Resultados
Retornar a Home Page
Tránsito Turístico
El análisis de tránsito turístico reveló que CAPH y TRC tuvieron el nivel de tránsito
más alto y bajo, respectivamente (Figura 3.2). Los dos albergues con los niveles de
tránsito más altos (CAPH y EI) a su vez son también aquellos albergues que han estado
operando por más tiempo que cualquier otro.
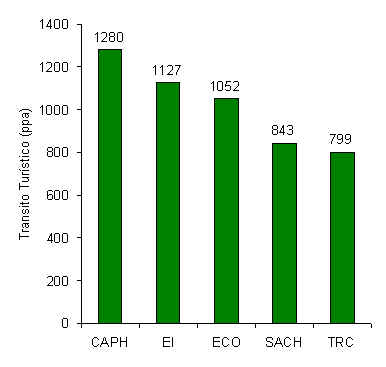
Figura 3.2. Intensidad de tránsito promedio a lo largo de los T, en términos de personas
por año (ppa), para la época 1994-1998. [Los datos no son iguales a los arribos anuales
a cada albergue.]
Hábitat y el Recurso Fruta
Cinco géneros dominaron las muestras (en orden de abundancia
decreciente): Pona* (Figura 3.4), Huicunguo* (Figura 3.6), Chimicua (Figura 3.7), Shebon*
(Figura 3.5) y Ficus* (Figura 3.8). Cuatro de estos (*) a su vez son recursos
"claves" en términos de fruta. Estos géneros representaron entre 70-90% de los
árboles muestreados, aunque la Pona era facilmente la más abundante entre 41-82% de las
muestras en cada albergue. La densidad total de árboles frutales, agrupando los
resultados de T y C, era mayor en el TRC (86 árboles/hectárea) y menor en el CAPH (39
árboles/hectárea)(Figura 3.3).
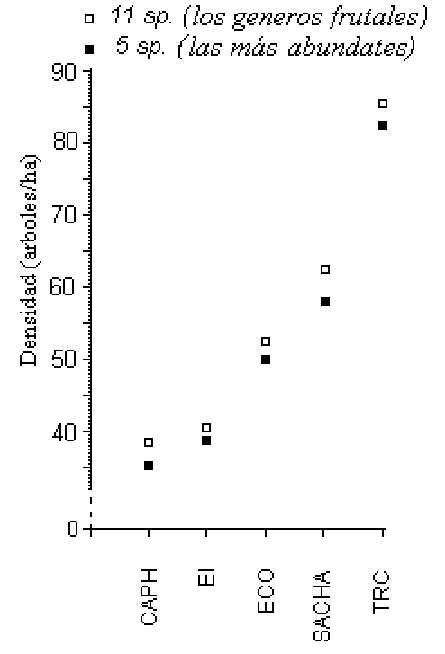
Figura 3.3. El patrón entre albergues en términos de densidad promedio acumulado de los
11 géneros de árboles frutales (cuadros blancos) y los 5 géneros más abundantes
(cuadros negros).
Pona (Iriartea)
Esta palmera es consistentemente la más abundante de todos los
árboles muestreados. Pruebas-t revelaron que sólo en EI hubo una diferencia en densidad
entre T y C (T = 28 árboles/hectárea; C = 21 árboles/hecttárea; t = 2.16, p = 0.05)
(Figura 3.4). Los albergues con la mayor y menor densidad de esta palmera fueron TRC (69
árboles/hectárea) y CAPH (14 árboles/hectáreas), respectivamente.
Shebon (Scheelea)
Pruebas-t revelaron que en EI, ECO y TRC hubo una diferencia
significativa en la densidad de esta palmera entre T y C, aunque no hubo un patrón
consistente entre ellos para sugerir si el turismo tenia algo que ver en la abundancia
(Figura 3.5). Los albergues con la mayor y menor densidad eran ECO (9 árboles/hectárea)
y TRC (0.5 árboles/hectárea), respectivamente. En TRC hubo una densidad
significativamente menor que en otros albergues, aunque la densidad promedio entre los
demás albergues no era diferente.
Huicungo (Astrocaryum)
Pruebas-t revelaron que en CAPH, ECO, SACHA y TRC hubo una diferencia
significativa en la densidad de este árbol entre T y C, aunque no hubo un patrón
consistente entre ellos para sugerir si el turismo tenia algo que ver en la abundancia
(Figura 3.6). Los albergues con la mayor y menor densidad eran ECO (13 árboles/hectárea)
y EI (2 árboles/hectárea), respectivamente. En el caso de EI la densidad fue
significativamente menor que en otros albergues, aunque la densidad promedio entre los
demás albergues no era diferente.
Chimicua (Pseudolmedia)
Pruebas-t revelaron que en CAPH, SACHA y TRC hubo una diferencia
significativa en la densidad de este árbol entre T y C, aunque no hubo un patrón
consistente entre ellos para sugerir si el turismo tenia algo que ver en la abundancia
(Figura 3.7). Los albergues con la mayor y menor densidad eran ECO (8 árboles/hectárea)
y TRC (4 árboles/hectárea), respectivamente. La densidad entre ECO y SACHA no era
significativamente diferente aunque estos dos tuvieron densidades más altos que los otros
3 albergues.
Ficus
Pruebas-t mostraron que sólo en ECO hubo una diferencia significativa
en densidad entre T y C (T = 3.4 árboles/hectárea; C = 2.2 árboles/hecttárea; t =
2.19, p < 0.03)(Figura 3.8). Los albergues con la mayor y menor densidad de esta
palmera eran ECO (2.8 árboles/hectárea) y EI (1.8 árboles/hectáreas), respectivamente.
Siguientes pruebas mostraron que en general no hubo ninguna diferencia en la densidad de
este género entre albergues.
Bambú (Guadua)
El bambú era totalmente aucente de los dos albergues ubicados a
orillas del río Madre de Dios (CAPH y ECO), y era relativamente común por el río
Tambopata, aunque era más abundante alrededor de SACHA donde hubo una mayor incidencia en
C que en T. No hubo una diferencia en cuanto a la densidad relativa de esta planta entre
SACHA y TRC, pero ambos tuvieron una abundancia más alta que la de EI.
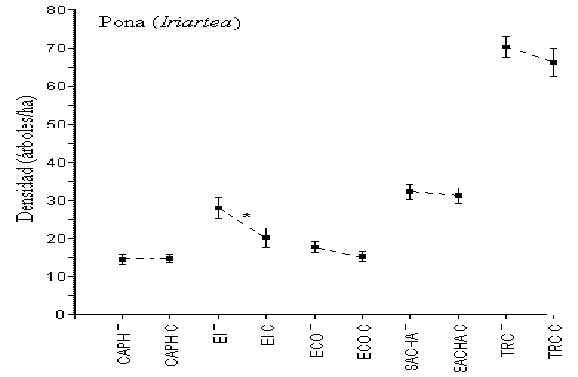
Figura 3.4. Densidad de Pona (Iriartea); * Diferencia significativa entre T y C.
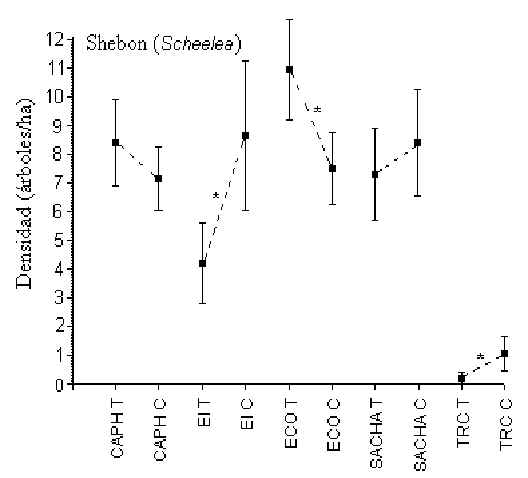
Figura 3.5. Densidad del Shebon (Scheelea); * Diferencia significativa entre T y C.
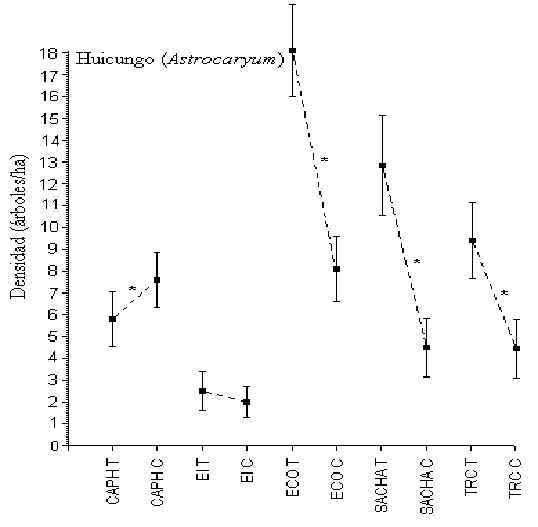
Figura 3.6. Densidad del Huicungo (Astrocaryum); * Diferencia significativa entre T y C.
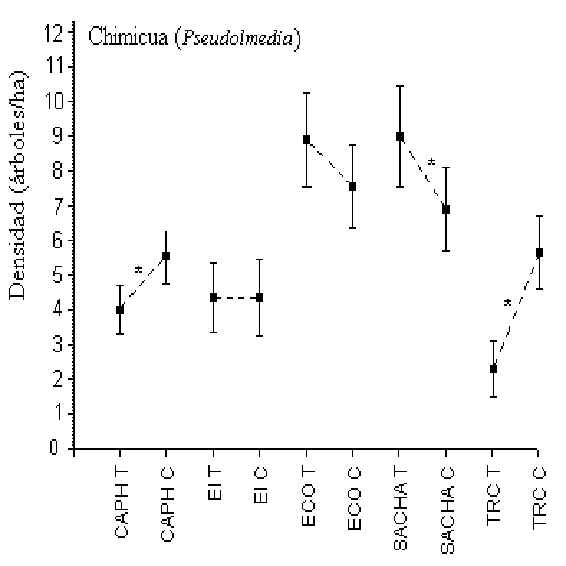
Figura 3.7. Densidad de la Chimicua (Pseudolmedia); * Diferencia significativa entre T y
C.
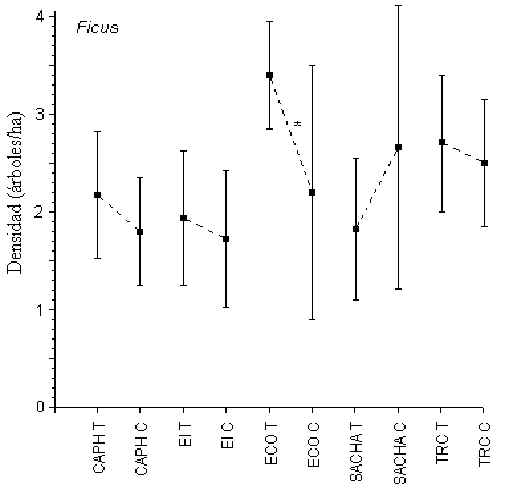
Figura 3.8. Densidad de Ficus; * Diferencia significativa entre T y C.
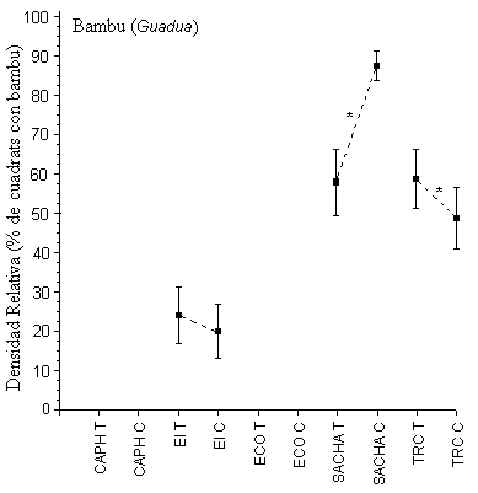
Figura 3.9. Densidad relativa del bambú (Guadua); * Diferencia significativa entre T y C.
Un Análisis de Conjuntos Jerárquicos, utilizando los datos de los 13 géneros de manera
simultanea (Figura 3.10), mostró que hubo gran similitud entre albergues que eran
geográficamente cercanos, un resultado esperado debido a que localidades más cercanos
probablemente comparten características edáficas (suelo), climatológicas y
antropomórficas, los cuales afectan la florística. Aunque más de 50% del patrón es
debido a la dominancia de un sólo árbol, la Pona (Iriartea). El agrupamiento de los
cuatro albergues en las zonas de colonización humana muestra una discrepancia grande con
TRC . Esto es probablemente una manifestación de la presión antrópica a través de los
años en esta zona. Además el clima podria haber afectado la distribución ya que la zona
alrededor de TRC es más húmedo que los otros albergues debido a su cercania a la
cordillera de los Andes.
ECO ---+
+-----+
CAPH ---+ +
+-----------------------------------------+
EI -----+ + +
+---+ +
SACHA-----+ +----------
+
TRC ---------------------------------------------------+
Figura 3.10. Análisis multivariado de la similitud entre albergues, utilizando el
Análisis de Conjuntos Jerárquicos con Distancias de Cuerda basadas en Distancias Simples
Eucledianas.
Presión de Caza
Tanto el número de cartuchos encontrados como el de encuentros con personas armadas
revela que tres de los albergues (SACHA, CAPH y ECO) tienen influencia de caza, aunque en
ECO no parece ser muy fuerte (Tabla 3.2). Un análisis de correlación entre el índice de
caza y la abundancia relatva de cinco especies frecuentemente perseguidos por cazadores en
Madre de Dios muestra una relación negativa (Tabla 3.3). Los datos recolectados sobre la
abundancia de la pava pucacunga (Penelope jacquacu), cracido grande y perseguido
frecuentemente por cazadores y a su vez muy sensible a la mínima presión de caza (Peres
1990, Robinson et al. 1991, Silva & Strahl 1991), refleja nuestra conclusión (Figura
3.11), y los resultados de Ascorra (1997) muestran que SACHA definitivamente tiene una
alta presión de caza. Es interesante que en EI no se encontró señales de caza, apesar
de la relativa cercania a dos comunidades, una nativa ubicada al Este del sistema de
trochas y otra mestiza al Oeste del albergue. Pareciera que durante los últimos 20 años
de la existencia del albergue estas comunidades han llegado a respetar el área alrededor
como una mini-reserva turística.
Tabla 3.2. Variables de caza monitoereados e índice proporcional de la presión de caza
en los albergues.
| Albergue | No. de personas encontrados con escopetas (90 días) |
Cartuchos encontrados (1=Sí, 0=No) |
Índice de presión de caza |
| SACHA | 5 |
1 |
6 |
| CAPH | 4 |
1 |
5 |
| ECO | 1 |
1 |
2 |
| EI | 0 |
0 |
0 |
| TRC | 0 |
0 |
0 |
Tabla 3.3. Correlación de Spearman entre el índice de caza y el índice de abundancia
de cinco especies más perseguidas normalmente por cazadores en Madre de Dios: Tayassu
tajacu, Tapirus terrestris, Mazama americana, Alouatta seniculus, Cebus apella. Abundancia
fue medido relativamente Raiz(IKA+1). * p < 0.05.
| Albergue | Índice de presión de caza |
Abundancia de 5 especies de mamífero |
Spearman |
| SACHA | 6 |
5,160 |
- 0.87 * |
| CAPH | 5 |
5,102 |
. |
| ECO | 2 |
5,357 |
. |
| EI | 0 |
5,378 |
. |
| TRC | 0 |
5,725 |
. |
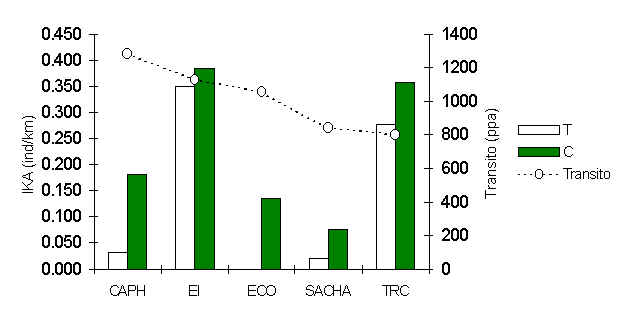
Figura 3.11. Abundancia relativa (IKA) de la pava pucacunga (Penelope jacquacu).
Esfuerzo de Muestreo
El esfuerzo de muestreo entre los T y C en cada albergues era muy similar. Sin embargo,
hubo diferencias significativas entre albergues en cuanto al esfuerzo total (T + C), los
extremos siendo 172 km (EI) y 315 km (CAPH), o una diferencia de 45% (Tabla 3.4). Las
diferencias fueron resultado de una combinación de factores; 1) la variación en la
longitud total de los transectos en cada albergue, en EI esta distancia era reducida, y 2)
el hecho que en TRC sólo se muestreo en 5 ocasiones y no 6 como originalmente se
planificó. El esfuerzo total durante los 23 meses del estudio fue de 1,274 km. En
relación con otros estudios sobre abundancia de mamíferos selváticos esta cifra es
relativamente alta. La velocidad promedio de los muestreos fue de 1.11 km/hr, que es muy
cerca de la óptima de 1 km/hr.
Tabla 3.4. Esfuerzo de muestreo por albergue.
| Albergue | Turístico (T) km |
Control (C ) km |
Total km |
| CAPH | 156 |
159 |
315 |
| EI | 86 |
86 |
172 |
| ECO | 155 |
155 |
310 |
| SACHA | 151 |
145 |
296 |
| TRC | 97 |
84 |
181 |
| Total | 645 |
629 |
1,274 |
Diversidad de Especies
Se registraron entre 21 - 27 especies en cada albergue, tanto visualmente como a través
de huellas utilizando datos recolectados durante los censos por transecto y durante
encuentros fortuitos en difrentes ocasiones (Figura 3.12). En total se registraron 31
especies durante los 23 meses del estudio. No hubo una diferencia significativa en cuanto
a la riqueza de especies entre T y C en ningun albergue. Aunque se encontro, a través de
un análisis de correlación que la riqueza general por albergue esta relacionada
fuertemente con la presión de caza (Spearman = -0.9, p < 0.05).
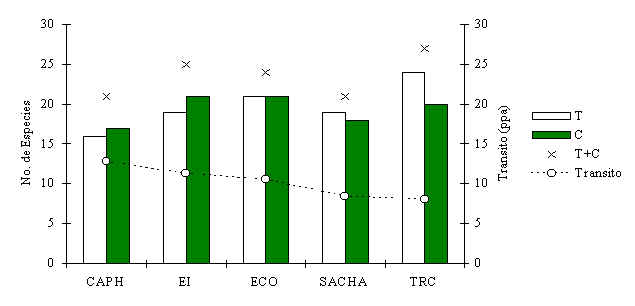
Figura 3.12. Riqueza de especies, determinada a través de encuentros visuales y
registros de huellas.
Número de Individuos por Grupo
Las estimaciones sobre el número promedio de individuos por grupo fueron calculados a
través de los datos de conteos completos de cada especie, tanto durante los censos así
como encuentros fortuitos. Se omitieron de este cálculo los conteos incompletos. La
especie con el mayor tamaño de grupo fue el Fraile (Saimiri boliviensis) que alcanzó un
máximo de 85 individuos en ECO. Los análises de pruebas-t revelaron que no hubo una
diferencia significativa entre T y C en cuanto a tamaño de grupo de ninguna de las
especies. Sin embargo, el mismo tipo de análisis, utilizando datos agrupados de T y C,
reveló que el Pichico (S. fuscicollis)(Figura 3.13; Tabla 3.5) y el Machin Negro (C.
apella)(Figura 3.14; Tabla 3.6) variaron significativamente entre algunos albergues. Un
análisis de correlación de Pearson encontró que el tamaño de grupo del Pichico era
positivamente relacionado con la magnitud de tránsito (Pearson = +0.92, p < 0.05),
mientras el tamaño de grupo del Machin Negro era negativamente relacionado con la
presión de caza (Pearson = -0.88, p < 0.05). En general el tamaño de grupo promedio
de muchas de las espcies estudiadas es muy similar a los que caraterizan otras zonas de
Madre de Dios (Janzen et al. 1980, Kiltie 1981, Freese et al. 1982, Terborgh 1983, Ascorra
1997)(Tabla 3.7).
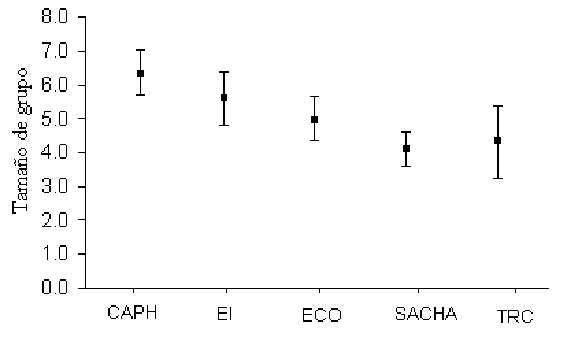
Figura 3.13. La variación en tamaño de grupo entre albergues para el Pichico (S.
fuscicollis).
Tabla 3.5. Probabilidades derivados de pruebas-t sobre el tamaño de grupo del Pichico. *
p < 0.05; ** p < 0.01.
| . | CAPH |
EI |
ECO |
SACHA |
| EI | 0.06 |
. | . | . |
| ECO | 0.00** |
0.15 |
. | . |
| SACHA | 0.00** |
0.00** |
0.02* |
. |
| TRC | 0.00** |
0.04* |
0.15 |
0.36 |
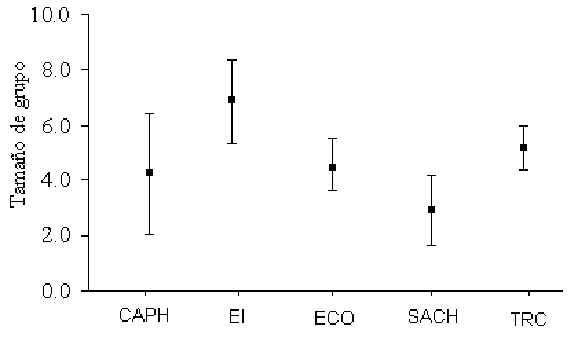
Figura 3.14. La variación en tamaño de grupo entre albergues para el Machin Negro (C.
apella).
Tabla 3.6. Probabilidades derivadas de pruebas-t sobre el tamaño de grupo del Machin
Negro. * p < 0.05; ** p < 0.01.
| . | CAPH | EI | ECO | SACHA |
| EI | 0.05* | . | . | . |
| ECO | 0.41 | 0.01* | . | . |
| SACHA | 0.16 | 0.00** | 0.02* | . |
| TRC | 0.25 | 0.03* | 0.18 | 0.00** |
Tabla 3.7. Resúmen del tamaño de grupo para 13 especies, y comparaciones bibliográficas
para 7 de ellas. 1 Freese et al. (1982), 2 Kiltie et al. (1983), 3 Warner (1999).
| Especies | . | Tamaño de Grupo | 95% IC |
Tamaño Grupo |
|||
| . | . | N |
Min. |
Max. |
Prom. |
. | . |
| Southern Tamandua | T. tetradactyla | 15 |
1 |
2 |
1.10 |
+/- 0.10 |
. |
| Saddleback Tamarin | S. fuscicollis | 288 |
1 |
16 |
5.40 |
+/- 2.20 |
6, 7 (1, 3) |
| Squirrel Monkey | S. boliviensis | 57 |
1 |
85 |
28.60 |
+/- 5.32 |
40, 18 (1, 3) |
| Dusky titi Monkey | C. brunneus | 44 |
1 |
6 |
2.70 |
+/- 1.90 |
3, 3 (1, 3) |
| Red Howler | A. seniculus | 54 |
1 |
10 |
4.00 |
+/- 0.60 |
5 (1) |
| White-fronted Capuchin | C. albifrons | 8 |
1 |
32 |
12.5 |
+/- 8.30 |
. |
| Brown capuchin | C. apella | 109 |
1 |
12 |
4.70 |
+/- 1.10 |
10, 6 (1, 3) |
| Black Spider Monkey | A. paniscus | 34 |
1 |
16 |
5.20 |
+/- 1.30 |
7 (1) |
| South American Coati | N. nasua | 25 |
1 |
15 |
5.40 |
+/- 1.80 |
. |
| Collared Peccary | T. tajacu | 75 |
1 |
9 |
2.00 |
+/- 0.30 |
2 (2) |
| Bolivian Squirrel | S. ignitus | 118 |
1 |
6 |
1.14 |
+/- 0.10 |
. |
| Southern Amazon Red Squirrel | S. spadiceus | 672 |
1 |
5 |
1.15 |
+/- 0.04 |
. |
| Brown Agouti | D. variegata | 364 |
1 |
3 |
1.10 |
+/- 0.03 |
. |
Número de Encuentros e Índices de Abundancia
Durante 23 meses de censos a lo largo de los transectos T y C hubo un total de 1,328
encuentros con mamíferos de 26 especies (Tabla 3.8). Estos datos se convirtieron en
Indices Kilométricos de Abundancia (IKA) por especie en terminos de grupos/km (Tabla
3.9), y después del análisis de tamaño de grupo en terminos de individuos/km (Tabla
3.10). Estos datos se utilizaron para analizar los impactos de los vairables de hábitat,
presión de caza y tránsito. En general, las tres especies más comunes, en términos de
encuentros y grupos/km fueron la Ardilla Roja, el Pichico, y el Añuje, aunque las tres
especies más comunes en términos de individuos/km fueron el Fraile, el Pichico y la
Ardilla Roja.
Densidad
La determinación de la densidad absoluta de una especie requiere de una estimación del
área boscosa efectivamente muestreada. Esta área es un factor de la distancia
muestreada, en km, y la distancia de detección perpendicular específica de una especie
(Psp). La variable Psp proviene de un análisis de todas las estimaciones de detección
perpendicular para una especie a través de un programa computarizada llamada
"Distance" (Laake et al. 1991). Una explicación más precisa de los parámetros
utilizados, de la teoría de esta ecuación y la determinación de Psp para una especie se
encuentra en Buckland et al. (1993). Primero, la detección perpendicular de cada
encuentro con cada especie se estimó con la Ecuación 3.1, donde; P1 = Distancia de
detección perpendicular de un encuentro (km); S = Distancia de detección
observador-animal (km); G = Ancho de Grupo (km); y A = Angulo transecto-animal (radianes)
(ver Figura 3.1). En el caso de encuentros con animales solitarios G es cero.
Ecuación 3.1: P1 = (S+0.5G) x Sin(A);
Tabla 3.8. Número de encuentros con 26 especies a lo largo de T y C en cada albergue.
| Albergues | CAPH |
EI |
ECO |
SACHA |
TRC |
Total |
|||||||||||
| Tipo de Transecto | T |
C |
T |
C |
T |
C |
T |
C |
T |
C |
. | ||||||
| Km Censados | 156 |
159 |
86 |
86 |
155 |
155 |
151 |
145 |
97 |
84 |
1,274 |
||||||
| Xenarthra | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Myrmecophaga tridactyla | - |
1 |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
||||||
| Tamandua tetradactyla | - |
- |
1 |
- |
4 |
4 |
4 |
- |
1 |
- |
14 |
||||||
| Bradypus variegata | - |
- |
- |
1 |
- |
- |
- |
- |
2 |
- |
3 |
||||||
| Dasypus sp. | - |
- |
- |
1 |
- |
- |
- |
- |
- |
- |
1 |
||||||
| Primates | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Saguinus fuscicollis | 47 |
25 |
10 |
7 |
31 |
22 |
10 |
12 |
4 |
8 |
176 |
||||||
| Saimiri boliviensis | 1 |
6 |
- |
1 |
11 |
5 |
8 |
9 |
13 |
3 |
57 |
||||||
| Aotus sp. | - |
- |
- |
1 |
- |
1 |
- |
13 |
1 |
- |
16 |
||||||
| Callicebus brunneus | 3 |
- |
4 |
2 |
- |
- |
1 |
- |
1 |
10 |
21 |
||||||
| Alouatta seniculus | 1 |
- |
6 |
1 |
4 |
10 |
3 |
1 |
8 |
2 |
36 |
||||||
| Cebus albifrons | - |
- |
- |
- |
1 |
4 |
- |
- |
- |
- |
5 |
||||||
| Cebus apella | - |
5 |
4 |
10 |
11 |
14 |
1 |
6 |
16 |
11 |
78 |
||||||
| Ateles paniscus | - |
- |
- |
- |
- |
- |
- |
- |
6 |
13 |
19 |
||||||
| Carnivora | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Atelocynus microtis | - |
- |
- |
- |
- |
- |
- |
- |
1 |
- |
1 |
||||||
| Nasua nasua | 1 |
- |
- |
1 |
2 |
3 |
- |
4 |
1 |
- |
12 |
||||||
| Eira barbara | 3 |
1 |
2 |
2 |
2 |
3 |
1 |
8 |
4 |
2 |
28 |
||||||
| Gallictis vitata | - |
- |
- |
- |
1 |
- |
- |
- |
- |
- |
1 |
||||||
| Lutra longicauda | - |
- |
- |
- |
- |
- |
- |
- |
- |
1 |
1 |
||||||
| Leopardus pardalis | 1 |
- |
- |
- |
- |
- |
2 |
- |
1 |
- |
4 |
||||||
| Perissodactyla | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Tapirus terrestris | - |
- |
- |
- |
- |
2 |
- |
- |
1 |
2 |
5 |
||||||
| Artiodactyla | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Tayassu pecari | - |
- |
- |
- |
- |
- |
- |
1 |
1 |
- |
2 |
||||||
| Tayassu tajacu | 3 |
7 |
8 |
7 |
- |
4 |
4 |
9 |
4 |
1 |
47 |
||||||
| Mazama americana | - |
3 |
- |
3 |
- |
5 |
2 |
5 |
1 |
1 |
20 |
||||||
| Rodentia | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Sciurus ignitus | 14 |
17 |
6 |
3 |
4 |
2 |
13 |
9 |
16 |
8 |
92 |
||||||
| Sciurus spadiceus | 110 |
87 |
35 |
18 |
79 |
58 |
36 |
44 |
19 |
30 |
516 |
||||||
| Dasyprocta variegata | 43 |
20 |
8 |
12 |
6 |
20 |
27 |
26 |
4 |
3 |
169 |
||||||
| Myoprocta pratti | - |
1 |
- |
1 |
1 |
- |
- |
- |
- |
- |
3 |
||||||
| Total | 227 |
173 |
84 |
71 |
157 |
157 |
112 |
147 |
105 |
95 |
|||||||
| T+C | 400 |
155 |
314 |
259 |
200 |
1,328 |
|||||||||||
Tabla 3.9. IKA en términos de grupos/km de 26 especies.
| Albergues | CAPH |
EI |
ECO |
SACHA |
TRC |
Prom (x10-2) |
|||||||||||
| Tipo de Transecto | T |
C |
T |
C |
T |
C |
T |
C |
T |
C |
. | ||||||
| Km Censados | 156 |
159 |
86 |
86 |
155 |
155 |
151 |
145 |
97 |
84 |
. | ||||||
| Xenarthra | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Myrmecophaga tridactyla | - |
0.012 |
- |
- |
- |
- |
- |
- |
- |
- |
0.10 |
||||||
| Tamandua tetradactyla | - |
- |
0.012 |
- |
0.026 |
0.026 |
0.026 |
- |
0.010 |
- |
1.00 |
||||||
| Bradypus variegata | - |
- |
- |
0.012 |
- |
- |
- |
- |
0.021 |
- |
0.03 |
||||||
| Dasypus sp. | - |
- |
- |
0.012 |
- |
- |
- |
- |
- |
- |
0.10 |
||||||
| Primates | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Saguinus fuscicollis | 0.301 |
0.157 |
0.116 |
0.081 |
0.200 |
0.142 |
0.066 |
0.083 |
0.041 |
0.095 |
12.82 |
||||||
| Saimiri boliviensis | 0.006 |
0.038 |
- |
0.012 |
0.071 |
0.032 |
0.053 |
0.062 |
0.134 |
0.036 |
4.44 |
||||||
| Aotus sp. | - |
- |
- |
0.012 |
- |
0.006 |
- |
0.090 |
0.010 |
- |
1.18 |
||||||
| Callicebus brunneus | 0.019 |
- |
0.047 |
0.023 |
- |
- |
0.007 |
- |
0.010 |
0.119 |
2.25 |
||||||
| Alouatta seniculus | 0.006 |
- |
0.070 |
0.010 |
0.026 |
0.065 |
0.020 |
0.007 |
0.082 |
0.024 |
3.10 |
||||||
| Cebus albifrons | - |
- |
- |
- |
0.006 |
0.026 |
- |
- |
- |
- |
0.32 |
||||||
| Cebus apella | - |
0.031 |
0.047 |
0.116 |
0.071 |
0.090 |
0.007 |
0.041 |
0.165 |
0.131 |
6.99 |
||||||
| Ateles paniscus | - |
- |
- |
- |
- |
- |
- |
- |
0.062 |
0.155 |
2.17 |
||||||
| Carnivora | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Atelocynus microtis | - |
- |
- |
- |
- |
- |
- |
- |
0.010 |
- |
0.10 |
||||||
| Nasua nasua | 0.006 |
- |
- |
0.012 |
0.013 |
0.019 |
- |
0.028 |
0.010 |
- |
0.88 |
||||||
| Eira barbara | 0.019 |
0.006 |
0.023 |
0.023 |
0.013 |
0.019 |
0.007 |
0.055 |
0.041 |
0.024 |
2.30 |
||||||
| Gallictis vitata | - |
- |
- |
- |
0.012 |
- |
- |
- |
- |
- |
0.12 |
||||||
| Lutra longicauda | - |
- |
- |
- |
- |
- |
- |
- |
- |
0.012 |
0.12 |
||||||
| Leopardus pardalis | 0.006 |
- |
- |
- |
- |
- |
0.013 |
- |
0.010 |
- |
0.29 |
||||||
| Perissodactyla | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Tapirus terrestris | - |
- |
- |
- |
- |
0.013 |
- |
- |
0.010 |
0.024 |
0.47 |
||||||
| Artiodactyla | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Tayassu pecari | - |
- |
- |
- |
- |
- |
- |
0.007 |
0.010 |
- |
0.17 |
||||||
| Tayassu tajacu | 0.019 |
0.044 |
0.093 |
0.081 |
- |
0.026 |
0.026 |
0.062 |
0.041 |
0.012 |
4.04 |
||||||
| Mazama americana | - |
0.019 |
- |
0.035 |
- |
0.032 |
0.013 |
0.034 |
0.010 |
0.012 |
1.55 |
||||||
| Rodentia | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Sciurus ignitus | 0.090 |
0.107 |
0.070 |
0.035 |
0.026 |
0.013 |
0.086 |
0.062 |
0.165 |
0.095 |
7.63 |
||||||
| Sciurus spadiceus | 0.705 |
0.547 |
0.407 |
0.209 |
0.510 |
0.374 |
0.238 |
0.303 |
0.196 |
0.357 |
38.46 |
||||||
| Dasyprocta variegata | 0.276 |
0.126 |
0.093 |
0.140 |
0.039 |
0.129 |
0.179 |
0.179 |
0.041 |
0.036 |
12.38 |
||||||
| Myoprocta pratti | - |
0.006 |
- |
0.012 |
0.006 |
- |
- |
- |
- |
- |
0.24 |
||||||
| Total | 1.462 |
1.107 |
1.000 |
0.814 |
0.994 |
0.987 |
0.742 |
1.014 |
1.082 |
1.131 |
|||||||
| Promedio | 1.285 |
0.907 |
0.991 |
0.878 |
1.107 |
1.033 |
|||||||||||
Tabla 3.10. IKA en términos de individuos/km de 26 especies.
| Albergues | CAPH |
EI |
ECO |
SACHA |
TRC |
Prom (x10-2) |
|||||||||||
| Tipo de Transecto | T |
C |
T |
C |
T |
C |
T |
C |
T |
C |
. | ||||||
| Km Censados | 156 |
159 |
86 |
86 |
155 |
155 |
151 |
145 |
97 |
84 |
. | ||||||
| Xenarthra | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Myrmecophaga tridactyla | - |
0.012 |
- |
- |
- |
- |
- |
- |
- |
- |
0.12 |
||||||
| Tamandua tetradactyla | - |
- |
0.013 |
- |
0.028 |
0.028 |
0.029 |
- |
0.011 |
- |
1.09 |
||||||
| Bradypus variegata | - |
- |
- |
0.012 |
- |
- |
- |
- |
0.021 |
- |
0.33 |
||||||
| Dasypus sp. | - |
- |
- |
0.012 |
- |
- |
- |
- |
- |
- |
0.12 |
||||||
| Primates | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Saguinus fuscicollis | 1.838 |
0.959 |
0.663 |
0.464 |
0.960 |
0.681 |
0.285 |
0.356 |
0.190 |
0.438 |
68.34 |
||||||
| Saimiri boliviensis | 0.213 |
1.257 |
- |
0.430 |
1.476 |
0.671 |
1.648 |
1.930 |
3.659 |
0.975 |
122.59 |
||||||
| Aotus sp. | - |
- |
- |
0.023 |
- |
0.026 |
- |
0.161 |
0.041 |
- |
2.51 |
||||||
| Callicebus brunneus | 0.044 |
- |
0.140 |
0.070 |
- |
- |
0.023 |
- |
0.025 |
0.286 |
5.88 |
||||||
| Alouatta seniculus | 0.026 |
- |
0.342 |
0.080 |
0.106 |
0.265 |
0.036 |
0.012 |
0.305 |
0.088 |
12.6 |
||||||
| Cebus albifrons | - |
- |
- |
- |
0.081 |
0.323 |
- |
- |
- |
- |
4.04 |
||||||
| Cebus apella | - |
0.110 |
0.377 |
0.942 |
0.305 |
0.388 |
0.023 |
0.141 |
0.940 |
0.746 |
39.72 |
||||||
| Ateles paniscus | - |
- |
- |
- |
- |
- |
- |
- |
0.322 |
0.805 |
11.27 |
||||||
| Carnivora | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Atelocynus microtis | - |
- |
- |
- |
- |
- |
- |
- |
0.010 |
- |
0.1 |
||||||
| Nasua nasua | 0.013 |
- |
- |
0.012 |
0.090 |
0.135 |
- |
0.221 |
0.134 |
- |
6.05 |
||||||
| Eira barbara | 0.019 |
0.006 |
0.030 |
0.030 |
0.015 |
0.023 |
0.008 |
0.066 |
0.082 |
0.048 |
3.27 |
||||||
| Gallictis vitata | - |
- |
- |
- |
0.012 |
- |
- |
- |
- |
- |
0.12 |
||||||
| Lutra longicauda | - |
- |
- |
- |
- |
- |
- |
- |
- |
0.012 |
0.12 |
||||||
| Leopardus pardalis | 0.006 |
- |
- |
- |
- |
- |
0.013 |
- |
0.010 |
- |
0.29 |
||||||
| Perissodactyla | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Tapirus terrestris | - |
- |
- |
- |
- |
0.013 |
- |
- |
0.010 |
0.024 |
0.47 |
||||||
| Artiodactyla | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Tayassu pecari | - |
- |
- |
- |
- |
- |
- |
0.207 |
0.515 |
- |
7.22 |
||||||
| Tayassu tajacu | 0.035 |
0.079 |
0.158 |
0.138 |
- |
0.077 |
0.061 |
0.143 |
0.078 |
0.023 |
7.92 |
||||||
| Mazama americana | - |
0.019 |
- |
0.035 |
- |
0.032 |
0.013 |
0.034 |
0.010 |
0.012 |
1.55 |
||||||
| Rodentia | . | . | . | . | . | . | . | . | . | . | . | ||||||
| Sciurus ignitus | 0.099 |
0.118 |
0.077 |
0.038 |
0.028 |
0.014 |
0.095 |
0.068 |
0.181 |
0.105 |
8.23 |
||||||
| Sciurus spadiceus | 0.846 |
0.657 |
0.488 |
0.251 |
0.612 |
0.449 |
0.286 |
0.364 |
0.235 |
0.429 |
46.17 |
||||||
| Dasyprocta variegata | 0.303 |
0.138 |
0.102 |
0.153 |
0.043 |
0.142 |
0.197 |
0.197 |
0.045 |
0.039 |
13.59 |
||||||
| Myoprocta pratti | - |
0.006 |
- |
0.012 |
0.006 |
- |
- |
- |
- |
- |
0.24 |
||||||
| Total | 3.442 |
3.361 |
2.401 |
2.610 |
3.757 |
3.268 |
2.716 |
3.901 |
6.827 |
4.029 |
|||||||
| Promedio | 3.402 |
2.506 |
3.513 |
3.309 |
5.428 |
3.631 |
|||||||||||
A continuación presentamos un resúmen de los resultados acerca de: ancho de grupo (G)
para las 10 especies más comunes; distancias de detección perpendicular de 12 especies
donde se encontró suficiente información para calcular límites de confiaza; el promedio
de las distancias de detección perpendicular de 14 especies;y finalmente estimaciones de
densidad para 12 especies.
Ancho de Grupo (G)
El ancho del grupo se identificó como la distancia-linial entre el primer individuo
avistado y el individuo, del mismo grupo, más alejado. Suficientes datos se obtuvieron
para determinar el ancho de grupo de 12 especies con una precisión aceptable (Tabla
3.11). Como se esperaba, el Fraile (S. boliviensis), que forma los grupos más grandes,
también tuvo el ancho de grupo más grande; con un promedio de 31 m. Como contraste el
Añuje (D. variegata) formó grupos pequeños de hasta 3 individuos, aunque normalmente
eran solitarios, con un ancho promedio de sólo 4 m. El ancho de grupo es generalmente
considerado una característica intrínsica de una especie y esta bajo la influencia de
variables tal como tamaño de grupo, tamaño corporal, y comportamiento.
Tabla 3.11. Promedio de ancho de grupo (G) de 10 especies. * Los datos que corresponden a
½ ancho de grupo son aquellos ajustados para incluir datos de encuentros con individuos
solitarios y es el valor utilizado en la ecuación 3.1.
| Especie | G (m) [2+ inds./gr.] |
½ G (m)* |
||
| . | N |
Promedio |
. | |
| Pichico | S. fuscicollis | 146 |
13 |
6.1 |
| Fraile | S. boliviensis | 11 |
31 |
14.6 |
| Tocón | C. brunneus | 13 |
11 |
4.7 |
| Coto | A. seniculus | 24 |
8 |
3.6 |
| Machin Negro | C. apella | 38 |
18 |
7.9 |
| Maquisapa | A. paniscus | 11 |
30 |
13.8 |
| Achuñi | N. nasua | 5 |
10 |
3.9 |
| Sajino | T. tajacu | 11 |
5 |
1.1 |
| Ardilla Roja | S. spadiceus | 28 |
6 |
0.2 |
| Añuje | D. variegata | 14 |
4 |
0.1 |
Distancia de Detección Perpendicular Específica
Cada especie de mamífero difiere en cuanto a su nivel de detectabilidad para un
observador humano, particularmente en hábitats densos como el bosque lluvioso donde la
visibilidad es reducida. Las variables importantes relacionadas con esta son: grado de
sociabilidad (las especies solitarias son generalmente más difíciles de detectar),
tamaño de grupo (los grupos grandes son más fáciles de detectar), peso promedio de la
especie (especies más grandes generalmente hacen más ruido al caminar, saltar o correr
incrementando así su detectabilidad), comportamiento antidepredador (algunas especies
tienden a correr al ver un depredador potencial (por ejemplo: un humano) mientras otras se
mantienen quietas) y el comportamiento de llamadas inter- e intra-grupal (las especies
más bulliciosas son más fáciles de detectar). Las innumerables permutaciones de estas
variables dotan a cada especie de una distancia de detección distinta.
Las distancias de detección perpendicular para las especies en Tabla 3.11 fueron
calculadas utilizando el Programa Distance (Laake et al. 1993). Este programa determina
una funcción (línia) que representa mejor los datos crudos de P1 de una especie. A la
vez el programa calcula la distancia perpendicular más alla del cual el número de
encuentros con grupos es igual al número de grupos que probablemente se pasaron por alto.
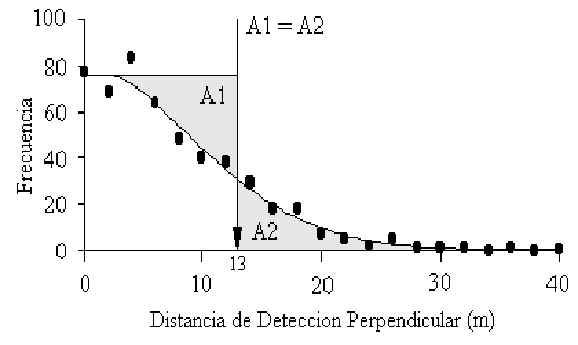
Figura 3.15. Distribución de la frecuencia de distancias de detección perpendiculares de
la Ardilla Roja (S. spadiceus). El Programa Distance calcula la funcción de detección
apropriada y es representada visualmente donde el área de A1 es gual al de A2; en este
caso 13m.
Tabla 3.12. Distancia de detección perpendicular específica (Psp), 95% IC, y modelos de
las funcciones de detección de 12 especies donde hubo suficientes datos. Modelo de
Funcciones: HNC = Half-Normal/Cosine; UC = Uniform/Cosine; HC = Hazard/Cosine.
| Especies | W (m) |
95% IC |
Modelo de Funcción |
. | Especies | W (m) |
95% IC |
Modelo de Funcción |
| T. tetradactyla | 22 |
12 – 38 |
UC |
. | N. nasua | 27 |
20 – 36 |
UC |
| S. fuscicollis | 32 |
28 – 35 |
HC |
. | T. tajacu | 15 |
13 – 18 |
UC |
| S. boliviensis | 50 |
40 –62 |
UC |
. | M. americana | 15 |
11 – 21 |
UC |
| C. brunneus | 26 |
18 – 39 |
HC |
. | S. ignitus | 10 |
9 – 12 |
UC |
| A. seniculus | 30 |
24 – 39 |
HNC |
. | S. spadiceus | 13 |
12 – 14 |
HNC |
| C. apella | 39 |
33 – 47 |
UC |
. | D. variegata | 13 |
11 – 15 |
HC |
Tabla 3.13. Promedio de la distancia de detección perpendicular (W), para 14 especies
donde no hubo suficientes datos para la determinación de límites de confianza 95%.
| Especies | W (m) |
. | Especies | W (m) |
| M. tridactyla | 15 |
. | E. barbara | 18 |
| B. variegata | 1 |
. | G. vitata | 10 |
| Dasypus sp. | 5 |
. | L. longicauda | 24 |
| Aotus sp. | 13 |
. | L. pardalis | 46 |
| C. albifrons | 77 |
. | T. terrestris | 50 |
| A. paniscus | 63 |
. | T. pecari | 50 |
| A. microtis | 18 |
. | M. pratti | 13 |
Tabla 3.14. Estimaciones de densidad de 12 especies (individuos/km2).
| Especies | CAPH |
EI |
ECO |
SACHA |
TRC |
|||||
| . | T |
C |
T |
C |
T |
C |
T |
C |
T |
C |
| T. tetradactyla | 0.0 |
0.0 |
0.5 |
0.0 |
1.1 |
1.1 |
1.2 |
0.0 |
0.5 |
0.0 |
| S. fuscicollis | 29.1 |
15.2 |
10.5 |
7.4 |
15.2 |
10.8 |
4.5 |
5.6 |
3.0 |
6.9 |
| S. boliviensis | 2.1 |
12.6 |
0.0 |
4.3 |
14.8 |
6.7 |
16.5 |
19.3 |
36.6 |
9.8 |
| C. brunneus | 0.8 |
0.0 |
2.6 |
1.3 |
0.0 |
0.0 |
0.4 |
0.0 |
0.5 |
5.4 |
| A. seniculus | 0.4 |
0.0 |
5.6 |
0.0 |
1.7 |
4.3 |
0.6 |
0.2 |
5.0 |
1.5 |
| C. apella | 0.0 |
1.4 |
4.8 |
12.0 |
3.9 |
4.9 |
0.3 |
1.8 |
11.9 |
9.5 |
| N. nasua | 0.2 |
0.0 |
0.0 |
0.2 |
1.7 |
2.5 |
0.0 |
4.1 |
2.5 |
0.0 |
| T. tajacu | 1.1 |
2.6 |
5.2 |
4.5 |
0.0 |
2.5 |
2.0 |
4.7 |
2.6 |
0.7 |
| M. americana | 0.0 |
0.6 |
0.0 |
1.2 |
0.0 |
1.1 |
0.4 |
1.1 |
0.3 |
0.4 |
| S. ignitus | 4.8 |
5.8 |
3.8 |
1.9 |
1.4 |
0.7 |
4.7 |
3.4 |
8.9 |
5.1 |
| S. spadiceus | 31.9 |
24.7 |
18.4 |
9.5 |
23.0 |
16.9 |
10.8 |
13.7 |
8.9 |
16.2 |
| D. variegata | 11.8 |
5.4 |
4.0 |
6.0 |
1.7 |
5.5 |
7.6 |
7.7 |
1.8 |
1.5 |
Hollos Forrageros
Se muestrearon un total de 342 quadrats durante el periodo del estudio, por un promedio de
3.1 muestras repetidas de cada quadrat, que equivale a un área total de muestreo de 13.68
hectáres (Tabla 3.15). Este esfuerzo logró encontrar un total de 751 hollos forrageros.
La distribución de hollos por quadrat era significativamente agrupada y caracterizada por
una distribución binomial negativa (Figura 3.16). Una prueba de Wilcoxon reveló que la
densidad de hollos a lo largo de los T en todos los albergues era significativamente menor
que los C (p = 0.03), un patrón que es evidente en la Figura 3.17. Además, una
correlación de Pearson reveló una relación negativa y significativa entre la intensidad
del tránsito y la densidad de hollos a lo largo de los T (Pearson = -0.92, p < 0.05).
Tabla 3.15. Resúmen de los datos sobre hollos forrageros. Area de cada cuadrat = 0.04 ha.
| Albergue |
. | No. de cuadrats |
No. total de muestras |
Área Muestreada (ha) |
No. de hollos |
Densidad Promedia (hollos/cuadrat) |
Var. |
Índice de Disperción |
| CAPH | T |
16 |
49 |
1.96 |
43 |
0.88 |
1.77 |
2.02 |
| . | C |
13 |
35 |
1.40 |
73 |
2.09 |
4.54 |
2.18 |
| EI | T |
7 |
23 |
0.92 |
24 |
1.04 |
2.50 |
2.40 |
| . | C |
7 |
24 |
0.96 |
156 |
6.50 |
40.58 |
6.24 |
| ECO | T |
11 |
40 |
1.60 |
36 |
0.90 |
2.86 |
3.18 |
| . | C |
13 |
52 |
2.08 |
119 |
2.29 |
3.03 |
1.32 |
| SACHA | T |
9 |
26 |
1.04 |
36 |
1.38 |
4.67 |
3.37 |
| . | C |
9 |
24 |
0.96 |
146 |
6.08 |
19.54 |
3.21 |
| TRC | T |
13 |
35 |
1.40 |
50 |
1.43 |
1.54 |
1.08 |
| . | C |
12 |
34 |
1.36 |
68 |
2.00 |
1.82 |
0.91 |
| Total | . | 110 |
342 |
13.68 |
751 |
2.20 |
9.73 |
2.59 |
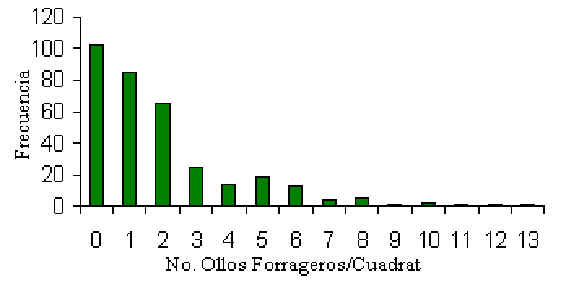
Figura 3.16. Distribución de la frecuencia de número de hollos forrageros por cuadrat.
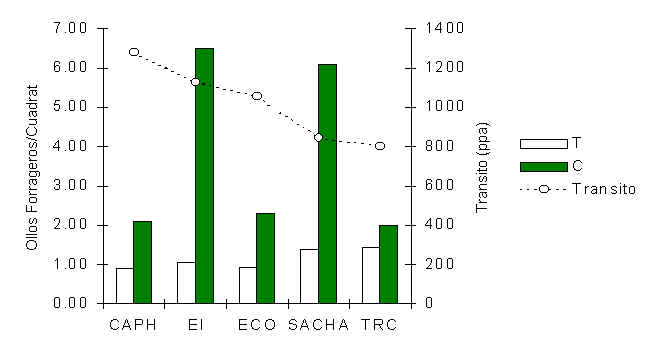
Figura 3.17. Densidad de hollos forrageros a lo largo de T y C en cada albergue.