Hábitat
Un total de 1,386 árboles fueron enumerados y medidos durante los muestreos de hábitats en las estaciones de conteo. La Tabla 5.1 muestra los valores promedios de todas las variables registradas en cada tipo de bosque. Todas las variables difieren significativamente entre los cinco albergues (one-way ANOVA, * p < 0.05, tamaño de muestra no igual, valores de promedios armónicos usados). La densidad de árboles era mayor en el tipo F y en tipo E/F, con 177 árboles/hectárea en SACHA y TRC, respectivamente, mientras el tipo B de ECO tenia una densidad mínima. Los árboles más grandes y más altos se registraron en tipo B, mientras que los árboles más pequeños con dbh menores se registraron en el tipo H. Este último tipo también tuvo un dosel más abierto y tenía una alta densidad de vegetación en el subdosel. La cobertura del dosel era más completo en tipos E/F (TRC), que también tenía una menor cobertura de vegetación a nivel del suelo.
El tipo E/F (TRC) tenia el mayor porcentaje de palmeras (31.9%) mientras que el tipo H (EI) tenía el menor porcentaje (10.2%). Menos árboles, 28.8% del total de los árboles registrados en E/F (TRC), soportaban lianas u otras trepadoras, mientras el 51.9 % de árboles en tipo B (ECO) también las tenían.
Tipo F (SACHA) tenia la mayor cobertura de vegetación al nivel del suelo y en el sotobosque en general. Esto es probablemente resultado de la presencia del bambú Guadua weberbauerii. La Tabla 2 es un resúmen de las características de bambú en el sotobosque de SACHA. Este hábitat cubre aproximadamente un área de 1.63 km2 cerca el albergue. Con un promedio de 5.4 tallos por cada quadrat de 8 x 8m, el bambú cuenta con el 6.3 % de todos los tallos > 0.02 m dbh en cada quadrat. Esto sugiere que el bambú de SACHA es menos denso que en los tipos de bosque dominados de bambú en EI y TRC. El promedio de dbh de tallos de bambú en SACHA era 0.03 m, que también sugiere que el bambú de este albergue es más joven que el encontrado en tipo E/F en TRC (ver Kratter, 1995b). Estos resultados no significa que el bambú es menos importante en SACHA que el bambú de TRC. La gama de especies indicadoras de bambú presentes en SACHA es similar a TRC, aunque muchos de los cuales no eran registradas en los hábitats con bambú en EI.
Tabla 5.3 muestra los resultados de la clasificación de la arquitectura de los árboles. Bosque de tipo B en ECO muestra los niveles de mayor perturbación. Árboles con cicatrices, resultado de ramas caídas, eran más abundantes en este tipo de bosque, también fue alto el porcentaje de árboles en regeneración. Tipo H en EI y el tipo B/F en CAPH también muestran niveles de perturbación alta. Los bosques de SACHA y TRC son estructuralmente indicativos de hábitats primarios sin mucha perturbación. Estos dos albergues contienen un porcentaje mínimo de árboles que brotan ramas a una altura baja y contienen un porcentaje reducido de árboles con cicatrices producto de ramas caídas. Mayor evidencia que apoya la noción que el tipo B experimenta niveles de perturbación más altos se pueden observar en la Tabla 5.4. En visitas consecutivas a este tipo de bosque durante 12 meses, unos cuantos árboles bajo monitoreo habían muerto. Un incremento en el número de árboles muertos se registró en cuatro de los cinco tipos de bosque durante 12 meses (Tablas 5.5 - 5.7). El número más alto de árboles muertos se registró en tipo H (Tabla 5.8). En en tipo B, hubo un incremento en la mortalidad durante el periodo del estudio, con el porcentaje total de árboles muertos incrementando en un 4%. Tal como EI, se muestrearon los árboles en CAPH en sólo dos ocasiones, una en enero y el otro en junio de 1998. Aunque en el caso de CAPH este periodo resultó en un incremento relativamente grande en el número de árboles muertos (Tabla 5.7). No es conocido si este incremento hubiera sido registrado en una visita más temprano al albergue. Hay que emfatizar que hay que ser cauteloso en la interpretación de estas conclusiones, ya que el número de muestreos consecutivos es relativamente bajo.
Tablas 5.4 – 5.6 muestran los datos de número de árboles en fructificación y floración durante 3 o más visitas a ECO, SACHA, y TRC, respectivamente. Como fue mencionado antes, sólo se efectuó dos visitas de muestreo de hábitat a los otros dos albergues. Aunque los datos respectivos son detallados en las Tablas 5.7 y 5.8, el número de visitas fue insatisfactoria para explicar el patrón de fructificación en los tipos de bosque muestreado.
En tipo B, ECO, el número de árboles fructificando fue mayor en Setiembre 1998, que también correspnde al mes cuando hubo más floración (Tabla 5.4). 19% de los árboles registrados en flor eran palmeras. El número de árboles en fructificación registrado en cada visita eran casí significativamente diferente (one-way ANOVA, F = 2.232, p = 0.08), mientras no hubo una diferencia significativa en el número de árboles en flor (F = 1.493, p = 0.215).
En tipo F, en SACHA, el mayor número de árboles en fruto se registro en Mayo 1998, mientras en Julio la mayoría estaban en flor. De los árboles en fruto, 52% eran palmeras. El número de árboles en fructificación registrado en cada visita era casí significativamente diferente (F = 2.480, p = 0.08), mientras que hubo una diferencia significativa en el número de árboles en flor (F = 6.698, p < 0.05). Una diferencia significativa en el número de árboles en flor entre visitas al tipo E/F de TRC también se encontró (F = 5.777, p < 0.05), mientras que no hubo una diferencia significativa en el número de árboles en fruto (F = 0.368, p = 0.692). El número de árboles fructificando fue mayor en la segunda visita, Febrero 1998, 80% de los cuales eran palmeras. El número mayor de árboles en flor se registro en Agosto 1998.
Tabla 5.1. Densidad de árboles y promedios de las variables de hábitat registrados en cada uno de los cinco tipos de bosque. Tipo B: ECO; tipo B/F: CAPH; tipo H: EI; tipo F: SACHA; tipo E/F: TRC. Densidad expresado como número de árboles grandes por hectárea.* diferencias significativas entre tipos de bosque (one-way ANOVA, p < 0.05).
| Variable | . | CAPH |
EI |
ECO |
SACHA |
TRC |
| Tipo de Bosque | . | B/F |
H |
B |
F |
E/F |
| No. total de árboles | * |
240 |
300 |
289 |
297 |
260 |
| Densidad de árboles | * |
160 |
177 |
119 |
128 |
177 |
| No. de palmeras | * |
52 |
32 |
44 |
85 |
83 |
| Árboles con lianas (%) | * |
41.7 |
43.0 |
51.9 |
35.4 |
28.8 |
| Distancia desde punto de conteo (m) | * |
8.7 |
10.1 |
10.0 |
8.5 |
8.2 |
| Dbh (m) | * |
0.48 |
0.40 |
0.56 |
0.55 |
0.43 |
| Altura (m) | * |
19.1 |
18.5 |
23.4 |
19.4 |
20.6 |
| Cobertura del suelo (%) | * |
29.8 |
19.5 |
20.3 |
35.0 |
17.1 |
| Cobertura sotobosque (%) | * |
39.2 |
41.5 |
37.2 |
50.3 |
29.4 |
| Cobertura medio-dosel (%) | * |
48.1 |
62.5 |
55.5 |
58.2 |
28.8 |
| Cobertura dosel (%) | * |
67.1 |
56.0 |
57.5 |
58.8 |
68.6 |
Tabla 5.2. Promedios de variables de hábitat por cada quadrat (8m x 8m), registrado en sotobosque de tipo F dominado por bambú en SACHA.
| Variable | . | . | Variable | . |
| No. de quadrats | 30 | . | No. de tallos no-bambú | 80.7 |
| Área muestreado (m2) | 1,926 | . | No. de tallos no-bambú vivos | 76.0 |
| No. de tallos bambú | 5.4 | . | No. de tallos no-bambú muertos | 2.4 |
| No. tallos de bambú vivos | 4.0 | . | Dbh de tallos bambú | 0.03 |
| No. tallos de bambú muertos | 1.4 | . | . | . |
Tabla 5.3. Resultados de clasificación de arquitectura de árboles por albergue. N = número total de árboles medidos. % (entre parentices) de número total de árboles medidos en cada albergue.
| Variable | CAPH |
EI |
ECO |
SACHA |
TRC |
| Tipo de bosque | B/F |
H |
B |
F |
E/F |
| No. de árboles medidos(N) | 240 |
300 |
289 |
297 |
260 |
| No. que bifurcan más arriba que ½ altura | 215 |
249 |
235 |
276 |
234 |
| % que bifurcan más arriba que ½ altura | 90 |
83 |
81 |
93 |
90 |
| No. y (%) más arriba con cicatrices | 2 (1) |
11 (4) |
17 (6) |
2 (1) |
1 (1) |
| No. y (%) más arriba sin cicatrices | 213 (89) |
238 (79) |
218 (75) |
274 (92) |
233 (90) |
| No. que bifurcan más abajo que ½ altura | 24 |
36 |
46 |
11 |
16 |
| % que bifurcan más abajo que ½ altura | 10 |
12 |
16 |
4 |
6 |
| No. y (%) más abajo con cicatrices | 3 (1) |
3 (1) |
38 (13) |
0 (0) |
2 (1) |
| No. y (%) más abajo sin cicatrices | 21(9) |
33 (11) |
8 (3) |
11 (4) |
14 (5) |
Tabla 5.4. Resúmen de datos de fructificación y el número de árboles muertos registrados en tipo B, entre Octubre 1997 y Setiembre 1998, en ECO.
Oct. 97 |
Mar. 98 |
Jun. 98 |
Set. 98 |
|
| No. de árboles en fruto (incl. Palmeras) | 10 |
12 |
20 |
21 |
| No. total de árboles en flor | 10 |
4 |
10 |
12 |
| No. total de palmeras en fruto | 4 |
8 |
8 |
4 |
| No. de árboles muertos | 8 |
11 (+3) |
13 (+2) |
14 (+1) |
Tabla 5.5. Resúmen de datos de fructificación y el número de árboles muertos registrados en tipo F, entre Noviembre 1997 y Julio 1998, en SACHA.
Nov. 97 |
May. 98 |
Jul. 98 |
|
| No. de árboles en fruto (incl. Palmeras) | 24 |
42 |
35 |
| No. total de árboles en flor | 15 |
14 |
36 |
| No. total de palmeras en fruto | 19 |
22 |
22 |
| No. de árboles muertos | 10 |
10 (+0) |
13 (+3) |
Tabla 5.6. Resúmen de datos de fructificación y el número de árboles muertos registrados en tipo E/F, entre Diciembre 1997 y Agosto 1998, en TRC.
| Dic. 97 | Feb. 98 | Ago. 98 | |
| No. de árboles en fruto (incl. Palmeras) | 38 | 44 | 36 |
| No. total de árboles en flor | 11 | 11 | 28 |
| No. total de palmeras en fruto | 32 | 35 | 18 |
| No. de árboles muertos | 7 | 11 (+4) | 11 (+0) |
Tabla 5.7. Resúmen de datos de fructificación y el número de árboles muertos registrados en tipo B/F, en CAPH durante dos visitas distintas en 1998.
| Ene. 98 | Jun. 98 | |
| No. de árboles en fruto (incl. Palmeras) | 1 | 13 |
| No. total de árboles en flor | 2 | 5 |
| No. total de palmeras en fruto | 1 | 2 |
| No. de árboles muertos | 1 | 5 (+4) |
Tabla 5.8. Resúmen de datos de fructificación y el número de árboles muertos registrados en tipo H, en EI durante dos visitas distintas entre Octubre 1997 y Abril 1998 en 1998.
| Oct. 97 | Abr. 98 | |
| No. de árboles en fruto (incl. Palmeras) | 12 | 29 |
| No. total de árboles en flor | 10 | 8 |
| No. total de palmeras en fruto | 5 | 19 |
| No. de árboles muertos | 15 | 15 (+0) |
Similitudes entre Tipos de Bosque
La Figura 5.1 muestra los resultados de un Análisis de Funcción Discriminatoria (AFD) sobre las variables de hábitat medidas en cada uno de los tipos de bosque. AFD se usó en este caso ya que los cinco tipos de bosque eran predeterminados y basados en el estudio de Phillips (1993). Para analizar la similitud entre los tipos de bosque basadolo en variables de hábitat, todas las variables se análizaron para ver si existía diferencias significativas (one-way ANOVA, p < 0.05 ver Tabla 5.1). De allí se utilizó un Análisis de Componentes Principales (ACP) para fusiónar las variables en grupos más pequeños de "componentes derivados" (Fielding, 1997). El resultado del ACP fue la reducción de los datos de variables a cinco componentes, que explica el 65.4% de la variabilidad. De allí se utilizó nuevamente el AFD para indicar las asociaciones respectivas.
El AFD produjó cuatro funciones discriminatorias. Los primeros dos, AFD1 y AFD2 explican el 60.0% y 26.4% de la variabilidad. El AFD1 tuvo un puntaje positivo y alto para el ACP dominado por altura de árboles (árboles muy altos), porcentaje estimado de cobertura de dosel (denso y cerrado), porcentaje estimado de cobertura en el sotobosque (sotobosque denso) y número de palmeras (número alto de palmeras), mientras que se mantenga un puntaje alto en la altura del árbol (punto de bifurcación). El AFD2 tuvo un puntaje positivo y alto en cuanto a altura de árboles, y un puntaje negativo y alto en cuanto a la altura del dosel, y número de palmeras (número pequeño de palmeras).
Los componentes del PCA y estas dos funciones se graficaron para producir la figura 5.1. Examinando las agrupaciones y centroides de cada agrupación se puede ver que los tres tipos de bosque B/F (CAPH), tipo F (SACHA), tipo E/F (TRC), son los más similares. Los tipos de bosque más distintos es decir más diferentes, son tipo H (EI) y tipo B (ECO).
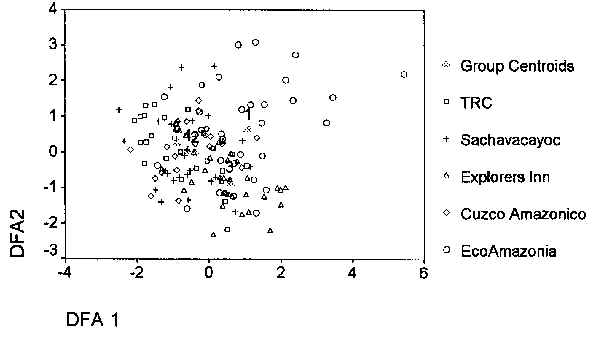
Figura 5.1. Perfil de hábitat, AFD1 vs AFD2.
Muestreos de Aves
Un total de 259 especies de aves fueron registradas durante 336 días de muestreos de PCV, TLDV, mallas de neblina y conteos visuales en los cinco albergues. De este total, 233 especies fueron registradas durante muestreos de PCV, que corresponde a 18,107 contactos con 21,453 individuos. De estos contactos, 94.4% fueron de tipo auditivo; los contactos visuales sólo llegaron a ser 1.1%, mientras el total de aves vistas y escuchadas llegó a 4.5% de todos los contactos. El 29.8% de los contactos eran aves registradas a la altura del subdosel, 23.0% a la altura del dosel, 21.7% a la altura del sotobosque, mientras aves terrestres constituyeron 18.1%. El resto de aves, 7.5%, se registraron volando sobre el dosel.
En TRC 180 especies se registraron durante muestreos PCV, mientras 179, 177, 172 y 144 especies fueron registradas en SACHA, EI, ECO, y CAPH respectivamente. La Figura 5.2 muestra las curvas de acumulación de especies para muestreos de PCV en cada albergue. Después de 336 días de muestreo el número de especies nuevas reportadas en cada albergue aún no habia llegado a una asíntota indicando que con las métodologias utilizadas en los tipos de bosque muestreados todavía existen especies que no fueron registrados. Tabla 5.9 muestra el número de individuos registrado en diferentes visitas durante el período de 21 meses. El mayor número de individuos se registró en el mes de Mayo 1997 y 1998, en los tipos B y F de ECO y SACHA, respectivamente. El mayor número de individuos se registró en el mes de Abril 1997, en tipo B/F de CAPH, en Febrero 1998 en tipo E/F de TRC y en Octubre 1997, en tipo H, en EI.
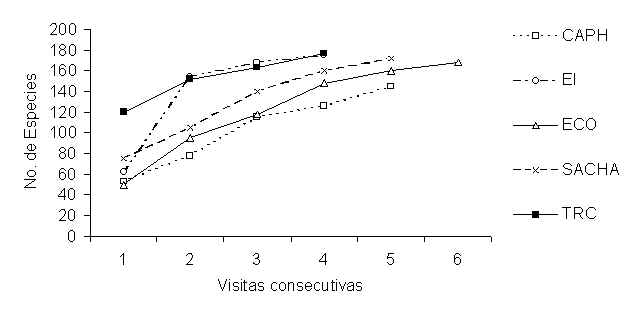
Figura 5.2. Curvas de acumulación de especies PCV).
Tabla 5.9. El número total de individuos registrados durante muestreos PCV durante cada visita a los albergues.
| Albergue | Mes |
Año |
Visita |
No. de Individuos |
| ECO | Ene. |
1997 |
1 |
562 |
| . | May. |
1997 |
2 |
1389 |
| . | Oct. |
1997 |
3 |
758 |
| . | Mar. |
1998 |
4 |
1275 |
| . | Jun. |
1998 |
5 |
777 |
| . | Set. |
1998 |
6 |
1144 |
| CAPH | Feb. |
1997 |
1 |
525 |
| . | Abr. |
1997 |
2 |
970 |
| . | Ago. |
1997 |
3 |
779 |
| . | Feb. |
1998 |
4 |
315 |
| . | Jun. |
1998 |
5 |
605 |
| EI | Mar. |
1997 |
1 |
537 |
| . | Jul. |
1997 |
2 |
697 |
| . | Oct. |
1997 |
3 |
1125 |
| . | Abr. |
1998 |
4 |
939 |
| SACHA | Mar. |
1997 |
1 |
782 |
| . | Jun. |
1997 |
2 |
1128 |
| . | Nov. |
1997 |
3 |
1233 |
| . | May. |
1998 |
4 |
1306 |
| . | Jul. |
1998 |
5 |
512 |
| TRC | Ago. |
1997 |
1 |
629 |
| . | Dic. |
1997 |
2 |
1018 |
| . | Feb. |
1998 |
3 |
1186 |
| . | Ago. |
1998 |
4 |
684 |
Densidad de Poblaciones de Aves
Tablas 5.10 – 5.17 muestran las densidades poblacionales de las especies indicadoras de los tipos de bosque muestreados. Se registraron 11 especies indicadoras de tipo B en ECO, mientras que actualmente 13 especies fueron registradas en este albergue, 2 más usando diferentes métodos (Tabla 5.10). Aunque, las densidades se calcularon sólo para cuatro de estas. En el tipo B/F (CAPH), 7 especies se registraron durante los muestreos, 8 fue el número total para este albergue (Tabla 5.11). Cinco especies no fueron reportados por ningún método en este tipo de bosque durante el estudio. Otras, densidades se calcularon para sólo cuatro especies las mismas que en ECO. De estas sólo C. cinereus era más abundante en tipo F/B.
De las 35 especies indicadoras de selva baja, más especies fueron registradas durante los muestreos en el bosque primarío de SACHA y TRC (31 y 30, respectivamente; 33 especies en total para los dos albergues) que en los otros albergues, aunque la diferencia es pequeña (Tablas 5.12 - 5.15). El mayor número de poblaciones donde sus densidades se podría calcular era para tipo H (EI) (21 especies, Tabla 5.13), con el menor número en tipo B/F (CAPH) (Tabla 5.12). Las densidades más bajas calculadas en CAPH es probablemente debido a un área reducida de hábitat disponible y la presencia de tipo B como un componente en el mosaico.
Examinando las tablas de densidades y usando la información de los hábitats, podemos concluir varíos puntos relacionados con la abundancia de aves indicadoras del bosque bajo, estos son resumidos a continuación (más información también esta contenido en la sección sobre ‘Descripciones de Especies’). Varias especies claramente tienen una preferencia para el bosque tipo H. Estas incluyen T. guttatus, C. variegatus, S. rutilans, T. aethiops, P. rubrocapilla, mientras otras especies, M. menetreisii, M. myotherinus, aunque encontrados en los cuatro tipos de bosque son más abundantes en el tipo H. Dos especies, C. nobilis y M. campanisoma, tienen densidades altos en los bosques primaríos de SACHA y TRC. Estas especies prefieren bosque de terraza donde son más abundantes que en bosques perturbados. C. torquata, es abundante en todos los tipos, y también tiene densidades más altos en los bosques primaríos. Aunque, O. stellatus era mucho más abundante en los tipos más perturbados B y B/F. D. cincta se ocurre en todos los tipos aunque siempre en densidades bajas y es naturalmente rara. Otras especies muestran una preferencia por los tipos de bosque inundados (D. certhia, S. caudacutus, F. colma, P. coronatus, H. rubrica) mientras otras los evitan (D. fuliginosa, X. spixii, M. marginatus).
Un total de 14 y 13 indicadoras de bambú fueron registradas durante PCV en hábitats con Guadua en SACHA y TRC respectivamente. En total hubo 17 y 19 especies registradas en estos albergues (Tablas 5.16 y 5.17). Una especie no fue reportado en ninguno de los dos albergues; Sporophila schistacea. Probablemente no fue reportado debido a la ecología de esta, un consumidor de semillas fuertemente amarrado a la ecología de floración de Guadua. Densidades se calcularon para 12 especies indicadores de bambú en SACHA, mientras era sólo posible calcular las densidades de la mitad de este número en TRC. La rázon por esto era que sólo cinco estaciones en el TRC estaban localizadas en tipo E/F con bambú, mientras la mitad de las estaciones en SACHA contenían bambú. De las seis especies, C. santaemariae y S. ucayalae se encontraron que eran más densos en E/F con bambú que en tipo F con bambú.
Tabla 5.10. Densidades para las especies indicadoras de Bosque Tropical Inundado, en tipo B (ECO). Densidad: + registrado durante PCV, pero número insuficiente de contactos para determinar densidad; - no registrado durante PCV, pero registrado con otros metodos; x sólo registrado en mallas de neblina; NR no reportado en el albergue. Método de Muestreo: PCV Plot Circular Variable; TLDV Transecto de Linea de Distancia Variable; MN Mallas de Neblina; CD Conteos Directos; OG Observaciones Generales.
| Especies Indicadoras de Bosque Tropical Inundado en ECO – Tipo B | Densidad Poblacional |
Método de Censo |
| Crypturellus bartletti | 25.9 |
TLDV |
| C. cinereus | 0.6 |
PCV |
| Agami agami | - |
OG |
| Leucopternis schistacea | + |
CD |
| Aramides cajaena | + |
TLDV |
| Eurypyga helias | - |
OG |
| Phaethornis hispidus | 6.7 |
MN |
| Hypocnemoides maculicauda | + |
PCV |
| Sclateria naevia | + |
PCV |
| Hylophylax punctulata | X |
MN |
| Schiffornis major | + |
PCV |
| Pipra fasiicauda | 35.2 |
PCV, MN |
| Turdus hauxwelli | 30.7 |
PCV |
Tabla 5.11. Densidades de poblaciones para las especies indicadoras de Bosque Tropical Inundado, en tipo B/F (CAPH).
| Especies Indicadoras de Bosque Tropical Inundado en CAPH – Tipo B/F | Densidad Poblacional |
Método de Censo |
| Crypturellus bartletti | 19.9 |
PCV |
| C. cinereus | 0.6 |
PCV |
| Agami agami | NR |
OG |
| Leucopternis schistacea | + |
CD |
| Aramides cajaena | - |
OG |
| Eurypyga helias | NR |
OG |
| Phaethornis hispidus | 4.6 |
MN |
| Hypocnemoides maculicauda | NR |
PCV |
| Sclateria naevia | + |
PCV |
| Hylophylax punctulata | NR |
MN, OG |
| Schiffornis major | NR |
PCV |
| Pipra fasiicauda | 9.8 |
PCV, MN |
| Turdus hauxwelli | 9.4 |
PCV |
Tabla 5.12. Densidades de poblaciones para las especies indicadoras de Bosque Bajo, en tipo B/F (CAPH).
| Especies Indicadoras de Bosque Bajo en CAPH – Tipo B/F | Densidad Poblacional |
Método de Censo |
| Tinamus guttatus | - |
OG |
| Crypturellus strigulosus | NR |
PCV, OG |
| C. variegatus | + |
PCV |
| Micrastur gilvicollis | + |
CD |
| Mitu tuberosa | NR |
PCV |
| Odontophorus stellatus | 11.0 |
PCV |
| Psophia leucoptera | - |
OG |
| Phaethornis phillipii | x |
MN |
| Malacoptila semicincta | 1.3 |
MN |
| Dendrocincla fuliginosa | 0.6, 1.3 |
PCV, MN |
| D. merula | 15.8 |
MN |
| Dendrocolaptes certhia | 1.5 |
PCV |
| Xiphorhynchus spixii | + |
PCV, MN |
| Synallaxis rutilans | + |
PCV |
| Automolus infuscatus | 1.3 |
PCV |
| Sclerurus caudacutus | 0.5, 2.0 |
PCV |
| Thamnophilius aethiops | NR |
PCV, OG |
| Thamnomanes ardesiacus | 3.0 |
PCV |
| Myrmotherula haematonata | 4.6 |
MN |
| M. hauxwelli | 7.0 |
PCV |
| M. leucophthalma | 4.1, 4.1 |
PCV, MN |
| M. longipennis | 4.6 |
PCV |
| M. menetriesii | 2.0 |
PCV |
| Dichrozona cincta | + |
PCV, MN |
| Myrmoborus myotherinus | 10.5 |
PCV |
| Gymnopithys salvini | 12.5 |
MN |
| Formicarius colma | NR |
PCV, OG |
| Chamaeza nobilis | - |
OG |
| Myrmothera campanisoma | + |
PCV |
| Conopophaga peruviana | 5.3 |
MN |
| Corythopis torquata | 14.7 |
PCV |
| Platyrinchus coronatus | 10.5 |
PCV |
| Pipra rubrocapilla | NR |
PCV, OG |
| Microcerculus marginatus | 0.5 |
PCV |
| Habia rubrica | 5.3 |
PCV |
Tabla 5.13. Densidades de poblaciones para las especies indicadoras de Bosque Bajo, en tipo H (EI).
| Especies Indicadoras de Bosque Bajo en EI – Type H forest | Densidad Poblacional |
Método de Censo |
| Tinamus guttatus | 13.3 |
PCV |
| Crypturellus strigulosus | NR |
OG |
| C. variegatus | 1.8 |
PCV |
| Micrastur gilvicollis | + |
CD |
| Mitu tuberosa | - |
OG |
| Odontophorus stellatus | 40.8 |
PCV |
| Psophia leucoptera | - |
OG |
| Phaethornis phillipii | X |
MN |
| Malacoptila semicincta | + |
MN |
| Dendrocincla fuliginosa | 2.7, 2.7 |
PCV, MN |
| D. merula | 10.5 |
MN |
| Dendrocolaptes certhia | 0.6 |
PCV |
| Xiphorhynchus spixii | 1.9 |
PCV |
| Synallaxis rutilans | 7.4 |
PCV |
| Automolus infuscatus | 2.5 |
PCV |
| Sclerurus caudacutus | 3.0 |
MN |
| Thamnophilius aethiops | 8.0 |
PCV |
| Thamnomanes ardesiacus | 2.4 |
PCV |
| Myrmotherula haematonata | NR |
PCV, MN, OG |
| M. hauxwelli | 8.9 |
PCV |
| M. leucophthalma | 3.5 |
MN |
| M. longipennis | 10.9 |
PCV |
| M. menetriesii | 11.1 |
PCV |
| Dichrozona cincta | + |
PCV |
| Myrmoborus myotherinus | 48.0 |
PCV |
| Gymnopithys salvini | 1.1, 16.0 |
PCV, MN |
| Formicarius colma | 3.7 |
PCV |
| Chamaeza nobilis | NR |
PCV, OG |
| Myrmothera campanisoma | 2.5 |
PCV |
| Conopophaga peruviana | + |
MN |
| Corythopis torquata | 12.9 |
PCV |
| Platyrinchus coronatus | + |
PCV, MN |
| Pipra rubrocapilla | 5.7 |
PCV |
| Microcerculus marginatus | 12.3 |
PCV |
| Habia rubrica | 2.2 |
PCV |
Tabla 5.14. Densidades de poblaciones para las especies indicadoras de Bosque Bajo, en tipo F (SACHA).
| Especies Indicadoras de Bosque Bajo en SACHA – Tipo F | Densidad Poblacional |
Método de Censo |
| Tinamus guttatus | + |
PCV |
| Crypturellus strigulosus | NR |
OG |
| C. variegatus | + |
PCV |
| Micrastur gilvicollis | + |
CD |
| Mitu tuberosa | - |
OG |
| Odontophorus stellatus | + |
PCV |
| Psophia leucoptera | + |
TLDV, PCV |
| Phaethornis phillipii | - |
OG |
| Malacoptila semicincta | + |
PCV, MN |
| Dendrocincla fuliginosa | 1.2, 1.2 |
PCV, MN |
| D. merula | 4.0 |
MN |
| Dendrocolaptes certhia | 1.0 |
PCV |
| Xiphorhynchus spixii | 1.3 |
PCV |
| Synallaxis rutilans | + |
PCV |
| Automolus infuscatus | + |
PCV |
| Sclerurus caudacutus | 1.4, 1.3 |
PCV, MN |
| Thamnophilius aethiops | + |
PCV |
| Thamnomanes ardesiacus | 1.6 |
PCV |
| Myrmotherula haematonata | NR |
PCV, MN |
| M. hauxwelli | 7.4 |
PCV |
| M. leucophthalma | 2.3 |
MN |
| M. longipennis | 13.0 |
PCV |
| M. menetriesii | 5.1 |
PCV |
| Dichrozona cincta | + |
PCV |
| Myrmoborus myotherinus | 11.8 |
PCV |
| Gymnopithys salvini | 3.3 |
MN |
| Formicarius colma | 15.8 |
PCV |
| Chamaeza nobilis | 1.4 |
PCV |
| Myrmothera campanisoma | 6.0 |
PCV |
| Conopophaga peruviana | 4.5 |
MN |
| Corythopis torquata | 28.6 |
PCV |
| Platyrinchus coronatus | 3.9 |
PCV |
| Pipra rubrocapilla | + |
PCV |
| Microcerculus marginatus | 10.7 |
PCV |
| Habia rubrica | 4.2 |
PCV |
Tabla 5.15. Densidades de poblaciones para las especies indicadoras de Bosque Bajo en tipo E/F (TRC).
| Especies Indicadoras de Bosque Bajo en TRC– Tipo E/F | Densidad Poblacional |
Método de Censo |
| Tinamus guttatus | 1.6 |
PCV |
| Crypturellus strigulosus | NR |
OG |
| C. variegatus | NR |
PCV |
| Micrastur gilvicollis | + |
CD |
| Mitu tuberosa | 8.2 |
TLDV |
| Odontophorus stellatus | + |
TLDV, PCV |
| Psophia leucoptera | + |
TLDV, PCV |
| Phaethornis phillipii | - |
OG |
| Malacoptila semicincta | + |
PCV, MN |
| Dendrocincla fuliginosa | 0.6, 1.4 |
PCV, MN |
| D. merula | 1.8 |
MN |
| Dendrocolaptes certhia | 0.6 |
PCV |
| Xiphorhynchus spixii | 5.8 |
PCV |
| Synallaxis rutilans | + |
PCV |
| Automolus infuscatus | 3.0 |
PCV |
| Sclerurus caudacutus | 1.8 |
MN |
| Thamnophilius aethiops | + |
PCV |
| Thamnomanes ardesiacus | 4.3 |
PCV |
| Myrmotherula haematonata | + |
MN |
| M. hauxwelli | 11.1 |
PCV |
| M. leucophthalma | 1.8 |
MN |
| M. longipennis | 14.8 |
PCV |
| M. menetriesii | 3.8 |
PCV |
| Dichrozona cincta | 0.3 |
PCV |
| Myrmoborus myotherinus | 26.2 |
PCV |
| Gymnopithys salvini | 0.7, 10.1 |
PCV, MN |
| Formicarius colma | 5.4 |
PCV |
| Chamaeza nobilis | 1.6 |
PCV |
| Myrmothera campanisoma | 5.1 |
PCV |
| Conopophaga peruviana | 1.8 |
MN |
| Corythopis torquata | 30.1 |
PCV |
| Platyrinchus coronatus | + |
PCV |
| Pipra rubrocapilla | - |
OG |
| Microcerculus marginatus | 11.4 |
PCV |
| Habia rubrica | 6.7 |
PCV |
Tabla 5.16. Densidades de poblaciones para las especies indicadoras de Bambú en tipo F (SACHA).
| Especies Indicadoras de Bambú en SACHA– Tipo F | Densidad Poblacional |
Método de Censo |
| Dromococcyx pavoninus | - |
OG |
| Nonnula ruficapilla | - |
OG |
| Monasa flavirostris | - |
OG |
| Campylorhamphus trochilirostris | + |
PCV, MN |
| Simoxenops ucayalae | 4.7 |
PCV |
| Anabazenops dorsalis | + |
PCV |
| Automolus melanpezus | 12.4 |
PCV |
| Cymbiliaimus sanctaemariae | 6.2 |
PCV |
| Myrmotherula iheringi | 3.8 |
PCV |
| Myrmotherula ornata | 2.6 |
PCV |
| Microrhopias quixensis | - |
OG |
| Drymophila devillei | 42.3 |
PCV |
| Cecromacra manu | 3.7 |
CD |
| Percnostola lophotes | 48.3 |
PCV |
| Myrmeciza goeldii | 31.1 |
PCV |
| Poecilotriccus albifacies | 4.4 |
PCV |
| Hemitriccus flammulatus | - |
OG |
| Ramphotrigon fuscicauda | 4.3 |
PCV |
| R. megacephala | 13.1 |
PCV |
| Sporophila schistacea | NR |
OG |
Tabla 5.17. Densidades de poblaciones para las especies indicadoras de Bambú en tipo E/F (TRC).
| Especies Indicadoras de Bambú en TRC– Tipo E/F | Densidad Poblacional |
Método de Censo |
| Dromococcyx pavoninus | - |
OG |
| Nonnula ruficapilla | NR |
OG |
| Monasa flavirostris | - |
OG |
| Campylorhamphus trochilirostris | 1.4 |
MN |
| Simoxenops ucayalae | 7.7 |
PCV |
| Anabazenops dorsalis | + |
PCV |
| Automolus melanpezus | + |
PCV |
| Cymbiliaimus sanctaemariae | 8.1 |
PCV |
| Myrmotherula iheringi | + |
PCV |
| Myrmotherula ornata | + |
PCV |
| Microrhopias quixensis | - |
PCV, OG |
| Drymophila devillei | 6.7 |
OG |
| Cecromacra manu | - |
CD |
| Percnostola lophotes | 14.8 |
PCV |
| Myrmeciza goeldii | 21.5 |
PCV |
| Poecilotriccus albifacies | + |
PCV |
| Hemitriccus flammulatus | - |
OG |
| Ramphotrigon fusiicauda | + |
PCV |
| R. megacephala | 6.0 |
PCV |
| Sporophila schistacea | NR |
OG |
Asociaciones entre Especies y Hábitats
Perfiles de PCA de los cinco tipos de bosque se hicieron siguiendo un análisis de varianza (one-way ANOVA) de variables de hábitat por tipo. El número de características variables y componentes del hábitat variaron entre tipos de bosque. Los dos primeroes componentes en cada tipo (PCA1 y PCA2) cuenta por el mayor porcentaje de la variabilidad en el hábitat. PCA1 y PCA2 para cada tipo de bosque se graficaron para producir el perfil de cada albergue. El número promedio de las indicadoras registadas en cada estación también se grafico (Figuras 5.3 – 5.9). Asociaciones de hábitat para las indicadoras de bambú en el TRC no se realizo ya que existe publicaciones detallados de este fenómeno Kratter (1995b, 1997).
Figura 5.3. Patron de ACP para especies indicadoras de Bosque Inundando en ECO.
Figura 5.4. Patron de ACP para especies indicadoras de Bosque Inundando en CAPH.
Figura 5.5. Patron de ACP para especies indicadoras de Bosque Bajo en CAPH.
Figura 5.6. Patron de ACP para especies indicadoras de Bosque Bajo en EI.
Figura 5.7. Patron de ACP para especies indicadoras de Bosque Bajo en SACHA.
Figura 5.8. Patron de ACP para especies indicadoras de Bambú en SACHA.
Figura 5.9. Patron de ACP para especies indicadoras de Bambú en TRC.
Para el tipo B (ECO), el PCA1 y PCA2 explico 49.2% de la variabilidad del hábitat. Figura 5.3 muestra el perfil de hábitat correspondiente donde PCA1 representa la altura del dosel, mientras PCA2 representa el porcentaje de cobertura del dosel. Figura 5.3 muestra que la mayor número de especies indicadoras de bosque inundado que fueron registrados en áreas donde el dosel es de una altura media/alta, y donde la cobertura del dosel es más abierta, es decir menos denso.
En tipo B/F (CAPH), el PCA1 y PCA2 explicaron 48.6% de la variabildiad del hábitat, y representa la altura del dosel y el estimado de cobertura del mismo. La Figura 5.4 muestra los resultados del número promedio de especies indicadoras de bosque inundado registradas en cada estación. Un promedio más alto de especies indicadoras se registro en áreas donde el dosel era alto. También, en tipo B, la mayoría de indicadoras fueras registradas donde la cobertura del dosel era menos denso.
En el caso de las indicadoras de bosque bajo, el número promedio de especies se registraron en áreas donde el dosel era de altura mediana, con menos especies en áreas donde el dosel era bajo. Estas indicadoras también son tolerantes a la densidad del dosel, con muchas especies registradas en áreas de cobertura abierto como denso (Figura 5.5).
El PCA1 y PCA2 explicaron 58.4% de la variabildad de hábitat en tipo H (EI), con PCA1 representando el número (densidad) de palmeras, y PCA2 representando una estimación del porcentaje de cobertura en el subdosel. La Figura 5.6 muestra que un mayor número de especies indicadoras en tipo H fueron registradas donde hubo menos palmeras aunque son especies que puedan tolerar coberturas de subdosel denso como abierto.
En Sachavacayoc, el PCA1 y PCA2 explicaron 56.6% de la variabilidad en el hábitat. El PCA1 es número de tallos de bambú vivos, mientras el PCA2 es altura de dosel. La Figura 5.7 muestra que el mayor número de especies indicadoras de bosque bajo fueron registradas donde la altura del dosel era alto y la densidad de bambú bajo. Lo opuesto es el caso con especies indicadoras de bambú. Estas especies son las más restringidas en cuando a hábitat en Tambopata, y el mayor número de ellas se registraron donde la densidad de bambú era alto, y la altura del dosel era bajo (Figura 5.8). Esto indica que los especialistas están menos dependientes en la cobertura de vegetación, y son más dependientes en la densidad y estructura de los hábitats con bambú (ver también Kratter, 1995b).
En el ultimo albergue TRC, el PCA1 y PCA2 explicaron 56.3% de la variabildiad. El primer componente era altura de dosel, mientras la segunda era la densidad de vegetación a nivel del suelo. La Figura 5.9 muestra el número promedio de indicadoras de bosque bajo estos componentes. El mayor número de indicadoras se registraron en E/F donde el dosel era muy alto y donde la estimación de cobertura del suelo era menor (menos denso).
Frugívoros del Dosel
Dos familias de frugívoros que viven en el dosel, Psittacidae y Ramphastidae, son los más buscados por los turistas que visitan Madre de Dios (Munn, 1992; también ver ‘Cuestionarío Turístico’). Un total de 1632 contactos con loros, 9% de todos los contactos, se hicieron durante 21 meses de PCV. 1251 de estas eran contactos volando por encima del dosel. Entonces los loros representan 92.7% de todos los contactos de este tipo. Un total de 347contactos (21.3% de todos los contactos con loros) eran con las tres especies de guacamayo más grandes; Ara ararauna, A. macao y A. chloroptera. La Tabla 5.18 muestra que el número total de contactos para todos los loros y para estas tres especies de guacamayo. Es importante resaltar de todas maneras que el número de visitas a cada albergue no fue igual, aunque los resultados presentados son interésantes.
Un gran número de contactos con loros durante el estudio se hizo en tipo B (ECO). Aunque, el número de contactos con guacamayos en este lugar era bajo, y una reflección de la abundancia de especies que prefieren tipos de bosque más humedos como pantanos y aguajales. También existen muchas especies que consumen frutos de las palmeras, que abundan, e incluso duermen en ellas. El mayor número de contactos se registró en los bosques primaríos de SACHA y TRC, en ambos también hubo una mayor abundancia de guacamayos. Los resultados que generan más preocupación provienen de los áreas aparentemente más perturbados, EI y CAPH. Una menor cantidad de contactos se tuvo con loros en EI, mientras en CAPH muestra una falta alarmante de loros en general pero especialmente guacamayos.
Durante el período de Setiembre 1997 a Setiembre 1998, el número total de especies de loros registradas en todos los albergues era positivamente correlacionado significativamente con el número total de árboles en fruto (Correlación de Pearson, r = 0.587, p < 0 .05, a = 0.05). El número de loros volando también era más alto y positivamente correlacionado con la misma variable (r = 0.575, p < 0.05), indicando que más loros estaban visitando la región en búsqueda de frutos. El mismo patrón se puede ver cuando se examina sólo los datos para las tres especies de guacamayos más grandes. El número de guacamayos es positivamente correlacionado nuevamente (r = 0.787, p < 0.05) tal como es el número de individuos volando (r = 0.759, p < 0.05). Más guacamayos se registraron, y existe más actividad en ellos, cuando la abundancia de árboles en fruto es alto. Esto tiene importantes consecuencias para el monitoreo de loros en Tambopata así como para el turismo (ver Discussión).
No hubo un patrón claro cuando se examinó el número de tucanes registrado bajo muestreos de PCV (Tabla 5.18). El número de especies de tucanes era más alto en EI, mientras el menor número se registro en tipo B/F (CAPH) y en E/F (TRC). Aunque, el mayor número de individuos se registro en tipo B (ECO). Se calculó que el número de tucanes en general no era relacionado significativamente con la cantidad de árboles en fruto (r = -0.179, p = 0.539).
Tabla 5.18 El número de frugivoros grandes del dosel registrados durante muestreos de PCV. El número de especies de guacamayos grandes se refiere a Ara ararauna, A. chloroptera, A. macao.
| Albergue | No. de Visitas |
No. de Loros Observados |
No. de Especies de Guacamayos Grandes Observados |
No. de Especies de Tucanes Observados |
No. de Individuos de Tucanes Observados |
| CAPH | 5 |
216 |
6 |
4 |
86 |
| EI | 5 |
175 |
26 |
7 |
182 |
| ECO | 6 |
665 |
45 |
6 |
226 |
| SACHA | 4 |
491 |
109 |
6 |
141 |
| TRC | 4 |
519 |
161 |
4 |
79 |
Mallas de Neblina
El esfuerzo de muestreo con mallas de neblina varió entre 1,620 horas/red en TRC hasta 2,700 horas/red en CAPH por un total de 10,530 horas/red (Tabla 5.19). Esto resultó en un total de 1,808 capturas representando 126 especies (Tabla 5.20). Las tres especies más comunmente capturadas eran Pipra fascicauda (249), Myrmotherula hauxwelli (157) y Gymnopithys salvini (107), mientras 26 especies eran capturadas solo una vez. Pipra fascicauda fue la especie más comunmente capturada en CAPH, ECO y TRC, G. salvini fue la especie más frecuentemente capturada en EI, y M. hauxwelli fue la especie más frecuentemente capturada en SACHA. Algunas estimaciones de densidad para varias especies, utilizando datos de captura, son representados en Tablas 5.10 – 5.17).
Tabla 5.19. Esfuerzo de Muestreo con Redes de Neblina
| Albergue | CAPH |
EI |
ECO |
SACHA |
TRC |
| Horas/Red | 2,160 |
1,800 |
2,700 |
2,250 |
1,620 |
TablA 5.20. Número de capturas para cada una de las 126 especies registradas a través de mallas de neblina.
| FAMILIA | No. | ESPECIE | CAPH |
EI |
ECO |
SACH |
TRC |
Total |
| Tinamidae | 1 | Crypturellus bartletti | 1 |
1 |
||||
| 2 | C. soui | 1 |
1 |
|||||
| Accipitridae | 3 | Leucopternis schistacea | 2 |
2 |
||||
| Falconidae | 4 | Micrastur gilbicollis | 1 |
1 |
||||
| 5 | M. ruficollis | 1 |
1 |
|||||
| Phasianidae | 6 | Odontophorus stellatus | 2 |
2 |
||||
| Columbidae | 7 | Geotrygon montana | 2 |
1 |
3 |
1 |
1 |
8 |
| Trochilidae | 8 | Florisuga mellivora | 1 |
1 |
||||
| 9 | Glaucis hirsuta | 14 |
3 |
17 |
||||
| 10 | Phaethornis ruber | 1 |
1 |
2 |
||||
| 11 | P. hispidus | 7 |
1 |
14 |
3 |
7 |
32 |
|
| 12 | Thalurania furcata | 1 |
2 |
1 |
4 |
|||
| 13 | Threnetes leucurus | 3 |
1 |
1 |
3 |
8 |
||
| Alcedinidae | 14 | Chloroceryle aenea | 1 |
5 |
6 |
|||
| Momotidae | 15 | Electron platyrhynchun | 1 |
1 |
||||
| 16 | Momotus momota | 2 |
3 |
5 |
||||
| Bucconidae | 17 | Bucco macrodactylus | 1 |
1 |
||||
| 18 | Malacoptila semicincta | 2 |
3 |
1 |
6 |
|||
| 19 | Monasa nigrifrons | 3 |
1 |
2 |
1 |
7 |
||
| 20 | Nystalus striolatus | 1 |
1 |
|||||
| Ramphastidae | 21 | Aulaconrhynchus prasinus | 2 |
2 |
||||
| 22 | Ramphastos culminatus | 1 |
1 |
|||||
| 23 | Selenidera reinwardtii | 2 |
2 |
4 |
||||
| Picidae | 24 | Picumnus rufiventris | 2 |
2 |
||||
| 25 | Veniliornis affinis | 1 |
1 |
|||||
| Dendrocolaptidae | 26 | Campylorhamphus trochilirostris | 3 |
3 |
||||
| 27 | Deconychura longicauda | 4 |
4 |
5 |
2 |
8 |
23 |
|
| 28 | Dendrocincla fuliginosa | 2 |
2 |
6 |
2 |
3 |
15 |
|
| 29 | D. merula | 24 |
21 |
22 |
7 |
4 |
78 |
|
| 30 | D. certhia | 2 |
2 |
|||||
| 31 | D. picumnus | 2 |
2 |
|||||
| 32 | Glyphorynchus spirurus | 3 |
33 |
2 |
5 |
43 |
||
| 33 | Sittasomus griseicapillus | 1 |
1 |
|||||
| 34 | Xyphorhynchus guttatus | 2 |
1 |
7 |
12 |
22 |
||
| 35 | X. ocellatus | 8 |
3 |
9 |
4 |
24 |
||
| 36 | X. spixii | 5 |
5 |
12 |
4 |
9 |
35 |
|
| Furnariidae | 37 | Ancistrops strigilatus | 2 |
2 |
||||
| 38 | Automolus dorsalis | 1 |
1 |
2 |
||||
| 39 | A. infuscatus | 2 |
9 |
8 |
19 |
|||
| 40 | A. melanopezus | 1 |
3 |
4 |
||||
| 41 | A. ochralaemus | 4 |
10 |
2 |
5 |
21 |
||
| 42 | A. rubiginosus | 1 |
1 |
|||||
| 43 | A. rufipileatus | 1 |
1 |
2 |
||||
| 44 | Hyloctistes subulatus | 4 |
4 |
|||||
| 45 | Philydor pirrhodes | 3 |
7 |
2 |
1 |
13 |
||
| 46 | Sclerurus caudacutus | 7 |
6 |
3 |
2 |
4 |
22 |
|
| 47 | Simoxenops ucayale | 1 |
1 |
1 |
3 |
|||
| 48 | Synallaxis rutilans | 3 |
3 |
|||||
| 49 | Xenops minutus | 3 |
4 |
6 |
3 |
4 |
20 |
|
| Formicariidae | 50 | Cercomacra manu | 2 |
2 |
||||
| 51 | Chamaeza nobilis | 1 |
1 |
|||||
| 52 | Conopophaga peruviana | 8 |
9 |
8 |
4 |
29 |
||
| 53 | Dichrozoma cincta | 3 |
1 |
1 |
5 |
|||
| 54 | Formicarius analis | 11 |
2 |
6 |
6 |
8 |
33 |
|
| 55 | F. colma | 2 |
3 |
2 |
7 |
|||
| 56 | Gymnopithys salvini | 19 |
32 |
28 |
6 |
22 |
107 |
|
| 57 | Hylophylax naevia | 13 |
13 |
|||||
| 58 | H. punctulata | 2 |
2 |
|||||
| 59 | H. poecilonota | 1 |
16 |
17 |
||||
| 60 | Hypocnemis cantator | 2 |
1 |
3 |
||||
| 61 | Hypocnemoides maculicauda | 1 |
1 |
|||||
| 62 | Microrhopias quixensis | 1 |
1 |
|||||
| 63 | Myrmeciza atrothorax | 1 |
1 |
|||||
| 64 | M. goeldii | 4 |
1 |
5 |
||||
| 65 | M. hemimelaena | 5 |
9 |
9 |
23 |
|||
| 66 | M. hyperythra | 4 |
6 |
3 |
2 |
15 |
||
| 67 | Myrmoborus leucophys | 1 |
1 |
5 |
6 |
13 |
||
| 68 | M. myotherinus | 8 |
11 |
3 |
10 |
6 |
38 |
|
| 69 | Myrmotherula axillaris | 2 |
7 |
2 |
5 |
16 |
||
| 70 | M. haematonata | 7 |
10 |
1 |
18 |
|||
| 71 | M. hauxwelli | 44 |
15 |
36 |
31 |
31 |
157 |
|
| 72 | M. iheringi | 1 |
1 |
2 |
||||
| 73 | M. lecophthalma | 3 |
7 |
1 |
4 |
4 |
19 |
|
| 74 | M. longipennis | 12 |
3 |
4 |
1 |
4 |
24 |
|
| 75 | M. menetriesii | 2 |
1 |
1 |
4 |
|||
| 76 | M. ornata | 1 |
1 |
|||||
| 77 | Schirtocichla leucostigma | 2 |
2 |
|||||
| 78 | Percnostola lophotes | 2 |
1 |
3 |
||||
| 79 | Phlegopsis nigromaculata | 10 |
9 |
14 |
13 |
46 |
||
| 80 | Pygiptila stellaris | 1 |
1 |
2 |
||||
| 81 | Sclateria naevia | 1 |
1 |
|||||
| 82 | Thamnomanes ardesiacus | 10 |
4 |
11 |
3 |
9 |
37 |
|
| 83 | T. schistogynus | 2 |
1 |
3 |
4 |
8 |
18 |
|
| 84 | Thamnophilus aethiops | 8 |
2 |
10 |
||||
| 85 | T. schistaceus | 3 |
2 |
1 |
1 |
2 |
9 |
|
| Cotingidae | 86 | Lipaugus vociferans | 1 |
1 |
2 |
|||
| Pipridae | 87 | Pipra chloromeros | 13 |
6 |
7 |
1 |
5 |
29 |
| 88 | P. coronata | 1 |
8 |
9 |
||||
| 89 | P. fascicauda | 69 |
9 |
85 |
22 |
64 |
249 |
|
| 90 | P. rubrocapilla | 1 |
2 |
1 |
7 |
|||
| 91 | Machaeropterus pyrocephalus | 1 |
1 |
|||||
| 92 | Schiffornis turdinus | 9 |
9 |
|||||
| Tyrannidae | 93 | Attila bolivianus | 1 |
1 |
||||
| 94 | A. spadiceus | 2 |
2 |
|||||
| 95 | Corythopis torquata | 7 |
8 |
5 |
11 |
31 |
||
| 96 | Hemitriccus flamulatus | 3 |
3 |
|||||
| 97 | H. zosterops | 2 |
1 |
3 |
||||
| 98 | Laniocera hypopyrrha | 2 |
1 |
1 |
4 |
|||
| 99 | Leptopogon amaurocephalus | 1 |
1 |
|||||
| 100 | Mionectes macconnelli | 1 |
1 |
2 |
||||
| 101 | M. oleagineus | 3 |
4 |
1 |
2 |
1 |
11 |
|
| 102 | Mionectes olivaceus | 1 |
2 |
4 |
7 |
|||
| 103 | Onychorhynchus coronatus | 1 |
2 |
1 |
8 |
12 |
||
| 104 | Platyrinchus coronatus | 26 |
17 |
9 |
3 |
55 |
||
| 105 | P. platyrhynchos | 3 |
1 |
1 |
1 |
6 |
||
| 106 | Ramphotrigon megacephala | 2 |
2 |
|||||
| 107 | Rhynchocyclus olivaceus | 1 |
1 |
|||||
| 108 | Rhytipterna simplex | 1 |
2 |
1 |
4 |
|||
| 109 | Terenotricus erythrurus | 3 |
3 |
4 |
5 |
3 |
18 |
|
| Troglodytidae | 110 | Cyphorhinus aradus | 9 |
4 |
10 |
11 |
3 |
37 |
| 111 | Microcerculus marginatus | 4 |
1 |
3 |
2 |
10 |
||
| Muscicapidae | 112 | Catharus ustulatus | 1 |
1 |
2 |
|||
| 113 | Turdus albicollis | 6 |
1 |
1 |
3 |
6 |
17 |
|
| 114 | T. amaurochalinus | 1 |
1 |
|||||
| 115 | T. hauxwelli | 4 |
10 |
3 |
17 |
|||
| 116 | T. lawrencii | 1 |
1 |
|||||
| Vireonidae | 117 | Hylophilus ochraceiceps | 2 |
1 |
3 |
|||
| Icteridae | 118 | Psarocolius oseryi | 1 |
1 |
||||
| Parulidae | 119 | Basileuterus fulvicauda | 2 |
2 |
||||
| Emberizidae | 120 | Habia rubica | 7 |
15 |
9 |
6 |
2 |
39 |
| 121 | Lanio versicolor | 2 |
2 |
4 |
8 |
|||
| 122 | Tangara schrankii | 2 |
1 |
3 |
6 |
|||
| 123 | Arremon taciturnus | 5 |
2 |
8 |
3 |
18 |
||
| 124 | Oryzoborus angolensis | 1 |
1 |
|||||
| 125 | Saltator maximus | 2 |
2 |
|||||
| 126 | Cyanocompsa cyanoides | 2 |
1 |
4 |
7 |
|||
| Total Capturas | 404 |
318 |
447 |
233 |
406 |
1808 |
||
| Total No. de Especies | 63 |
62 |
69 |
55 |
82 |
126 |
Descripción y Estatus de Conservación de las Especies