.
Richards, J.H., Beck, J.Z. and Hirsch, A.M.. 1983. Structural investigations of asexual reproduction in Nephrolepis exaltata and Platycerium bifurcatum. Am. J. Bot. 70(7):993-1001.
Rigby, M. 1998. "Fern tissue culture: Microsorum pteropus ferns" http://www.egroups.com/[email protected]/ (July 19, 1999)
Riffle, R. L. 1998. The tropical look: An encyclopedia of landscape plants for worldwide use. London: Timber Press. 428 p.
Salgado, A. E. 1990. A checklist of Philippine ferns. Phil. J. Sci. 119(2):107-148.
Sheffield, E., Douglas, G.E. and Cove, D.J. 1997. Growth and development of fern gametophytes in an airlift fermenter. Plant Cell Rep. 16:561-564.
Sheffield, E. and Bell, P.R. 1987. Current studies in Pteridophyte life cycle. Bot. Rev. 53(4):442-490.
Slack, S.A. and Tufford, L.A. 1995. Meristem culture for virus elimination. Plant cell, tissue and organ culture: Fundamental methods. Springer lab manual. 73-79pp.
Sporne, K.R. 1968. The morphology of pteridophytes: The structure of ferns and allied plants. London: Hutchinson and Co. Ltd. 192pp.Steeves, T.A. 1952. In vitro culture of a fern callus. Nature 170:672-673.
Steeves, T.A. and Sussex, I.M. 1957. Studies on the development of excised leaves in sterile culture. Am. J. Bot. 44:665-673.
Steeves, T.A., Sussex, I.M. and Partanen, C.R. 1955. In vitro studies on abnormal growth of prothalli of the bracken fern. Am. J. Bot. 42(3):232-244.
Steiner, M. L. 1986. Philippine ornamental plants and their care. 3rd ed. Manila:Enrian Press. 233p.
Street, H.E. 1977. Introduction. Plant tissue and cell culture. 2nd ed. Berkeley:University of California Press. 1-11pp.
Sussex, I.M. and Steeves, T.A. 1953. Growth of excised fern leaves in sterile culture. Nature 172:624-625.
Swami, P. and Raghavan, V. 1980. Control of morphogenesis in the gametophyte of a fern by light and growth hormones. Can. J. Bot. 58:1464-1473.
Szweykowska, A. 1974. The role of cytokinins in the control of cell growth and differentiation. in culture. Tissue Culture and Plant Science. Proc. of 3rd Intl. Cong. of Plant Tissue and Cell Culture: Leicester
Teng. W.L. 1997. Activated charcoal affects morphogenesis and enhances sporophyte regeneration during leaf cell suspension culture of Platycerium bifurcatum. Plant Cell Rep. 17:77-83.
Teng, W.L. and Teng, M.C.. 1997. In vitro regeneration patterns of Platycerium bifurcatum leaf cell suspension culture. Plant Cell Rep. 16:820-824.
Villareal, R.L., Rasco, E.T., Mendoza, E.T., Barba, R.C. and Zamora, A.B. 1989. Plant biotechnology program of the Institute of Plant Breeding. Asean Science and Technology Week. Conference proceedings. 2:186-201.
Warne, T.R., Walker, G.L. and Hickok, L.G. 1986. A novel method for surface-sterilizing and sowing spores. Am. Fern J. 76(4):187-188.
Wetmore, R.H. 1953. Carbohydrate supply and leaf development in sporeling ferns. Science 118(3072):578.
White, R.A. 1968. Vegetative reproduction in the ferns I. Leaf buds of Grammitis tenella. Am. Fern J. 58(3):108-118.
Whittier, D.P. 1964. The influence of cultural conditions on the induction of apogamy in Pteridium gametophytes. Am. J. Bot. 51(7):730-736.
___________. 1981. Spore germination and young gametophyte development of Botrychium and Ophioglossum in axenic culture. Am. Fern J. 71(1):13-19.
Whittier, D.P. and Steeves, T.A. 1960. The induction of apogamy in the bracken fern. Can. J. Bot. 38: 925-1531.
Yeoman, M.M. and Macleod, A.J. 1977. Tissue (callus) cultures-Techniques. Plant tissue and cell culture. 2nd ed. Berkeley:University of California Press. 31-59 pp.
Zamora, P.M. 2000. Urban ferns and fern allies. Quezon City: University of the Philippines Press. 170pp.
Zamora, P.M., Chaimongkol, S., Beltran, C. and Marzan, M. 1993. Structure and development of the gametophytes of Philippine Cheilanthoid ferns, I. Cheilanthes dilimanensis Zamora. Phil. J. Sci. 122:233-253.
Zamora, P.M. and Co, L. 1986. Guide to Philippine flora and fauna: Economic ferns, Endemic ferns. Quezon City: JMC Press Inc 1-210pp.
FIGURES
| Click thumbnail for larger picture |

|

|

|

|

|

|

|

|
|
| Figure 1 |
Figure 2 |
Figure 3 |
Figure 4 |
Figure 5 |
Figure 6 |
Figure 7 |
Figure 8 |
Figure 9 |

|

|

|

|

|

|
|

|

|
| Figure 10 |
Figure 11 |
Figure 12 |
Figure 13 |
Figure 14 |
Figure 15 |
Figure 16 |
Figure 17 |
Figure 18 |

|

|
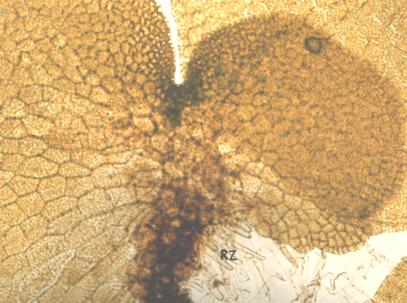
|
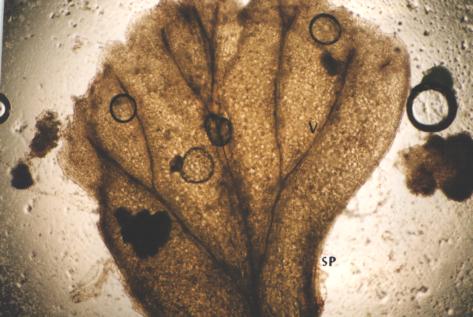
|

|

|

|

|

|
| Figure 19 |
Figure 20 |
Figure 21 |
Figure 22 |
Figure 23 |
Figure 24 |
Figure 25 |
Figure 26 |
Figure 27 |
|

|
|

|

|

|

|
 |
|
|
Figure 28 |
Figure 29 |
Figure 30 |
Figure 31 |
Figure 32 |
Figure 33 |
Figure 34 |
|