
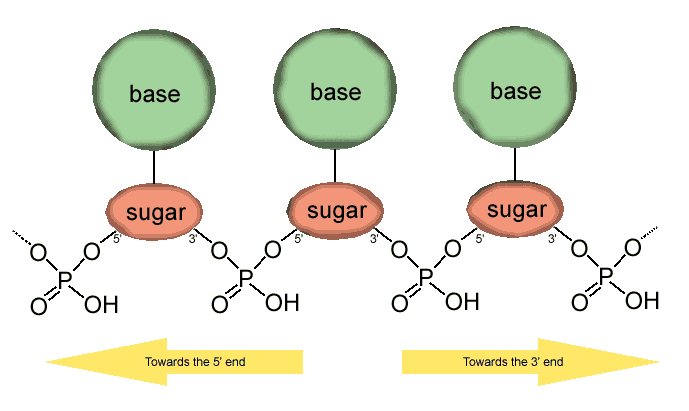
Conserved in most eukaryotic organisms, the RNAi pathway is thought to have evolved as a form of innate immunity against viruses and also plays a major role in regulating development and genome maintenance.The RNAi pathway is initiated by the enzyme dicer, which cleaves doublestranded RNA dsRNA to short doublestranded fragments of 20�25 base pairs. One of the two strands of each fragment, known as the guide strand, is then incorporated into the RNAinduced silencing complex RISC and basepairs with complementary sequences. The most wellstudied outcome of this recognition event is a form of posttranscriptional gene silencing. This occurs when the guide strand base pairs with a messenger RNA mRNA molecule and induces degradation of the mRNA by argonaute, the catalytic component of the RISC complex. The short RNA fragments are known as small interfering RNA siRNA which are perfectly complementary to the gene to which they are suppressing as they are derived from long dsRNA of that same gene or MicroRNA miRNA which are derived from the intragenic regions or an intron and are thus only partially complementary. The RNAi pathway has been particularly wellstudied in certain model organisms such as the nematode worm Caenorhabditis elegans, the fruit fly Drosophila melanogaster, and the flowering plant Arabidopsis thaliana.

;
The dicer protein from Giardia intestinalis, which catalyzes the cleavage of dsRNA to siRNAs. The RNase domains are colored green, the PAZ domain yellow, the platform domain red, and the connector helix blue. The distance between the RNase and PAZ domains, determined by the length and angle of the connector helix, may determine the length of siRNA molecules produced by dicer variants. The dicer protein from Giardia intestinalis, which catalyzes the cleavage of dsRNA to siRNAs. The RNase domains are colored green, the PAZ domain yellow, the platform domain red, and the connector helix blue. The distance between the RNase and PAZ domains, determined by the length and angle of the connector helix, may determine the length of siRNA molecules produced by dicer variants.RNAi is an RNAdependent gene silencing process that is mediated by the RNAinduced silencing complex RISC and is initiated by short doublestranded RNA molecules in the cytoplasm, where they interact with the catalytic RISC component argonaute. When the dsRNA is exogenous, coming from infection by a virus with an RNA genome or laboratory manipulations, the RNA is imported directly into the cytoplasm and cleaved to short fragments by the enzyme dicer. The initiating dsRNA can also be endogenous, as in premicroRNAs expressed from RNAcoding genes in the genome. The primary transcripts from such genes are first processed to the characteristic stemloop structure of premiRNA in the nucleus, then exported to the cytoplasm to be cleaved by dicer. Thus the two pathways for exogenous and endogenous dsRNA converge at the RISC complex, which mediates gene silencing effects.
Exogenous dsRNA initiates RNAi by activating the ribonuclease protein dicer,which binds and cleaves doublestranded RNAs dsRNAs to produce doublestranded fragments of 20�25 base pairs with a few unpaired overhang bases on each end.Bioinformatics studies on the genomes of multiple organisms suggest this length maximizes targetgene specificity and minimizes nonspecific effects.These short doublestranded fragments are called small interfering RNAs siRNAs. These siRNAs are then separated into single strands and integrated into an active RISC complex. After integration into the RISC, siRNAs basepair to their target mRNA and induce cleavage of the mRNA, thereby preventing it from being used as a translation template.Exogenous dsRNA is detected and bound by an effector protein known as RDE4 in C. elegans and R2D2 in Drosophila that stimulates dicer activity.This protein only binds long dsRNAs, but the mechanism producing this length specificity is unknown.These RNAbinding proteins then facilitate transfer of cleaved siRNAs to the RISC complex.This initiation pathway may be amplified by the cell through the synthesis of a population of 'secondary' siRNAs using the dicerproduced initiating or 'primary' siRNAs as templates.These siRNAs are structurally distinct from dicerproduced siRNAs and appear to be produced by an RNAdependent RNA polymerase RdRP.
;
MicroRNAs miRNAs are genomically encoded noncoding RNAs that help regulate gene expression, particularly during development. The phenomenon of RNA interference, broadly defined, includes the endogenously induced gene silencing effects of miRNAs as well as silencing triggered by foreign dsRNA. Mature miRNAs are structurally similar to siRNAs produced from exogenous dsRNA, but miRNAs must first undergo extensive posttranscriptional modification. An miRNA is expressed from a much longer RNAcoding gene as a primary transcript known as a primiRNA, which is processed in the cell nucleus to a 70nucleotide stemloop structure called a premiRNA by the microprocessor complex. This complex consists of an RNase III enzyme called Drosha and a dsRNAbinding protein Pasha. The dsRNA portion of this premiRNA is bound and cleaved by dicer to produce the mature miRNA molecule that can be integrated into the RISC complex thus, miRNA and siRNA share the same cellular machinery downstream of their initial processing.The siRNAs derived from long dsRNA precursors differ from miRNAs in that miRNAs, especially those in animals, typically have incomplete base pairing to a target and inhibit the translation of many different mRNAs with similar sequences. In contrast, siRNAs typically basepair perfectly and induce mRNA cleavage only in a single, specific target. In Drosophila and C. elegans, miRNA and siRNA are processed by distinct argonaute proteins and dicer enzymes.Left A fulllength argonaute protein from the archaea species Pyrococcus furiosus. Right The PIWI domain of an argonaute protein in complex with doublestranded RNA.
The basestacking interaction between the 5' base on the guide strand and a conserved tyrosine residue light blue is highlighted the stabilizing divalent cation magnesium is shown as a gray sphere.Left A fulllength argonaute protein from the archaea species Pyrococcus furiosus. Right The PIWI domain of an argonaute protein in complex with doublestranded RNA. The basestacking interaction between the 5' base on the guide strand and a conserved tyrosine residue light blue is highlighted the stabilizing divalent cation magnesium is shown as a gray sphere.
;
The catalyticallyactive components of the RISC complex are endonucleases called argonaute proteins, which cleave the target mRNA strand complementary to their bound siRNA. As the fragments produced by dicer are doublestranded, they could each in theory produce a functional siRNA. However, only one of the two strands, which is known as the guide strand, binds the argonaute protein and directs gene silencing. The other antiguide strand or passenger strand is degraded during RISC activation.Although it was first believed that an ATPdependent helicase separated these two strands, the process is actually ATPindependent and performed directly by the protein components of RISC.The strand selected as the guide tends to be that with a more stable 5' end, but strand selection is unaffected by the direction in which dicer cleaves the dsRNA before RISC incorporation.Instead, the R2D2 protein may serve as the differentiating factor by binding the lessstable 5' end of the passenger strand.The structural basis for binding of RNA to the argonaute protein was examined by Xray crystallography of the binding domain of an RNAbound argonaute protein. Here, the phosphorylated 5' end of the RNA strand enters a conserved basic surface pocket and makes contacts through a divalent cation such as magnesium and by aromatic stacking between the 5' nucleotide in the siRNA and a conserved tyrosine residue. This site is thought to form a nucleation site for the binding of the siRNA to its mRNA target.It is not understood how the activated RISC complex locates complementary mRNAs within the cell.
Although the cleavage process has been proposed to be linked to translation, translation of the mRNA target is not essential for RNAimediated degradation.Indeed, RNAi may be more effective against mRNA targets that are not translated.Argonaute proteins, the catalytic components of RISC, are localized to specific regions in the cytoplasm called Pbodies also cytoplasmic bodies or GW bodies, which are regions with high rates of mRNA decay miRNA activity is also clustered in Pbodies.Disruption of P bodies in cells decreases the efficiency of RNA interference, suggesting that they are the site of a critical step in the RNAi process.Illustration of the major differences between plant and animal gene silencing. Natively expressed microRNA or exogenous small interfering RNA is processed by dicer and integrated into the RISC complex, which mediates gene silencing. In general, miRNAs in plants match perfectly to their gene targets and induce direct messenger RNA cleavage, while miRNAs in animals often have less than perfect base pairing to a number of targets, and induce translational repression.Illustration of the major differences between plant and animal gene silencing. Natively expressed microRNA or exogenous small interfering RNA is processed by dicer and integrated into the RISC complex, which mediates gene silencing. In general, miRNAs in plants match perfectly to their gene targets and induce direct messenger RNA cleavage, while miRNAs in animals often have less than perfect base pairing to a number of targets, and induce translational repression.
;
Organisms vary in their ability to take up foreign dsRNA and use it in the RNAi pathway. The effects of RNA interference can be both systemic and heritable in plants and C. elegans, although not in Drosophila or mammals. In plants, RNAi is thought to propagate by the transfer of siRNAs between cells through plasmodesmata.A broad general distinction between plants and animals lies in the targeting of endogenously produced miRNAs in plants, miRNAs are usually perfectly or nearly perfectly complementary to their target genes and induce direct mRNA cleavage by RISC, while animals' miRNAs tend to be more divergent in sequence and induce translational repression.This translational effect may be produced by inhibiting the interactions of translation initiation factors with the messenger RNA's polyadenine tail.The selective and robust effect of RNAi on gene expression makes it a valuable research tool, both in cell culture and in living organisms synthetic dsRNA introduced into cells can induce suppression of specific genes of interest. RNAi may also be used for largescale screens that systematically shut down each gene in the cell, which can help identify the components necessary for a particular cellular process or an event such as cell division. Exploitation of the pathway is also a promising tool in biotechnology and medicine.
Historically, RNA interference was known by other names, including post transcriptional gene silencing, transgene silencing, and quelling. Only after these apparentlyunrelated processes were fully understood did it become clear that they all described the RNAi phenomenon. RNAi has also been confused with antisense suppression of gene expression, which does not act catalytically to degrade mRNA but instead involves singlestranded RNA fragments physically binding to mRNA and blocking translation. In 2006, Andrew Fire and Craig C. Mello shared the Nobel Prize in Physiology or Medicine for their work on RNA interference in the nematode worm C. elegans, which they published in 1998.Some eukaryotic protozoa such as Leishmania major and Trypanosoma cruzi lack the RNAi pathway entirely.Most or all of the components are also missing in some fungi, most notably the model organism Saccharomyces cerevisiae. Certain ascomycetes and basidiomycetes are also missing RNA interference pathways this observation indicates that proteins required for RNA silencing have been lost independently from many fungal lineages, possibly due to the evolution of a novel pathway with similar function, or to the lack of selective advantage in certain niches.
;
RNA interference is a vital part of the immune response to viruses and other foreign genetic material, especially in plants where it may also prevent selfpropagation by transposons.Plants such as Arabidopsis thaliana express multiple dicer homologs that are specialized to react differently when the plant is exposed to different types of viruses.Even before the RNAi pathway was fully understood, it was known that induced gene silencing in plants could spread throughout the plant in a systemic effect, and could be transferred from stock to scion plants via grafting.This phenomenon has since been recognized as a feature of the plant innate immune system, and allows the entire plant to respond to a virus after an initial localized encounter.In response, many plant viruses have evolved elaborate mechanisms that suppress the RNAi response in plant cells.These include viral proteins that bind short doublestranded RNA fragments with singlestranded overhang ends, such as those produced by the action of dicer.Some plant genomes also express endogenous siRNAs in response to infection by specific types of bacteria.
These effects may be part of a generalized response to pathogens that downregulates any metabolic processes in the host that aid the infection process.Although animals generally express fewer variants of the dicer enzyme than plants, RNAi in some animals has also been shown to produce an antiviral response. In both juvenile and adult Drosophila, RNA interference is important in antiviral innate immunity and is active against pathogens such as Drosophila X virus. A similar role in immunity may operate in C. elegans, as argonaute proteins are upregulated in response to viruses and worms that overexpress components of the RNAi pathway are resistant to viral infection.The role of RNA interference in mammalian innate immunity is poorly understood, and relatively little data is available. However, the existence of viruses that encode genes able to suppress the RNAi response in mammalian cells may be evidence in favour of an RNAidependent mammalian immune response. However, this hypothesis of RNAimediated immunity in mammals has been challenged as poorly substantiated.Alternative functions for RNAi in mammalian viruses also exist, such as miRNAs expressed by the herpes virus that may act as heterochromatin organization triggers to mediate viral latency.
;
Components of the RNA interference pathway are used in many eukaryotes in the maintenance of the organisation and structure of their genomes. Modification of histones and associated induction of heterochromatin formation serves to downregulate genes pretranscriptionally this process is referred to as RNAinduced transcriptional silencing RITS, and is carried out by a complex of proteins called the RITS complex. In fission yeast this complex contains argonaute, a chromodomain protein Chp1, and a protein called Tas3 of unknown function.As a consequence, the induction and spread of heterochromatic regions requires the argonaute and RdRP proteins.Indeed, deletion of these genes in the fission yeast S. pombe disrupts histone methylation and centromere formation causing slow or stalled anaphase during cell division. In some cases, similar processes associated with histone modification have been observed to transcriptionally upregulate genes.The mechanism by which the RITS complex induces heterochromatin formation and organization is not well understood, and most studies have focused on the matingtype region in fission yeast, which may not be representative of activities in other genomic regions or organisms. In maintenance of existing heterochromatin regions, RITS forms a complex with siRNAs complementary to the local genes and stably binds local methylated histones, acting cotranscriptionally to degrade any nascent premRNA transcripts that are initiated by RNA polymerase.
The formation of such a heterochromatin region, though not its maintenance, is dicerdependent, presumably because dicer is required to generate the initial complement of siRNAs that target subsequent transcripts.Heterochromatin maintenance has been suggested to function as a selfreinforcing feedback loop, as new siRNAs are formed from the occasional nascent transcripts by RdRP for incorporation into local RITS complexes.The relevance of observations from fission yeast matingtype regions and centromeres to mammals is not clear, as heterochromatin maintenance in mammalian cells may be independent of the components of the RNAi pathway.Endogenously expressed miRNAs, including both intronic and intergenic miRNAs, are most important in translational repression and in the regulation of development, especially the timing of morphogenesis and the maintenance of undifferentiated or incompletely differentiated cell types such as stem cells.The role of endogenously expressed miRNA in downregulating gene expression was first described in C. elegans in 1993.In plants this function was discovered when the JAW microRNA of Arabidopsis was shown to be involved in the regulation of several genes that control plant shape.In plants, the majority of genes regulated by miRNAs are transcription factors thus miRNA activity is particularly wideranging and regulates entire gene networks during development by modulating the expression of key regulatory genes, including transcription factors as well as Fbox proteins. In many organisms, including humans, miRNAs have also been linked to the formation of tumors and dysregulation of the cell cycle. Here, miRNAs can function as both oncogenes and tumor suppressors.RNA sequences siRNA or miRNA that are complementary to parts of a promotor can increase gene transcription, a phenomenon dubbed RNA activation. As in gene silencing by RNA interference, Dicer and RISC are involved, but the mechanism for how RISC upregulates a gene is not known.
;
The type of RNA editing that is most prevalent in higher eukaryotes converts adenosine nucleotides into inosine in dsRNAs via the enzyme adenosine deaminase ADAR.It was originally proposed in 2000 that the RNAi and A?I RNA editing pathways might compete for a common dsRNA substrate.Indeed, some premiRNAs do undergo A?I RNA editing,and this mechanism may regulate the processing and expression of mature miRNAs. Furthermore, at least one mammalian ADAR can sequester siRNAs from RNAi pathway components.Further support for this model comes from studies on ADARnull C. elegans strains indicating that A?I RNA editing may counteract RNAi silencing of endogenous genes and transgenes.Gene expression in prokaryotes is influenced by an RNAbased system similar in some respects to RNAi. Here, RNAencoding genes control mRNA abundance or translation by producing a complementary RNA that binds to an mRNA by base pairing. However these regulatory RNAs are not generally considered to be analogous to miRNAs because the dicer enzyme is not involved.It has been suggested that CRISPR systems in prokaryotes are analogous to eukaryotic RNA interference systems, although none of the protein components are orthologous.Based on parsimonybased phylogenetic analysis, the most recent common ancestor of all eukaryotes most likely already possessed an early RNA interference pathway the absence of the pathway in certain eukaryotes is thought to be a derived characteristic. The ancestral RNAi system probably contained at least one dicerlike protein, one argonaute, one PIWI protein, and an RNA dependent RNA polymerase that may have also played other cellular roles.
A largescale comparative genomics study likewise indicates that the eukaryotic crown group already possessed these components, which may then have had closer functional associations with generalized RNA degradation systems such as the exosome.This study also suggests that the RNAbinding argonaute protein family, which is shared among eukaryotes, most archaea, and at least some bacteria such as Aquifex aeolicus, is homologous to and originally evolved from components of the translation initiation system.The ancestral function of the RNAi system is generally agreed to have been immune defense against exogenous genetic elements such as transposons and viral genomes.Related functions such as histone modification may have already been present in the ancestor of modern eukaryotes, although other functions such as regulation of development by miRNA are thought to have evolved later.RNA interference genes, as components of the antiviral innate immune system in many eukaryotes, are involved in an evolutionary arms race with viral genes. Some viruses have evolved mechanisms for suppressing the RNAi response in their host cells, an effect that has been noted particularly for plant viruses. Studies of evolutionary rates in Drosophila have shown that genes in the RNAi pathway are subject to strong directional selection and are among the fastestevolving genes in the Drosophila genome.
;
A normal adult fruit fly Drosophila melanogaster, a common model organism used in RNAi experiments.A wildtype adult Caenorhabditis elegans nematode worm, grown under RNAi suppression of a nuclear hormone receptor involved in desaturase regulation. These worms have abnormal fatty acid metabolism but are viable and fertile.A wildtype adult Caenorhabditis elegans nematode worm, grown under RNAi suppression of a nuclear hormone receptor involved in desaturase regulation. These worms have abnormal fatty acid metabolism but are viable and fertile.The RNA interference pathway is often exploited in experimental biology to study the function of genes in cell culture and in vivo in model organisms. Doublestranded RNA is synthesized with a sequence complementary to a gene of interest and introduced into a cell or organism, where it is recognized as exogenous genetic material and activates the RNAi pathway. Using this mechanism, researchers can cause a drastic decrease in the expression of a targeted gene. Studying the effects of this decrease can show the physiological role of the gene product. Since RNAi may not totally abolish expression of the gene, this technique is sometimes referred as a knockdown, to distinguish it from knockout procedures in which expression of a gene is entirely eliminated.
Extensive efforts in computational biology have been directed toward the design of successful dsRNA reagents that maximize gene knockdown but minimize offtarget effects. Offtarget effects arise when an introduced RNA has a base sequence that can pair with and thus reduce the expression of multiple genes at a time. Such problems occur more frequently when the dsRNA contains repetitive sequences. It has been estimated from studying the genomes of H. sapiens, C. elegans, and S. pombe that about 10% of possible siRNAs will have substantial offtarget effects.A multitude of software tools have been developed implementing algorithms for the design of general, mammalspecific, and virusspecific siRNAs that are automatically checked for possible crossreactivity.Depending on the organism and experimental system, the exogenous RNA may be a long strand designed to be cleaved by dicer, or short RNAs designed to serve as siRNA substrates. In most mammalian cells, shorter RNAs are used because long doublestranded RNA molecules induce the mammalian interferon response, a form of innate immunity that reacts nonspecifically to foreign genetic material. Mouse oocytes and cells from early mouse embryos lack this reaction to exogenous dsRNA and are therefore a common model system for studying geneknockdown effects in mammals.Specialized laboratory techniques have also been developed to improve the utility of RNAi in mammalian systems by avoiding the direct introduction of siRNA, for example, by stable transfection with a plasmid encoding the appropriate sequence from which siRNAs can be transcribed,or by more elaborate lentiviral vector systems allowing the inducible activation or deactivation of transcription, known as conditional RNAi.
;
Most functional genomics applications of RNAi in animals have used C. elegans and D. melanogaster, as these are the common model organisms in which RNAi is most effective. C. elegans is particularly useful for RNAi research for two reasons firstly, the effects of the gene silencing are generally heritable, and secondly because delivery of the dsRNA is extremely simple. Through a mechanism whose details are poorly understood, bacteria such as E. coli that carry the desired dsRNA can be fed to the worms and will transfer their RNA payload to the worm via the intestinal tract. This delivery by feeding is just as effective at inducing gene silencing as more costly and timeconsuming delivery methods, such as soaking the worms in dsRNA solution and injecting dsRNA into the gonads.Although delivery is more difficult in most other organisms, efforts are also underway to undertake largescale genomic screening applications in cell culture with mammalian cells.Approaches to the design of genomewide RNAi libraries can require more sophistication than the design of a single siRNA for a defined set of experimental conditions.
Artificial neural networks are frequently used to design siRNA libraries and to predict their likely efficiency at gene knockdown.Mass genomic screening is widely seen as a promising method for genome annotation and has triggered the development of highthroughput screening methods based on microarrays. However, the utility of these screens and the ability of techniques developed on model organisms to generalize to even closelyrelated species has been questioned, for example from C. elegans to related parasitic nematodes.Functional genomics using RNAi is a particularly attractive technique for genomic mapping and annotation in plants because many plants are polyploid, which presents substantial challenges for more traditional genetic engineering methods. For example, RNAi has been successfully used for functional genomics studies in the hexaploid wheat Triticum aestivum, as well as more common plant model systems Arabidopsis thaliana and Zea mays.
;
It may be possible to exploit RNA interference in therapy. Although it is difficult to introduce long dsRNA strands into mammalian cells due to the interferon response, the use of short interfering RNA mimics has been more successful.Among the first applications to reach clinical trials were in the treatment of macular degeneration and respiratory syncytial virus,RNAi has also been shown to be effective in the reversal of induced liver failure in mouse models.Other proposed clinical uses center on antiviral therapies, including the inhibition of viral gene expression in cancerous cells, knockdown of host receptors and coreceptors for HIV, the silencing of hepatitis A and hepatitis B genes, silencing of influenza gene expression,and inhibition of measles viral replication. Potential treatments for neurodegenerative diseases have also been proposed, with particular attention being paid to the polyglutamine diseases such as Huntington's disease.RNA interference is also often seen as a promising way to treat cancer by silencing genes differentially upregulated in tumor cells or genes involved in cell division. A key area of research in the use of RNAi for clinical applications is the development of a safe delivery method, which to date has involved mainly viral vector systems similar to those suggested for gene therapy.Despite the proliferation of promising cell culture studies for RNAibased drugs, some concern has been raised regarding the safety of RNA interference, especially the potential for offtarget effects in which a gene with a coincidentally similar sequence to the targeted gene is also repressed.
A computational genomics study estimated that the error rate of offtarget interactions is about 10%.One major study of liver disease in mice led to high death rates in the experimental animals, suggested by researchers to be the result of oversaturation of the dsRNA pathway.RNA interference has been used for applications in biotechnology, particularly in the engineering of food plants that produce lower levels of natural plant toxins. Such techniques take advantage of the stable and heritable RNAi phenotype in plant stocks. For example, cotton seeds are rich in dietary protein but naturally contain the toxic terpenoid product gossypol, making them unsuitable for human consumption. RNAi has been used to produce cotton stocks whose seeds contain reduced levels of deltacadinene synthase, a key enzyme in gossypol production, without affecting the enzyme's production in other parts of the plant, where gossypol is important in preventing damage from plant pests.Similar efforts have been directed toward the reduction of the cyanogenic natural product linamarin in cassava plants.Although no plant products that use RNAibased genetic engineering have yet passed the experimental stage, development efforts have successfully reduced the levels of allergens in tomato plants and decreased the precursors of likely carcinogens in tobacco plants.Other plant traits that have been engineered in the laboratory include the production of nonnarcotic natural products by the opium poppy, resistance to common plant viruses,and fortification of plants such as tomatoes with dietary antioxidants.Previous commercial products, including the Flavr Savr tomato and two cultivars of ringspotresistant papaya, were originally developed using antisense technology but likely exploited the RNAi pathway.
;
Example petunia plants in which genes for pigmentation are silenced by RNAi. The left plant is wildtype the right plants contain transgenes that induce suppression of both transgene and endogenous gene expression, giving rise to the unpigmented white areas of the flower.The discovery of RNAi was preceded first by observations of transcriptional inhibition by antisense RNA expressed in transgenic plants and more directly by reports of unexpected outcomes in experiments performed by plant scientists in the U.S. and The Netherlands in the early 1990s.In an attempt to alter flower colors in petunias, researchers introduced additional copies of a gene encoding chalcone synthase, a key enzyme for flower pigmentation into petunia plants of normally pink or violet flower color. The overexpressed gene was expected to result in darker flowers, but instead produced less pigmented, fully or partially white flowers, indicating that the activity of chalcone synthase had been substantially decreased in fact, both the endogenous genes and the transgenes were downregulated in the white flowers. Soon after, a related event termed quelling was noted in the fungus Neurospora crassa,although it was not immediately recognized as related. Further investigation of the phenomenon in plants indicated that the downregulation was due to posttranscriptional inhibition of gene expression via an increased rate of mRNA degradation.This phenomenon was called cosuppression of gene expression, but the molecular mechanism remained unknown.Not long after, plant virologists working on improving plant resistance to viral diseases observed a similar unexpected phenomenon.
While it was known that plants expressing virusspecific proteins showed enhanced tolerance or resistance to viral infection, it was not expected that plants carrying only short, noncoding regions of viral RNA sequences would show similar levels of protection. Researchers believed that viral RNA produced by transgenes could also inhibit viral replication.The reverse experiment, in which short sequences of plant genes were introduced into viruses, showed that the targeted gene was suppressed in an infected plant. This phenomenon was labeled virusinduced gene silencing VIGS, and the set of such phenomena were collectively called post transcriptional gene silencing.After these initial observations in plants, many laboratories around the world searched for the occurrence of this phenomenon in other organisms. Craig C. Mello and Andrew Fire's 1998 Nature paper reported a potent gene silencing effect after injecting double stranded RNA into C. elegans.In investigating the regulation of muscle protein production, they observed that neither mRNA nor antisense RNA injections had an effect on protein production, but doublestranded RNA successfully silenced the targeted gene. As a result of this work, they coined the term RNAi. Fire and Mello's discovery was particularly notable because it represented the first identification of the causative agent of a previously inexplicable phenomenon. Fire and Mello were awarded the Nobel Prize in Physiology or Medicine in 2006 for their work.