{kind=link}
{kind=link}
{kind=link}
| Brasil,
|
 |
INTRODUÇÃO
A aplicação da biologia molecular em microbiologia é
muito vasta, passando pela área da sistemática microbiana com
base molecular, pelo estudo da evolução dos microrganismos e
também pela sua aplicação na biotecnologia através
da engenharia genética, que permite ao homem a produção
de várias substâncias úteis, em que a insulina e antibióticos
são bons exemplos.
Os microrganismos apresentam uma grande diversidade e, muitos deles são ainda desconhecidos do homem. Mas como surgiu toda essa diversidade? Atualmente é claro que uma tão grande variedade das características morfológicas, fisiológicas e ecológicas dos diversos grupos de microrganismos é controlada pela sua constituição genética dos organismos em interação com o ambiente do qual fazem parte. Esta diversidade microbiana surge como o resultado da evolução: mutações e recombinação genética que ocorrem nos microrganismos que são capazes de viver em vários habitats. Quando um novo habitat surge, fica sujeito à colonização quer pelos organismos pré-existentes quer por novos microrganismos que tenham surgido por mutação genética. Os microrganismos mais aptos para crescer e proliferar e, ainda, competir com os outros de forma vantajosa estabelecem-se nesse novo habitat. Assim, o ambiente está constantemente a selecionar os indivíduos mais adaptados. A grande variedade físico-química dos habitats existentes na Terra permite a grande diversidade de microrganismos que atualmente se conhecem.
Com o conhecimento atual é possível escolher a composição genética das células com o objetivo de produzir novas substâncias. Este processo é chamado de engenharia genética ou tecnologia recombinante de DNA. Os investigadores podem combinar desta forma características únicas de células tão diferentes como uma célula bacteriana e uma célula humana. Assim, com estas técnicas, microrganismos foram transformados para a produção industrial de várias substâncias como o hormônio do crescimento, o agente anticancerígeno interferon, vacinas, para além dos já referidos.
Os avanços científicos
que tornaram possível a engenharia genética têm implicações
excitantes e amplas para o futuro de cientistas e das pessoas em geral. Pela
introdução de genes estranhos em microrganismos é possível
desenvolver estirpes de micróbios que apresentam novas soluções
para problemas tão diversos como a poluição, a falta
de alimento ou de energia, e ainda para o controlo de doenças. Pelo
fato dos cientistas poderem agora possuir microrganismos novos, únicos
e extremamente úteis, uma nova indústria se desenvolve com base
na engenharia genética - a biotecnologia.
SISTEMÁTICA
MICROBIANA
A imensa biodiversidade existente no nosso planeta levou a que, desde cedo,
o Homem procurasse um modo de organizar, ordenar e denominar os seres vivos.
Os primeiros sistemas de classificação geravam grupos cujos
organismos, normalmente, partilhavam características morfológicas,
independentemente das suas relações filogenéticas. Ao
longo do tempo, com o aprofundamento das ciências biológicas,
novos sistemas de classificação têm sido propostos. A
taxonomia, embora indispensável a outras ciências, uma vez que
permite a existência de uma linguagem científica universal, depende
de conceitos e metodologias de várias disciplinas que possibilitam
a interpretação e integração dos dados taxonômicos.
Durante a primeira metade do século XX, a classificação filogenética dos organismos multicelulares teve um grande desenvolvimento. Contrariamente, a simplicidade morfológica dos microrganismos procarióticos, por não permitir a ocorrência de um registro fóssil fiável para estudos de filogenia, desencorajou a criação de sistemas de classificação baseados nas relações de ancestralidade dos organismos. Os primeiros sistemas de identificação e comparação dos microrganismos baseavam-se num pequeno número de características, principalmente morfológicas e fisiológicas, escolhidas subjetivamente pelo taxonomista. Algumas destas características eram consideradas de maior relevância, sendo, por isso, suficientes para o estabelecimento dos agrupamentos. Deste modo, tais sistemas de classificação resultavam, normalmente, no agrupamento de organismos que não possuíam verdadeiras afinidades.
A partir da década de 50, novos conceitos de classificação foram introduzidos na sistemática microbiana. O conceito de classificação natural, segundo Gilmour, descreve a formação de grupos com elementos que partilham o máximo de características fenotípicas, todas com a mesma relevância. Os sistemas de classificação em que o agrupamento dos organismos é baseado na similaridade dos seus indivíduos, incluindo características fenotípicas e genotípicas, sem referência a conceitos de ancestralidade denominam-se fenéticos.
O desenvolvimento de técnicas de seqüenciamento de proteínas e de ácidos nucléicos e a sua aplicação à sistemática microbiana vieram permitir o esclarecimento das relações filogenéticas dos organismos procarióticos. Estes estudos, iniciados por Woese (1987), baseiam-se, principalmente, na comparação de zonas conservativas do gene de rRNA 16S. Recentemente, várias macromoléculas, ou os genes que as codificam, têm sido utilizados para esclarecer as relações de ancestralidade dos organismos. Contrariamente aos sistemas de classificação fenética, na classificação filogenética os organismos são agrupados de acordo com a sua antiguidade. Deste modo, em filogenia não é considerada a similaridade atual entre os indivíduos.
Alguns componentes celulares, embora com distribuição universal, apresentam, em diferentes grupos de organismos, variabilidade suficiente para que possam ser aplicados como marcadores taxonômicos. Nesta área da taxonomia, designada por quimiotaxonomia, faz-se a análise comparativa de componentes celulares como os ácidos nucléicos, os componentes da parede celular, os lipídeos, as proteínas, os ácidos gordos e as quinonas respiratórias.
As classificações mais estáveis e úteis são aquelas em que os taxa são comparados através da aplicação de vários métodos de caracterização taxonômica independentes, baseadas num conjunto de dados tão lato quanto possível, integrando informação fenotípica e genotípica. As características fenotípicas devem incluir, para além das propriedades morfológicas e fisiológicas, características quimiotaxonômicas. Um correto sistema de classificação microbiano deve ser uma abordagem polifásica, isto é, basear-se em informação fenética e filogenética.
O objetivo último
da sistemática microbiana atual é delinear categorias taxonômicas
filogeneticamente coerentes e fáceis de reconhecer através de
características fenotípicas.
Conceito de espécie
A unidade taxonômica
básica da sistemática microbiana é a espécie.
Tradicionalmente uma espécie bacteriana era distinguida por correlação
de características fenotípicas e os membros de uma espécie
possuíam um conjunto de características únicas. Esta
definição de espécie é subjetiva, pois depende
dos critérios adotados pelo taxonomista. Embora não haja uma
definição universal de espécie em bacteriologia podem
distinguir-se grupos denominados por:
- taxospécies, em que as estirpes desse grupo partilham uma elevada
proporção de características fenéticas;
- genospécies, grupos de organismos capazes de trocar informação
genética;
- nomenspécies, grupos de organismos que possuem características
comuns, independentemente da validade desses grupos com base noutros estudos.
- espécies genômicas, grupos de organismos com elevada similaridade
genômica traduzida por altos valores de reassociação do
DNA.
A espécie genômica é a única que pode ser definida
em termos filogenéticos.
Para a descrição
de uma espécie bacteriana é necessário o isolamento de
várias estirpes bacterianas e recomenda-se que uma espécie seja
constituída por estirpes em que os valores de homologia de DNA sejam
superiores a 70% e que o valor de Tm [diferença entre o ponto de fusão
do DNA heterólogo (de estirpes diferentes) e homólogo (da mesma
estirpe)] seja inferior a 5ºC. Estes resultados devem sempre ser apoiados
por características fenotípicas. Os resultados de homologia
de DNA devem ser confirmados com estudos de seqüenciamento de rRNA 16S.
TÉCNICAS
UTILIZADAS
Nas últimas décadas, têm sido desenvolvidas inúmeras
técnicas de classificação e identificação
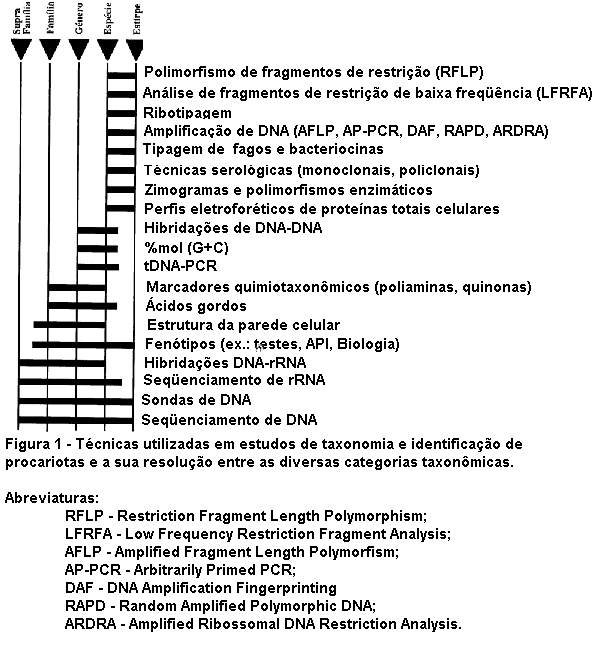
de microrganismos (Figura 1). Embora todas
as técnicas de classificação se baseiem, direta ou indiretamente,
na informação genética contida na célula, independentemente
do componente celular em análise, há vários níveis
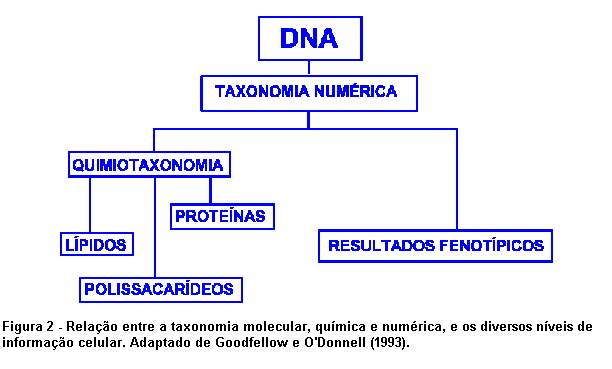
de informação (Figura 2).
A seqüência completa de bases nucleotídicas do genoma bacteriano é o primeiro nível de informação celular e, portanto, é considerado como referência padrão para a estimativa dos relacionamentos entre os microrganismos. Na prática, é impossível determinar a seqüência completa das bases nucleotídicas, mas um grande número de técnicas de análise genômica tem sido desenvolvido para a classificação dos microrganismos, como, por exemplo, a seqüenciamento do gene de rRNA 16S ou de zonas conservativas de DNA, a estimativa da composição em bases do DNA, a hibridação de DNA-DNA ou de DNA-rRNA, o polimorfismo de fragmentos gerados por enzimas de restrição e a análise de ácidos nucléicos através de sondas de oligonucleotídeos.
O segundo nível de informação é representado pelas proteínas que são uma cópia direta do genoma. O terceiro nível é expresso na estrutura de um grande número de componentes e produtos celulares, que são marcadores quimiotaxonômicos como, por exemplo, aminoácidos, açúcares e lipídeos. Finalmente, o quarto nível de informação traduz-se na morfologia e no comportamento das células.
O tipo de informação
que se analisa e os organismos em estudo condicionam, de certo modo, o nível
de categoria taxonômica a que é possível a sua discriminação.
Na figura 1 descrevem-se algumas técnicas
atualmente utilizadas e a sua resolução entre as diversas categorias
taxonômicas.
Composição em bases nucleotídicas
A composição em bases nucleotídicas, expressa pela percentagem molar do conteúdo em guanina e citosina [%mol (G+C)], é utilizada para caracterizar o genoma microbiano. Normalmente, esta é também uma das características requeridas para descrever espécies ou gêneros de microrganismos procarióticos.
A desnaturação térmica do DNA é um dos métodos mais utilizados para determinar a sua composição em bases. A separação das cadeias é acompanhada pelo aumento da absorvência a 260 nm. O ponto de fusão do DNA, Tm, corresponde ao ponto médio de absorbância entre o estado não desnaturado e de desnaturação total da dupla cadeia de DNA. O ponto de fusão do DNA é proporcional ao conteúdo em guanina e citosina, uma vez que estas bases nucleotídicas se ligam através de três pontes de hidrogênio enquanto que a timina e a adenina se ligam apenas por duas. Assim, quanto maior for o conteúdo em (G+C), maior a quantidade de energia térmica requerida para separar as duas cadeias de DNA.
O conteúdo em
%mol de (G+C) pode variar entre 24 e 76%, dependendo do grupo de microrganismos.
Normalmente, o conteúdo em %mol de (G+C) de estirpes que constituem
uma espécie filogeneticamente coerente não difere mais de 3%,
podendo apresentar maior variação entre diferentes espécies
de um gênero.
Homologia de DNA
A capacidade do DNA de cadeia simples em reassociar, sob condições apropriadas, para formar DNA de cadeia dupla é conhecida desde o início da década de 60. Com base nesta propriedade é possível avaliar a similaridade dos genomas entre diferentes organismos, havendo diversos métodos disponíveis.
Um destes métodos baseia-se na determinação espectrofotométrica do valor das taxas iniciais de reassociação. Este método baseia-se no princípio de que a taxa de reassociação de amostras de DNA de cadeia única varia de acordo com a complementaridade das cadeias reassociantes. Deste modo, a taxa máxima de reassociação ocorre entre cadeias únicas de DNAs homólogos. Note-se que dois organismos só podem ser comparados se apresentarem pelo menos 80% de similaridade de seqüências primárias dos seus DNAs. Por outro lado, os valores de reassociação de DNA não medem a similaridade da estrutura primária do DNA, mas sim a sua complementaridade. Realmente, híbridos que apresentem 90 a 95% de similaridade nas seqüências das suas bases nucleotídicas podem apresentar valores de hibridação DNA - DNA de 50% .
Os estudos de homologia
do DNA apresentam a vantagem de permitirem a comparação entre
quaisquer tipos de microrganismos procarióticos, independentemente
do seu metabolismo ou condições de crescimento.
Na maior parte dos estudos que utilizam este método, os organismos
que apresentam valores de homologia elevados (acima de 50%) têm características
fenotípicas semelhantes. A correlação entre propriedades
fenotípicas e genotípicas, levou à proposta de que os
organismos procarióticos que possuam valores de homologia de DNA iguais
ou superiores a 70% sejam considerados como pertencentes à mesma espécie
genômica. Mais tarde estendeu-se esta recomendação para
que o valor de Tm das cadeias heterólogas não excedesse os 5ºC.
Seqüenciamento do gene de rRNA 16S
A seqüência do genoma bacteriano é o termo de comparação por excelência, mas na impossibilidade da sua completa determinação, tem sido válida a análise sistemática da seqüência da subunidade rRNA 16S ou da seqüência do gene que codifica esta estrutura no DNA genômico.
O seqüenciamento do gene que codifica o RNAr 16S permite proceder à elaboração de árvores filogenéticas onde se podem colocar vários grupos evolutivos de bactérias próximos e observar as suas relações de ancestralidade.
Quando a seqüência
do gene de rRNA 16S de várias estirpes apresenta valores superiores
a 95% são considerados como membros da mesma espécie.
BIOTECNOLOGIA
Técnicas e conceitos de genética molecular como a recombinação e transformação genética, a conjugação e mobilização de cromossomos, plasmídios, transposição e seqüências de inserção de genes, tornaram possível o desenvolvimento de metodologias sofisticadas para o isolamento, manipulação e expressão do material genético. A engenharia genética tem aplicação quer análises de rotina quer em pesquisa aplicada. Em análises de rotina, pode-se estudar o mecanismo de replicação e expressão de genes em procariontes, eucariontes e vírus. Na área da pesquisa aplicada, a engenharia genética permite o desenvolvimento de culturas microbianas transformadas capazes de originar importantes produtos, como são exemplo a insulina e o hormônio do crescimento humanas, vacinas e algumas enzimas industriais.
A engenharia genética tem aplicação comercial, e é por isso chamada de biotecnologia, revelando capacidades ilimitadas (Figura 3).
Essencial para a engenharia genética é o isolamento e purificação dos genes específicos para a substância pretendida, por um processo que se designa por clonagem de genes.
Possuindo grandes quantidades de DNA puro é possível a manipulação dos genes e dos seus produtos. Depois dos genes serem clonados, pode-se determinar a seqüência de nucleotídeos de um gene, através do qual se pode descobrir, via código genético, a seqüência de aminoácidos, da proteína. O próprio gene, depois de clonado, pode ser utilizado como sonda para determinar a estrutura de moléculas de DNA mais complexo como o do genoma humano.
Pela remoção
de genes a organismos que são de difícil manipulação
ou perigosos por possível patogenicidade e transpondo-os para microrganismos
seguros e bem caracterizados, pode-se produzir valiosas substâncias
biológicas sem grandes custos e em quantidades impensáveis até
ao advento da engenharia genética. Esta engenharia pode, literalmente,
desenhar novas seqüências/genes pela alteração da
seqüência de um gene clonado num determinado local, possibilitando
produtos biológicos muito úteis que estão indisponíveis
na sua fonte natural.
Sistemas de clonagem de genes
A clonagem é a base da maioria das técnicas de engenharia genética. A finalidade da clonagem é o isolamento de grandes quantidades de um gene específico na sua forma pura. Teoricamente é possível isolar fragmentos de DNA puro, com um único gene a partir da digestão do cromossomo com enzimas de restrição, mas, uma breve reflexão mostra a impraticabilidade desta abordagem. Vejamos, considere-se um organismo geneticamente simples como a E. coli, um gene específico desta bactéria representa 1-2 kilobases do genoma de 4 700 Kb. Em média, um gene em E. coli corresponde a menos de 0,05% do DNA total da célula. No homem, o problema é ainda mais complicado, uma vez que um gene apresenta sensivelmente o mesmo tamanho, mas o genoma é 1 000 vezes maior!
Assim, a estratégia
base da clonagem de genes é a remoção do gene que se
pretende de um genoma grande e complexo para um genoma mais pequeno e simples.
Felizmente, os conhecimentos da química e da enzimologia do DNA permitem-nos
cortar e ligar a molécula de DNA in vitro. Este processo é conhecido
por recombinação in vitro. Enzimas de restrição,
DNA ligase e DNA sintetase são ferramentas importantes para a recombinação
in vitro.
A clonagem de genes pode ser dividida em vários passos:
1. Isolamento e fragmentação da fonte de DNA.
A fonte de DNA pode
ser todo o DNA de um organismo, ou DNA sintetizado a partir de um molde de
RNA pela enzima transcriptase reversa, ou mesmo DNA sintetizado in vitro a
partir de nucleotídeos.
Se o DNA genômico for a fonte de DNA, ele é, normalmente, cortado
com uma enzima de restrição para originar uma mistura de fragmentos
de menor tamanho.
2. Junção dos fragmentos de DNA a um vetor
de clonagem pela ação da DNA ligase.
Os pequenos elementos
genéticos, utilizados para replicar os genes são conhecidos
como vetores de clonagem, são replicados independentemente. Estes vetores
de clonagem são construídos, normalmente, de forma a permitir
a recombinação do DNA estranho com o DNA da célula, nos
locais de restrição cortados pelo vetor, mas não afetam
a sua replicação. Se a fonte de DNA e do vetor for cortada com
a mesma enzima de restrição, então a sua junção
pode ocorrer pelo emparelhamento das regiões de cadeia única
do DNA chamadas zonas adesivas.
3. Incorporação num hospedeiro.
A molécula
de DNA recombinante elaborada in vitro pode ser introduzido num hospedeiro
por transformação da célula ou por infecção
da célula com partículas virais, também estas elaboradas
in vitro. A incorporação do DNA no hospedeiro é feita,
normalmente, numa ampla mistura de clones bacterianos. Algumas células
contêm o gene pretendido enquanto que outras células contêm
outros clones originados pela junção do vetor de clonagem à
fonte de DNA. Esta mistura de clones é conhecida como "livraria
de DNA" ou livraria de genes, uma vez que vários clones se encontram
presentes podem ser purificados a partir daqui, possuindo cada um diferentes
fragmentos do DNA clonado.
4. Detecção e purificação
do clone requerido
Um passo crucial na
tecnologia da recombinação do DNA é o encontrar o clone
pretendido na mistura de clones originados pela recombinação
do DNA.
Procedimentos especiais são necessários para a detecção
do gene estranho no clone hospedeiro. Se o gene se expressa, a presença
de uma proteína estranha na célula pode ser detectada quer pela
sua atividade quer através de anticorpos específicos, evidenciando
assim a presença deste gene. No entanto, se o gene não é
expresso, a sua presença pode ser detectada utilizando sondas radioativas
de ácidos nucléicos, que pesquisam uma cadeia de DNA complementar.
5. Produção de um grande número
de células ou bacteriófagos
Estas células
contêm o clone pretendido para o isolamento e estudo do DNA clonado
e conseqüentemente para a produção em grande quantidade
da substância que este gene codifica.
Em resumo,
- o isolamento de grandes quantidades de um gene específico por clonagem
de genes é feito, normalmente, em bactérias utilizando um plasmídio
ou um fago como vetor de clonagem.
- enzimas de restrição e DNA ligase são as ferramentas
utilizadas na recombinação in vitro para produção
da molécula de DNA híbrida.
- uma vez introduzida, a molécula de DNA híbrida, por transformação
numa bactéria hospedeira conveniente, o DNA pode então ser produzido
em grandes quantidades por controlo do vetor de clonagem.
BIBLIOGRAFIA
Brock, T.D.; M. T. Mandigan; J. M. Martinko; J. Parker. 1996. Biology of Microorganisms, 6th edition. Prentice Hall, USA.
Pelczar, M. J., Jr; E. C. S. Chan; N. R. Krieg. 1993. Microbiology - Concepts and applications. McGraw-Hill, Inc.
Nunes, O. C. 1995. Caracterização e classificação de estirpes do gênero Rhodothermus. Tese de doutoramento. Universidade de Coimbra.
Goodfellow, M.; A. G. O'Donnell. 1993. Roots of bacterial systematics, p.3-54. In: Goodfellow, and A. G. O'Donnell (eds), Handbook of new bacterial systematics. Academic Press, USA.
Woese, C., 1987. Bacterial evolution. Microbiol. Rev. 51, 221-271.
Woese, C. R. 1994. There must be a prokaryote somewhere: microbiology's search for itself. Microbiol. Rev. 58, 1-9.
Norris, J. R. 1980. Introduction, p. 1-10. In: M. Goodfellow, and R. G. Board (eds), Microbiological classification and identification. Academic Press, USA.
Schleifer, K. H.; Ludwing, W.1994. Molecular taxonomy: classification and identification, p. 1-16. In: F. G. Priest, A. Ramos-Cormenza, B. J. Tindall (eds), Bacterial diversity and systematics. Plenum Press. USA.
Murray, R. G. E.; D.
J. Brenner, R. R. Colwell; P de Vos; M. Goodfellow; P. A. D. Grimont; N. Pfenning;
E. Stackebrandt; G. A. Zavarzin. 1990. Report of the ad hoc committee on approaches
to taxonomy within the Proteobacteria. Int. J. Syst. Bacteriol. 40, 213-215.
|
|||
|
|||