El Impacto del Turismo sobre la Herpetofauna
en Tambopata, Sureste del Peru.
Tiffany M. Doan & Wilfredo Arizábal Arriaga
Introducción
Los mamíferos y aves de Sudamerica, han recibido una inmensa cantidad de atención; contrariamente los anfíbios y reptiles de la selva amazónica han quedado mayormente sin estudiar (Vitt 1996). El primer estudio exhaustivo de una comunidad de herpetofauna en Sudamérica fue una lista anotada de las especies de Santa Cecilia, Ecuador (Duellman 1978). Posteriormente, se estudiaron las comunidades de la herpetofauna de Iquitos, Perú (Dixon y Soini 1986, Duellman y Mendelson 1995) y Parque Nacional del Manú, Perú (Rodríguez y Cadle 1990, Morales y McDiarmid 1996), así que se comenzó el Proyecto de Tamaño Mínimo Critico de Ecosistemas cerca a Manaus, Brasil (Zimmerman y Rodrigues 1990). Para la región Tambopata del sureste de Perú, herpetólogos escribieron listas preliminares de cuatro lugares (McDiarmid y Cocroft, no publ., Duellman y Salas 1991, Salas 1995, Sehgelmeble 1997). Todos los estudios arriba mencionados fueron simplemente listas de especies; Duellman (1990, también Duellman y Koechlin 1991) resumió el conocimiento de comunidades de herpetofauna de la selva neotropical, encontrando que se necesita realizar más investigaciones sobre la ecología de las especies.
La unidad de herpetofauna de TReeS-RAMOS Proyecto Tambopata, es el primer estudio que análiza los impactos antropogénicos potenciales en las comunidades de herpetofauna de la selva. Aunque muchos estudios han demostrado que el turismo modifica comunidades de mamíferos y aves (Griffiths and Van Schaik 1993, Hindinger 1996, Yoon et al. 1997), no se conoce los efectos en la herpetofauna. Los anfíbios y reptiles, de los cuales muchos son nocturnos, arborícolas, y subterraneos, no tienen tanto contacto con turistas como los mamíferos y aves más grandes y visibles. Sin embargo, la presencia de turistas y sus trochas puede afectar la herpetofauna local.
Para el estudio de herpetofauna, escogimos con cuidado las ubicaciones de las parcelas para igualar los tipos de bosque (referirse a Phillips 1993) y la historia de la utilización de las trochas. Como cada albergue turístico tiene diferentes tipos de bosques disponibles, escogimos un tipo por albergue para censar. Preferíamos los tipos más antiguos y altos (Bosques de Terraza Vieja, Tipo F; y Terra Firme, Tipos G y H), para facilitar los censos. Como resultado de esta selección, varias especies presentes en los albergues no fueron registradas. Por ejemplo, hay muchos anfíbios que sólo se encuentran en las riveras de los lagos y ríos, sin embargo nuestras parcelas no incluyeron estos microhábitats. Este estudio no intentó registrar la totalidad de poblaciones de las especies; una cantidad menor es suficiente para mostrar patrones en áreas con turismo (tratamiento) y áreas sin turismo (control).
Los tipos de bosques que se utilizaron en la investigación de herpetofauna se muestran a continuación:
EcoAmazonía (ECO) - Bosque Estacionalmente Inundado (Tipo B)
Cusco Amazónico (CAPH) - Bosque Estacionalmente Inundado / Bosque de Terraza Vieja Transición (Tipo B/F).
Explorers’ Inn (EI) - Bosque de Terra Firme Arcilla-Arenosa (Tipo H)
Sachavacayoc (SACHA) - Bosque de Terraza Vieja / Bosque de Terra Firme Arcilloso Transición (Tipo F/G) con bambú
Tambopata Research Center (TRC) - Bosque de Terraza Vieja / Bosque de Terra Firme Arcilloso Transición (Tipo F/G) con bambú
Metodología
Cada uno de los cinco albergues fueron visitados cinco veces durante los dos años de estudio con la misma cantidad de días de muestreo durante cada visita. Para calcular los niveles de población de anfíbios y reptiles en áreas de turismo actual y áreas de control sin turismo, usamos dos métodos principales: "Quadrats" y "MEV" (Muestreo de Encuentro Visual).
Quadrats
El método de quadrats es una de las técnicas más efectivas para la herpetofauna (Jaeger and Inger 1994). Cada quadrat, que mide 8 x 8 metros, fue muestreado intensivamente por cuatro observadores. Cada observador empezó en una de las esquinas del quadrat. Siguiendo a la misma velocidad, cada una de las cuatro personas buscó un metro de ancho, para luego avanzar hasta la siguiente esquina y finalmente reunirse en el centro del quadrat. El movimiento sincronizado sirvió para impedir que los anfíbios y reptiles puedan escapar del quadrat antes de ser registrados. Los cuatro observadores consistieron de dos coordinadores (los autores) y dos asistentes. Con cuatro observadores, terminamos cada quadrat en un promedio de 11,33 minutos durante el día y 14,88 minutos en la noche.
Los observadores buscaron reptiles y anfíbios tanto en hojarasca como en troncos, ramas y hojas, hasta una altura de 2 metros. Cada individuo fue capturado directamente con la mano, medido hasta 0,05 mm con calipers, pesado en una romana de precisión de 0,25 g, marcado y devuelto al punto de captura. Además anotamos el substrato y altura de cada captura. Las ranas y lagartijas fueron marcados individualmente cortando los dedos de acuerdo al método de Twitty (1966, Donnelly et al. 1994). No se marcaron las culebras y caecilias por la muy baja probabilidad de recaptura.
Para cada quadrat de 8 x 8 m se anotó características ambientales y del hábitat, registrándose la temperatura ambiental al inicio del muestreo y el diámetro de todos los árboles mayores a los 10 cm (dbh). La hojarasca fue medida contando el número de hojas atravésadas por un punzón. En cada lado del quadrat se tomó cuatro registros para finalmente tener 16 estimaciones de la cantidad de altura de hojarasca. Tomamos las variables ambientales para explicar la variación entre quadrats.
Los quadrats se instalaron en parcelas de 80 x 70 m. Cada uno de los 56 quadrats por parcela fue ubicado dentro de un cuadrado de 10 x 10 m dejando 2 m entre quadrats contiguos y un metro entre los quadrats y el borde de la parcela. En cada albergue fueron instaladas dos parcelas de quadrats sobre las trochas en uso por turistas y otras dos parcelas de control fueron instaladas por lo menos 200 m alejadas de estas y de las parcelas de tratamiento en general. La distancia de 200 m se determinó después de concluir que la mayoría de los anfíbios y reptiles registrados no tienen un rango mayor de 100 m.
De los 56 quadrats en cada parcela, 28 fueron muestreados por visita. Los quadrats muestreados se eligieron al azar, utilizando una tabla de números aleatorios. Durante las dos primeras visitas a un albergue trabajamos 14 de los 28 quadrats elegidos al azar en la mañana y 14 en la noche. Empezando con la tercera visita, trabajamos 22 de los quadrats en la noche y el resto (6) en la mañana. La razón por el cambio es que la taza de captura para casí todas las especies es mayor durante la noche, entonces el cambio aumentó la taza de captura. En total, 112 quadrats fueron muestreados en cada visita por albergue.
Muestreo por Encuentro Visual (MEV)
Se instalaron dos parcelas, de 1 hectárea cada una, ubicando el tratamiento en una trocha usada por turistas y otra de control. Veinticuatro transectos lineales de 100 m fueron marcados por parcela, con 4 m de separación entre transectos contiguos. Por cada transecto se habilitó un sendero para facilitar el muestreo y asegurar que los observadores no salgan del transecto cuando se orientan con brújulas durante las evaluaciones nocturnas. Cada transecto fue muestreado dos veces por visita, una de día y otra de noche. Al tercer día de instalación de los transectos se inició la evaluación, dejando por lo menos dos días entre las visitas a los mismos transectos.
El tiempo empleado en cada uno de los transectos duró en promedio de 25,71 minutos para transectos diurnos y 39,63 minutos para transectos nocturnos. Este tiempo lento sirvió para que los observadores pudieran buscar sobre toda la vegetación a su alrededor. La búsqueda de anfíbios y reptiles en transectos se realizaron visualmente sin la necesidad de perturbar la vegetación como ocurre en el caso de los quadrats. El equipo de observadores consistió de un coordinador, quien dirigió y determinó el lapso de tiempo, acompañado de dos asistentes quienes procesaron todos los ejemplares. Los individuos registrados en los transectos fueron procesados en forma similar a los quadrats, con la diferencia que fueron anotados en el número de transecto, la distancia a lo largo del transecto y la hora de cada captura. También el diámetro de los árboles no fue anotado en las parcelas de MEV.
Tanto los quadrats y MEV fueron útilizados en este estudio porque con cada método se registran diferentes especies (Crump and Scott 1994). Las lagartijas grandes, culebras y ranas arborícolas fueron registradas mayormente con MEV, mientras que los anfíbios y lagartijas pequeñas que viven en la hojarasca fueron capturados en quadrats.
Análisis
Cada albergue fue muestreado cinco veces durante el estudio de dos años. Se calculó el total de horas-persona por resumir la cantidad total de quadrats y transectos terminados durante el período de dos años (3,336 quadrats; 2,832 transectos) y por multiplicar esos números por la duración promedio del método (diurno y nocturno censos separados) y el número de investigadores presentes.
Se hicieron cuatro categorías de análisis. Los tres primeros son análisis individuales para cada albergue. La última categoría es una comparación entre todos los alberguees de Tambopata. El primer tipo de análisis comparó las tres parcelas de tratamiento (dos quadrats, un transecto) contra las parcelas de control para determinar si existe algún efecto directo en la abundancia, riqueza de especies o diversidad de las comunidades de herpetofauna. El segundo tipo probó efectos en las especies individualmente. El tercero examinó los efectos de las trochas de turistas en la comunidad de herpetofauna. El cuarto tipo de análisis examinó la comunidad de herpetofauna completa, considerando los factores abióticos. Consideramos cada tipo de análisis más profundamente a continuación.
Tratamiento vs. Control
Se calculó la diversidad de herpetofauna con el Índice de Shannon-Wiener (H’) para combinar entre las tres parcelas de tratamiento y las tres parcelas de control. Se calculó la semejanza de comunidad por los índices de Morisita y Porcentaje de Semejanza. Usamos los dos índices porque producen resultados ligeramente diferentes; Krebs (1999) recomendó Porcentaje de Semejanza mientras Wolda (1981) recomendó el Índice de Morisita como el mejor índice disponible. Se probó las abundancias de tratamiento versus control, por especie de anuro y lagartija, usando la Prueba de Wilcoxon. Las abundancias de culebras no eran suficientemente grandes para usar en este análisis.
Abundancia por Especie en Parcelas
Considerando las seis parcelas individualmente, probamos las abundancias de especies de los tratamientos versus control. No probamos todas las especies encontrados en Tambopata porque varias especies no son suficientemente abundantes por parcela para producir resultados válidos. Entonces, escogimos las ocho especies de anuros más abundantes de la región para el análisis. Algunas especies no están presentes (o están presentes parcialmente) en todos los albergues; no análizamos estas especies. Además, pusimos en orden las abundancias de especies de anuros por albergue y utilizamos todas las especies de las diez mayores de cada albergue (que no eran las ocho primeras). De lagartijas, probamos Anolis fuscoauratus, presente en todos los albergues; otras tres especies de lagartijas: Gonatodes humeralis, Pseudogonatodes guianensis y Priónodactylus eigenmanni son abundantes en dos albergues del río Madre de Dios (ECO y CAPH). Comparamos estadísticamente las especies de este análisis con la prueba de Mann-Whitney U. No análizamos las culebras.
Efectos de Trocha
Para determinar si la trocha de turistas afecta la abundancia, riqueza de especies o diversidad de anfíbios y reptiles, los quadrats en contacto con la trocha (quadrats de trocha) fueron comparados a una cantidad igual de quadrats elegidos al azar que no tocaron la trocha. Las pruebas de Mann-Whitney U compararon la riqueza de especies, diversidad y abundancia de especies comunes. Utilizamos los índices de Porcentaje de Semejanza y Morisita para comparar entre las comunidades de anuros y lagartijas de la trocha usada por turistas y "no trocha". Se omitieron las culebras de todos los análices de trocha por sus poblaciones bajas.
Para adquirir una perspectiva más fina de los efectos potenciales de la trocha, agrupamos los quadrats dentro de cada parcela en unidades basadas en la distancia de la trocha al quadrat. Así se establecieron cinco grupos: quadrats empezando no más cerca de cinco metros de la trocha y no más lejos de diez metros de la trocha, quadrats empezando 11-15 m de la trocha, quadrats empezando 16-20 m de la trocha y quadrats más de 20 m de la trocha. Se excluyeron los quadrats empezando menos de cinco metros de la trocha. Determinamos la cantidad de quadrats en cada grupo por el número de quadrats disponibles en todos los grupos, dependiendo en la ubicación de la trocha y la parcela. Algunas parcelas no tuvieron cantidades suficientes de quadrats por grupo (por ejemplo parcelas donde una trocha sinuosa llenó casí toda la parcela), entonces, excluimos esas parcelas. Comparamos la diversidad de cada grupo con la distancia de la trocha en metros usando una regresión lineal para determinar si la distancia de la trocha causa directamente los niveles de diversidad en anuros. Adicionalmente, probamos la diversidad y abundancia de especies comunes por grupo por albergue con la prueba de Kruskal-Wallis.
Efectos Generales en Tambopata
El último tipo de análisis examinó los patrones potenciales de la región entera de Tambopata, reuniendo los resultados de los cinco alberguees a la vez. Primero, todos los quadrats por cada grupo de trocha (referirse a 3. Efectos de Trocha, arriba) por albergue fueron combinados y la diversidad de la región fue determinado con el Índice de Shannon-Wiener. Usando los cinco albergues, la prueba de Kruskal-Wallis determinó si la distancia de la trocha turística afectó la diversidad de anuros como patrón general en Tambopata.
También, comparamos la diversidad de tratamiento (dos parcelas de quadrats más una parcela de transectos) de cada albergue (referirse a Tabla 2) versus la cantidad anual de turistas de cada albergue con una regresión (usamos la escala logarítmica de cantidad de turistas para controlar variabilidad). Utilizamos esta regresión para determinar si la cantidad de turistas (quienes caminan las trochas turísticas de este estudio todos los días) afecta la diversidad de anuros o lagartijas en Tambopata.
Resultados
El intensivo estudio de herpetofauna durante los dos años del Proyecto Tambopata rindió 6,012.92 horas-personas, dividido entre censos diurnos y nocturnos y metodologías de quadrats y MEV. El número de individuos capturados en todo el estudio fue 19,957, con 18,146 anfíbios y 1,811 reptiles. Cuando se refiere a la riqueza de especies, capturamos 88 especies de anfíbios y 55 especies de reptiles durante los censos. Más específicamente, eran 87 especies de anuros, una caecilia, una especie de tortuga, 23 especies de lagartijas y 32 especies de culebras. Registramos más especies durante horas fuera del estudio pero no presentamos esos datos aquí. La Figura 4.1 es una representación gráfica del número de especies nuevas registradas por mes del estudio. La Figura 4.2 representa un gráfico de la acumulación de esas especies por mes. Se puede ver en esta figura que la curva de anfíbios empieza a suspender la incrementación, haciéndose constante, después del mes diecisiete, significando que el porcentaje mayor de todas las especies fuer registrado en el estudio. La porción de las especies que no fueron registradas hasta ese mes representa una porción no significante de las especies que actualmente viven en Tambopata. Para el caso de la curva de los reptiles, que empieza a suspender la incrementación en el mes catorce, significa que la mayoría de las especies que se puede capturar con las metodologías de quadrats y transectos ya fueron capturados antes de este mes. En la actualidad, estudios anteriores realizados por otros autores (por ejemplo McDiarmid y Cocroft, no publ., Duellman y Salas 1991) registraron más especies al censar una diversidad más amplia de tipos de bosques que el presente estudio. Las otras especies consisten mayormente de culebras (gran parte de las cuales son arborícolas y acuáticas).
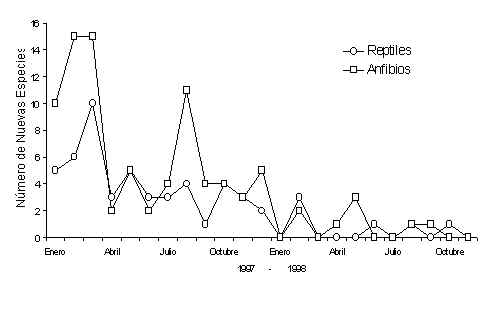
Figura 4.1. Número de nuevas especies por mes.
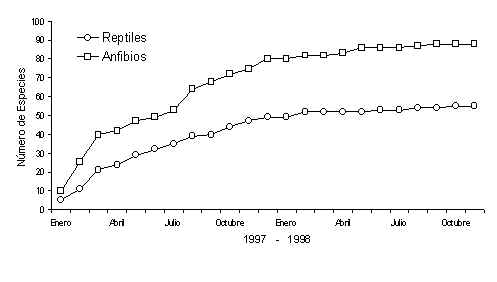
Figura 4.2. Acumulación de especies.
Tratamiento vs. Control
Se presenta en la Tabla 4.1 la riqueza de especies de herpetofauna en las áreas de tratamiento y control en cada albergue. Se muestra el Índice de Diversidad de Shannon-Wiener para cada albergue en la Tabla 2. TRC tiene la mayor diversidad de anfíbios; EI presenta la menor diversidad. Aunque no existe una prueba significativa para Shannon-Wiener, es obvio que CAPH tiene una mayor diversidad de anfíbios dentro de sus áreas de control, mientras EI tiene más alta diversidad en sus áreas de tratamiento, los otros tres albergues tienen diversidades similares entre tratamiento y control. Para los reptiles, CAPH tiene la mayor diversidad mientras SACHA tiene la más baja diversidad. Tanto EI como SACHA tienen mucha mayor diversidad en las áreas de tratamiento que en las áreas de control, pero en los otros tres albergues las diversidades son similares en ambas áreas.
Tabla 4.1. Riqueza de especies capturadas durante muestreos en los cinco albergues.
| Albergue | Tratamiento |
Control |
||||
| . | Ranas |
Lagartijas |
Culebras |
Ranas |
Lagartijas |
Culebras |
| CAPH | 32 |
12 |
11 |
36 |
12 |
12 |
| EI | 39 |
4 |
11 |
27 |
5 |
6 |
| ECO | 42 |
14 |
14 |
41 |
14 |
12 |
| SACHA | 38 |
10 |
15 |
36 |
7 |
9 |
| TRC | 43 |
10 |
9 |
44 |
8 |
13 |
Tabla 4.2. Diversidad de Anfíbios y Reptiles en Tambopata, Índice de Shannon-Wiener, Tratamiento vs. Control.
| Albergue | Tratamiento |
Control |
||
| . | Anfíbios |
Reptiles |
Anfíbios |
Reptiles |
| CAPH | 0.833 |
1.001 |
1.076 |
1.005 |
| EI | 0.943 |
0.934 |
0.794 |
0.777 |
| ECO | 1.062 |
0.896 |
1.004 |
0.934 |
| SACHA | 0.980 |
0.863 |
0.960 |
0.564 |
| TRC | 1.104 |
0.986 |
1.156 |
0.911 |
Tabla 4.3. Semejanza de comunidad de herpetofauna de Tambopata; Tratamiento vs Control.
| Albergue | % Similitud |
Morisita |
||
| . | Anfíbios |
Reptiles |
Anfíbios |
Reptiles |
| CAPH | 72.883 |
75.060 |
0.867 |
0.958 |
| EI | 74.236 |
62.973 |
0.921 |
0.951 |
| ECO | 80.545 |
84.680 |
0.919 |
0.982 |
| SACHA | 86.616 |
73.622 |
0.971 |
0.967 |
| TRC | 62.793 |
68.109 |
0.742 |
0.983 |
Tabla 4.4. Abundancia relativa por especie probabilidades de Wilcoxon, * P<0.05, ** P<0.01, *** P<0.001, T = tratamiento, C = control.
| Albergue | Anuros |
Reptiles |
| CAPH | 0.896 |
0.030 *C |
| EI | 0.000 ***T |
0.138 |
| ECO | 0.899 |
0.334 |
| SACHA | 0.153 |
0.001 ***T |
| TRC | 0.628 |
0.861 |
Para una perspectiva más amplia de los resultados del estudio turístico de la herpetofauna en total, usamos índices de semejanza. Se muestran los resultados de los Índices de Porcentaje de Semejanza y Morisita en la Tabla 4.3. Según los dos índices, TRC tiene la diferencia más grande de anuros entre tratamiento y control (significando que el turismo ha afectado la comunidad de anfíbios y reptiles más que los otros alberguees) mientras SACHA tiene el efecto menor. Para reptiles, los resultados son muy diferentes, ECO con un efecto de turismo menor, mientras EI mostró el efecto mayor.
Cuando comparamos las abundancias relativas de todas las especies presentes por albergue en tratamiento versus control, la prueba de Wilcoxon mostró que sólo EI (Z = -4.133, p = 0.000) tuvo abundancias de anuros significativamente diferentes en la totalidad de las especies con abundancias mayores en el área de tratamiento (referirse a Tabla 4). SACHA tuvo abundancias relativas mayores de reptiles en el área de tratamiento, pero CAPH tuvo abundancias relativas mayores en las áreas de control.
Abundancia de Parcela por Especie Individual
Se presenta los resultados de la prueba de Mann-Whitney de abundancia de parcela por especie individual en la Tabla 4.5. Los números en la tabla son los niveles calculados de probabilidad (p). Ninguna de las cuatro especies de lagartijas produjo resultados significativos; en cambio algunas especies de anuros tuvieron abundancias mayores en cualquiera de las áreas de tratamiento o control.
Ninguno de los dos albergues en el río Madre de Dios, ECO ni CAPH, tuvo diferencias significativas, mientras que cada uno de los albergues en el río Tambopata tuvieron por lo menos dos resultados significativos. EI tuvo abundancias mayores del dendrobatido Colostethus marchesianus, el bufonido Dendrophryniscus minutus y el leptodactylido Physalaemus petersi en las parcelas de tratamiento. SACHA registró abundancias mayores del leptodactylido Phyllonastes myrmecoides y el microhylido Ctenophryne geayi en las parcelas de tratamiento. TRC tuvo abundancias mayores del bufonido Bufo cf. typhonius, el microhylido Chiasmocleis bassleri y Physalaemus petersi en las parcelas de tratamiento, mientras el dendrobatido Epipedobates pictus estuvo más abundante en el área de control. Para todas las otras especies en esta prueba, se aceptó la hipótesis nula de ninguna relación entre las parcelas de turismo y sin turismo.
Efectos de Trocha
Cuando consideramos los efectos potenciales de la trocha turística, no encontramos un efecto significativo de la riqueza de especies de anuros en ningún albergue (Tabla 4.6). En cambio, la diversidad de anuros fue distinto significativamente en SACHA (U = -60.00, p = 0.029) por tener mayor diversidad en los quadrats de la trocha, pero en ningún otro albergue (referirse a Tabla 4.6). Especie por especie, se encontró sólo el microhylido Hamptophyrne boliviana más en la trocha turística que fuera de la trocha (referirse a Tabla 4.7). En todos los casos de riqueza de especies, diversidad y por especie, las abundancias de lagartijas y culebras fueron demasiadamente bajas para descubrir algún patrón. Los índices de semejanza de anuros entre trocha y grupos fuera de la trocha mostraron alta semejanza, siendo CAPH el mayor y TRC el menor (Tabla 4.8). Las comunidades de reptiles tienen semejanza baja, probablemente porque hay muchas especies con pocos individuos por especie.
En la escala más fina, la regresión de diversidad de grupos de trocha en distancia de los quadrats de la trocha turística no produjo resultados significativos (Tabla 4.9). Cuando consideramos distancia de la trocha turística, no descubrimos patrones en abundancia de especies individuales (referirse a Tabla 4.10).
Tabla 4.5. Resultado de abundancia de parcela, probabilidades de Mann-Whitney. Al = Abundancia insuficiente para probar; * p<0.05, ** p<0.01, *** p<0.001, T = Tratamiento, C = Control.
| . | . | Albergues |
||||
| . | Los ocho anuros más abundantes | CAPH |
EI |
ECO |
SACHA |
TRC |
| 1 | Adenomera andreae | 0.275 |
0.184 |
0.513 |
0.513 |
0.127 |
| 2 | Hamptophryne boliviana | 0.275 |
0.121 |
0.127 |
0.827 |
0.825 |
| 3 | Eleutherodactylus toftae | 0.376 |
IA |
IA |
0.827 |
0.275 |
| 4 | Chiasmocleis ventrimaculata | 0.827 |
IA |
0.827 |
0.813 |
0.507 |
| 5 | Colostethus marchesianus | 0.513 |
0.050*T |
0.513 |
IA |
IA |
| 6 | Eleutherodactylus peruvianus | 0.261 |
0.827 |
0.827 |
0.261 |
0.827 |
| 7 | Phyllonastes myrmecoides | IA |
0.121 |
IA |
0.050*T |
0.513 |
| 8 | Bufo cf. Typhonius | 0.500 |
0.127 |
0.827 |
IA |
0.050*T |
| Otros anuros abundantes | ||||||
| 9 | Dendrophryniscus minutus | . | 0.050*T |
. | . | . |
| 10 | Epipedobates pictus | . | . | 0.184 |
. | 0.050*C |
| 11 | Hyla fasciata | . | . | . | . | 0.827 |
| 12 | Osteocephalus taurinus | . | 0.077 |
. | . | . |
| 13 | Scarthyla ostinodactyla | . | . | 0.513 |
. | . |
| 14 | Scinax cruentomma | . | . | . | 0.827 |
. |
| 15 | Scinax garbei | . | . | 0.507 |
. | . |
| 16 | Scinax icterica | . | . | . | 0.513 |
. |
| 17 | Scinax pedromedinai | 0.268 |
. | 0.513 |
. | . |
| 18 | Scinax rubra | . | . | . | 0.513 |
. |
| 19 | Eleutherodactylus skydmainos | . | . | . | . | 0.127 |
| 20 | Eleutherodactylus sp. 1 | . | 0.077 |
. | . | . |
| 21 | Leptodactylus mystaceus | 0.127 |
. | 0.827 |
0.268 |
. |
| 22 | Physalaemus petersi | . | 0.050*T |
. | . | 0.050*T |
| 23 | Chiasmocleis bassleri | . | . | . | . | 0.046*T |
| 24 | Ctenophryne geayi | 0.376 |
. | . | 0.050*T |
. |
| 25 | Elachistocleis bicolor | 0.127 |
. | . | . | . |
| . | Las cuatro especies de lagartijas más abundantes | . | . | . | . | . |
| 1 | Anolis fuscoauratus | 0.268 |
0.658 |
0.376 |
0.268 |
0.513 |
| 2 | Priónodactylus eigenmanni | 0.827 |
. | 0.827 |
. | . |
| 3 | Gonatodes humeralis | 0.127 |
. | 0.827 |
. | . |
| 4 | Pseudogonatodes guianensis | 0.513 |
. | 0.658 |
. | . |
Efectos Generales en Tambopata
Cuando utilizamos los grupos de trocha de todos los albergues para descubrir patrones en la totalidad de la región de Tambopata, no detectamos diferencias significativas (K-W = 1.935, p = 0.748). Las regresiones de diversidad de la parcela de tratamiento en las cantidades anuales de turistas también no fueron significativos. La regresión de anuros sugirió que un patrón ligero puede estar presente (F = 6.981, p = 0.078), mientras la diversidad de lagartijas no mostró ningún patrón (F = 0.344, p = 0.599).
Tabla 4.6. Resultados de trocha vs. no-trocha de anuros, probabilidades de Mann-Whitney.
| Albergue | Riqueza de Especies |
Diversidad de Especies |
| CAPH | 0.563 |
0.659 |
| EI | 0.920 |
0.582 |
| ECO | 0.126 |
0.286 |
| SACHA | 0.183 |
0.029*T |
| TRC | 0.196 |
0.157 |
Tabla 4.7. Resultados por especie de trocha vs no-trocha por albergue, probabilidades de Kruskall-Wallis.; * p < 0.05, ** p < 0.01, *** p < 0.001.
| Albergue | CAPH |
EI |
ECO |
SACHA |
TRC |
| Adenomera andreae | 0.600 |
0.885 |
. | 0.786 |
. |
| Eleutherodactylus toftae | . | . | . | 0.562 |
0.919 |
| Physalaemus petersi | . | 0.796 |
. | . | . |
| Chiasmocleis ventrimaculata | 0.697 |
. | . | 0.376 |
. |
| Hamptophryne boliviana | 0.713 |
. | 0.006**T |
0.727 |
. |
Tabla 4.8. Resultados de índices de semejanza entre comunidades, trocha vs no-trocha.
| Albergue | % Similitud |
Morisita |
||
| . | Anfíbios |
Reptiles |
Anfíbios |
Reptiles |
| CAPH | 87.668 |
70.370 |
1.008 |
1.074 |
| EI | 81.715 |
75.000 |
0.991 |
1.875 |
| ECO | 75.655 |
66.194 |
0.973 |
0.946 |
| SACHA | 80.967 |
40.000 |
0.953 |
0.908 |
| TRC | 69.423 |
IA |
0.911 |
IA |
Tabla 4.9. Resultados de regresión por albergue basado en grupos de trocha.
| Albergue | Probabilidad de Prueba F |
| CAPH | 0.324 |
| EI | 0.986 |
| ECO | 0.545 |
| SACHA | 0.512 |
| TRC | 0.199 |
Tabla 4.10. Resultados de grupos de trocha, probabilidades de Kruskall-Wallis * p < 0.05, ** p < 0.01, *** p < 0.001.
| Especies | CAPH |
EI |
ECO |
SACHA |
TRC |
| Dendrophryniscus minutus | . | 0.204 |
. | . | . |
| Scinax spp. | . | . | . | 0.970 |
. |
| Adenomera andreae | 0.746 |
0.850 |
. | 0.168 |
. |
| Eleutherodactylus peruvianus | . | 0.134 |
. | . | . |
| Eleutherodactylus toftae | . | . | . | 0.146 |
0.975 |
| Leptodactylus mystaceus | . | . | . | 0.970 |
. |
| Phyllonastes myrmecoides | . | . | . | 0.234 |
. |
| Physalaemus petersi | . | 0.417 |
. | . | . |
| Chiasmocleis ventrimaculata | 0.826 |
. | . | 0.223 |
. |
| Ctenophryne geayi | . | . | . | 0.975 |
. |
| Hamptophryne boliviana | 0.646 |
. | 0.212 |
. | . |
Discusión
La unidad de herpetofauna de TReeS-RAMOS Proyecto Tambopata censó una variedad espectacular de especies de anfíbios y reptiles. Nunca anteriormente investigadores han realizado un estudio de herpetofauna de esta escala o duración en el biotopo de selva tropical. En consecuencia, con casí 20.000 individuos registrados, este estudio únicamente puede distinguir patrones del efecto turístico en la herpetofauna local si, en la actualidad, existen tales patrones.
Las pruebas entre tratamiento y control nos proporcionan una variedad grande de perspectivas que producen una serie igualmente grande de resultados diversos. Existen unos patrones importantes que podemos recoger de estos resultados. EI tiene diversidad y abundancia relativa de anuros más alta en las áreas de tratamiento. Para los reptiles, EI y SACHA son más diversos en las áreas de tratamiento. En cambio, CAPH tiene diversidad y abundancia relativa más alta de anfíbios y reptiles en las áreas de control. Los otros alberguees no tienen relaciones claras entre turismo y la diversidad o abundancia relativa de herpetofauna.
En efecto, la presencia del turismo afecta a la comunidad de anuros. Es el caso más obvio en CAPH donde la diversidad y abundancia relativa de anfíbios son más altas en las áreas de control después de casí veinticinco años de la presión diaria de turistas. Cuando la vegetación de la trocha está pisoteada y luego muere, la ausencia de la cobertura de plantas puede forzar que se muden los artrópodos, la presa primaria de anuros. Entonces, los anuros han perdido su fuente de alimentación y tienen que vaciar la región de la trocha para buscar su presa de artrópodos. A tiempo, los anuros descubren otras fuentes de alimentación fuera del área de la trocha.
Los efectos en EI también apoyan la hipótesis alternativa que el turismo afecta a la comunidad de herpetofauna, pero en otro sentido. Parecido a CAPH, EI ha estado recibiendo turistas para casí veinticinco años. Pero en forma disimilar a CAPH, que tiene bosque bajo e inundado estacionalmente, el bosque Tipo H de las parcelas de EI nunca se inunda. La trochas de EI son muy erosiónadas y se vuelven en posos durante la época lluviosa, atrayendo muchas especies de ranas a la fuente limitada de agua. Los autores han observado en repetidas veces oviposición por varias especies de anuros en las trochas de EI (dentro de las parcelas de tratamiento). Los posos temporales aparentemente son albergues de oviposición conveniente, pero cuando cuarenta turistas pisotean el área al día siguiente, la taza de supervivencia de los huevos y renacuajos es probablemente mínimo. En EI los anuros no pueden migrar a áreas libres de turismo porque los albergues de oviposición son muy limitados en el "bosque alto" que no inunda. En vez de mudar, se quedan cerca de los posos creados por el hombre.
En consecuencia, la abundancia de reptiles en CAPH, y la abundancia y diversidad en EI, siguen los resultados de los anuros. Donde hay más anfíbios, sus depredadores, las lagartijas y culebras, son más abundantes y diversas también. Se puede explicar este patrón fácilmente por disponibilidad de presa. El único albergue que no se puede explicar hasta este punto es SACHA. La diversidad y abundancia relativa de reptiles de las parcelas de tratamiento son más altas que las parcelas de control, pero sin un patrón similar de los anfíbios. Explicaremos los patrones de SACHA con los análices más adelante.
Además de los resultados de gran escala de la comunidad, algunas especies individuales son afectadas también por el turismo. En casí todos los casos, las especies que son afectadas por el turismo tienen abundancia más alta en las áreas de tratamiento. Solamente Epipedobates pictus de TRC es más abundante en las áreas de control. Las especies de EI siguen a los resultados anteriores de tratamiento versus control. En efecto, observamos congregaciones de Physalaemus petersi cantando y poniendo huevos en "Trocha Principal" en EI para varias noches al inicio de la época lluviosa. En otras zonas de las trochas de EI, machos de Colostethus marchesianus depositan sus renacuajos directamente en la trocha. A veces se ven juveniles recién metamorfoseados saliendo de los positos en la trocha.
En SACHA y TRC los microhylidos Ctenophryne geayi y Chiasmocleis bassleri son más abundantes en las áreas de tratamiento. Adicionalmente, Dendrophryniscus minutus y Phyllonastes myrmecoides son más abundantes en áreas de tratamiento en EI y SACHA, respectivamente. Una razón probable de su presencia significativa en las áreas de tratamiento es que estas cuatro especies son especialistas en hormigas (Rodríguez y Duellman 1994) que prefieren vivir cerca de las trochas turísticas donde las hormigas de muchas especies (e.j. Atta y Echiton) utilizan comúnmente las trochas hechas por el hombre (Doan, obs. pers.). Igualmente, muchos bufonidos prefieren las trochas para su locomoción debido a su tamaño grande; los resultados de Bufo cf. typhonius concuerdan con esta noción.
Las especies de Tambopata del género Epipedobates (femoralis, pictus y trivittatus) prefieren el microhábitat específico de los árboles caídos (Duellman 1978, obs. pers.). Cuando se crean las trochas turísticas generalmente evitan las caídas de árboles porque los dueños y guías del albergue prefieren mostrar a los turistas lo que consideran bosque " prístino ", sin huecos en el dosel. Por evitar el microhábitat del árbol caído, están evitando inadvertidamente áreas donde las ranas dardo venenosas son abundantes, según lo mostrado en el TRC donde Epipedobates pictus es muy abundante.
Los análices de trocha utilizan un acercamiento muy diferente que los análices anteriores, examinando el turismo usando solamente las parcelas de tratamiento, y buscando los efectos en la escala más pequeña de la misma parcela. No se encontró ningún efecto significativo sobre la riqueza de especies de anuros, apoyando la afirmación de Ludwig y Reynolds (1988) que la riqueza de las especies no es una medida adecuada de una comunidad. SACHA era el único albergue a diferenciar significativamente en biodiversidad entre los quadrats de trocha y quadrats fuera de la trocha. Sin embargo, los índices de semejanza en SACHA, resultan con una alta semejanza entre la comparación de los quadrats de trocha versus fuera de trocha. Como los índices de semejanza no tienen distribuciones estadísticas de probabilidades asociadas a ellos (Krebs 1999), una confianza más alta se debe poner en las pruebas de Mann-Whitney U que en los índices de semejanza. Por lo tanto, la diferencia significativa de la diversidad en SACHA es un resultado importante. Sin embargo, ninguna especie individual tenía abundancias significativamente diferentes en los quadrats de trocha versus fuera de trocha.
La única especie que tenía abundancia significativamente diferente entre los quadrats de trocha y no-trocha era el microhylido Hamptophyrne boliviana en ECO. Como la segunda especie más abundante de ECO (detrás de Adenomera andreae) esto es un resultado muy importante. De acuerdo a las especies de microhylidos arriba mencionadas, es probable que este especialista en hormigas se quede cerca de las trochas abiertas donde muchas especies de hormigas siempre transitan.
Se puede comparar directamente los resultados de semejanza de tratamiento versus control y trocha versus fuera de trocha para determinar que efecto de turismo (escala grande o escala pequeña) es más importante para la estructuración de las comunidades de herpetofauna en Tambopata (Tablas 4.3 y 4.8). En tres albergues (CAPH, EI y TRC) el efecto de tratamiento versus control es el más grande mientras que los efectos en ECO y SACHA están en la escala de la parcela, trocha versus fuera de trocha. Debido a estos resultados dictamos, no se puede hacer en este momento ninguna acepción general de cual es el efecto más importante.
Ni las regresiones lineales en grupo de distancia de la trocha ni las pruebas de los grupos de trocha en especies individuales fueron significativas estadísticamente. Este resultado puede significar una de dos posibilidades: 1) la distancia real de la trocha no es un factor importante, o 2) el muestreo era inadecuado para determinar tal efecto. En el caso del análisis de grupos de trocha es posible que el muestreo censó ligeramente las parcelas elegibles para contener una cantidad suficiente de individuos para este tipo de análisis. Como lo mencionado arriba, los sistemas de trocha de algunos albergues serpentearon tanto que llenaron casí toda la parcela, anulándolos del análisis. En otros albergues la heterogeneidad del hábitat y el sistema de trocha causaron que la parcela contuviera demasiado poco la trocha para utilizarla en el análisis de grupos de trocha. Debido a la cantidad limitada de las parcelas disponibles, no se encontró ningún resultado significativo. Para intentar contradecir el problema de tamaño de muestra por grupo de trocha, todos los quadrats por grupo de trocha por albergue fueron agrupados en una combinación para otro análisis. Cuando los albergues fueron utilizados como réplicas, nuevamente no se encontró ningún resultado significativo. Por lo tanto, o los efectos de la distancia de la trocha están localizados pero no detectables en estos análisis o no hay un efecto de la distancia de la trocha.
Las cantidades anuales de turistas pueden producir un efecto sobre la diversidad de la herpetofauna local. Aunque la regresión no era significativa (p = 0.078), una correlación de los mismos datos muestra una relación negativa fuerte (Pearson = -0.900) de cantidades anuales de turistas y la diversidad de tratamiento. Como fue concluido con los datos de tratamiento versus control, los albergues con altas cantidades de turistas por muchos años (por ejemplo EI y CAPH) son afectados por la presión de turismo.
La estructura fija de los programas turísticos actuales en cuatro de los albergues usados en este estudio permite al turista la poca oportunidad de ver los anfíbios y reptiles. Tres albergues ofrecen un viaje nocturno en bote para mirar caimanes, que de vez en cuando dan a los turistas observación de la especie herpetofauna más grande del área. Las ranas, lagartijas y culebras son vistas muy pocas veces y la opinión general de la gerencia y personal de los albergues es que los turistas tienen poco interés en ver los otros representantes de estas dos clases. Sin embargo, se debe mencionar, que el TRC, ofrece caminatas nocturnas con el propósito expreso de buscar los reptiles y anfíbios. El cuestionario de herpetofauna nos proporciónó algunos resultados inesperados que se pueden utilizar para modificar programas actuales de turismo para incluir oportunidades de ver el herpetofauna más críptica. Como se puede concluir en la Figura 3, los turistas desean ver los anfíbios y reptiles, y participarían en una caminata de noche si fuera ofrecida. Finalmente, setenta y tres por ciento de los turistas evaluados solicitan que en el albergue exista mayor información disponible sobre las serpientes venenosas.
Conclusiones
De la discusión anteriormente presentada se puede observar que el turismo, de hecho, tiene un impacto en las comunidades de herpetofauna. Confiamos en esta conclusión debido al período largo de estudio, a las metodologías rigurosamente aplicadas y del extenso e intenso muestreo. Los factores trás los efectos del turismo estan relacionadas con la intensidad y el período del turismo en un área. De los datos herpetológicos de TReeS-RAMOS Proyecto Tambopata podemos ver que, para los anfíbios, la diversidad disminuye con el tiempo y con la mayor cantidad de turistas que utilizan las trochas. Después de un período largo, los anfíbios o emigran a otras áreas que no estén bajo la presión turística o, en el caso del bosque alto de EI, permanecen en áreas de turismo, pero probablemente tienen una aptitud reproductiva muy baja.
Por otra parte, la presencia de la trocha turística no es necesariamente perjudicial. Para muchas especies la trocha sirve como área conveniente para buscar su presa ya que tanto la vegetación como la hojarasca estan reducidas. Los anfibios representan presa para ciertas especies de serpientes y lagartijas. Además, las trochas facilitan el movimiento de las especies grandes, incluyendo los sapos bufonidos y las serpientes.
Para manejar la alta diversidad de herpetofauna, los resultados de este estudio sugieren varias estrategias que la administración de los albergues deban poner en ejecución. Una, es esparcir el impacto de los turistas reduciendo el uso de las actuales trochas, que se podria hacer en combinación con la implementación de nuevas trochas y un mejor manejo de las rutas, con la finalidad que ningún área de bosque sea utilizada excesivamente. Además deben proporcionar más opciones a los turistas y a los guías, en cuanto al tipo de excursión, para eliminar la necesidad de utilizar un área de bosque o sección de trocha sucesivamente con la mayoría de grupos. Un problema con estas estrategias es que, aunque pueden beneficiar a la herpetofauna, númerosos investigadores (Mieczkowski 1995, Hunter y Green 1995) han demostrado que aperturar nuevas trochas causa daño a la vegetación. Creando muchas trochas de bajo-impacto podria ser más perjudicial al bosque que pocas trochas de alto impacto. Otra posibilidad, en lugar de esto seria tener una o dos rutas turísticas con un camino elevado y bien mantenido. Estos caminos pueden ser de madera, metal u otro material que eleve el turista sobre el suelo y no dañe excesivamente la vegetación. Además esto bajaria los efectos de la erosión del suelo y la formación de charcos, la perturbación de la vegetación que puede ser una de las razones principales de por qué huyen algunas especies de anfíbios.
Los resultados del cuestionario también sugieren implicaciones para el manejo de turistas. Las caminatas de noche expondrían los turistas a una mayor diversidad de reptiles, anfíbios y otras especies nocturnas, que elevaría su conocimiento de la selva. Los albergues no deben enjaular o mantener ninguna especie de herpetofauna como mascota, y a los caimanes no se les debe capturar durante las excursiones nocturnas, ya que los animales deben permanecer en su ambiente natural y porque los turistas se oponen drasticamente a tales prácticas. Los albergues deben proporcionar a los turistas mayor información en general sobre la herpetofauna de la zona, específicamente sobre serpientes venenosas, ya que los turistas tienen interés en aprender sobre esta y una mayor comprensión de las especies que habitan la selva de Tambopata enriquecería su estadía.