PHYLOGENY AND BIOGEOGRAPHY OF FAGACEAE SENSO LATO
BY
CHARLES HENRY WALKER, II
I) PHYLOGENY
Fagaceae senso lato includes the family Fagaceae and the genus Nothofagus. Fagaceae senso stricto and Nothofagus were for a long time united because
their flowers/fruits were contained within cupules. There is wide variation in cupule and fruit morphology, as well as pollination syndrome (wind vs. generalistic
insect), within Fagaceae senso lato. Differences in flower, fruit, and pollen structure have been cited for placing Nothofagus into a separate family, the
Nothofagaceae (Nixon, 1982). Betulaceae and Juglandaceae have similar inflorescence morphology and are closely related (Figure 1).
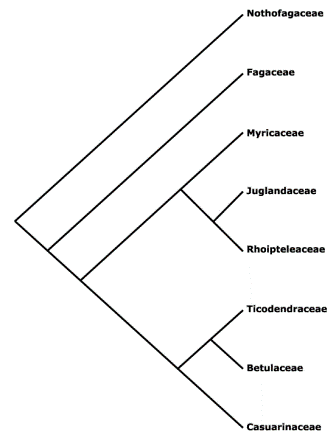
Figure 1.Cladogram of Fagaceae and other closely related families (Missouri Botanical Gardens, 2002).
In addition Hammamelidaceae and Salicaceae have pollen that is similar in shape, size, etc. with that of Fagaceae (Crepet & Daghlian, 1980). Fagaceae has
been classified within the Rosids in the Order Fagales along with Betulaceae, Casuarinaceae, Juglandaceae, Myricaceae, Nothofagaceae, Rhoipteleaceae, and
Ticodendraceae (Figure 2).
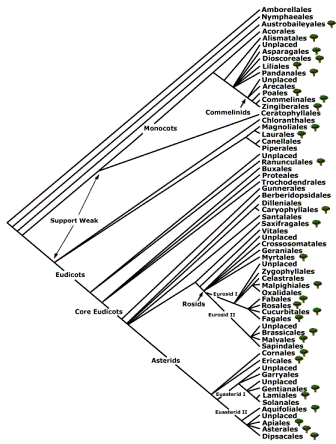
Figure 2. Relationship of Fagales to other orders (Bremer, et al., 1998).
The distribution of Fagaceae senso stricto is predominantly in the Northern Hemisphere and presently it has 7 genera with about 1000 species (Manos, et al.,
2001). The center of diversity for Fagaceae senso lato extends from Yunnan, China to Queensland, Australia and includes Castanopsis, Fagus, Lithocarpus,
Nothofagus, Quercus, and Trigonobalanus (Schuster, 1972). The number of species included in Fagaceae senso lato is in flux, e.g., new species are being
discovered: Quercus ichnusae, an endemic species of Sardinia (Mossa, et al., 1999). In some classifications Fagaceae is subdivided into 4 subfamilies, the
Castaneoideae, Fagoideae, Quercoideae, and Trigonobalanoideae (Figure 3).
GENERA.................................................................................................................SUBFAMILY
1. Castanea(10)
2. Castanopsis (99)(Pasaniopsis)................................................................................A)Castaneoideae
3. Chrysolepis (2)
4. Lithocarpus (300) (Pasania)5. Fagus (10) .....................................................................................................................B) Fagoideae
6. Quercus (450) (Cyclobalanopsis)..............................................................................C)Quercoideae
7. Trigonobalanus (3) (Colombobalanus) (Formanodendron)................................D) Trigonobalanoideae
Figure 3. Subfamilies and genera of Fagaceae sensu stricto (Bremer, et al., 1998). The number of species within each genus is enclosed within parentheses.
A) Subfamily Castaneoideae
Castanea is distributed throughout the temperate areas of the Northern Hemisphere with its greatest diversity in eastern Asia. It is also found in the United
States, Europe, and Japan. It has six long styles per pistil, and fruits that are essentially enclosed by the cupules. (Crepet & Nixon, 1989a). Castanopsis is
found in North America (2) and tropical and subtropical Asia (99). There are three fruits per cupule in this genus (Crepet & Nixon, 1989a). There are two
species of Chrysolepis and both of these are found only in the western United States. These two species have relatively long styles with fruits that are enclosed
by the cupules (Crepet & Nixon, 1989a). One species of Lithocarpus is found in the United States, and it is found throughout much of Southeast Asia with 2
centers of diversity in Indochina and Borneo. There is only one fruit per cupule in this genus (Crepet & Nixon, 1989a).
B) Subfamily Fagoideae
Fagus includes the American beech found in eastern North America southward to Mexico along with 9 other species in Eurasia and Taiwan (Schuster, 1972).
C) Subfamily Quercoideae
The genus Quercus is distributed throughout temperate and tropical areas of the Northern Hemisphere with a center of diversity in America (Daghlian &
Crepet, 1983). Quercus petraea and Q. robur are found further north than any other species of oak, reaching 60º in the Baltic region (Axelrod, 1983).
Rubrae (Erythrobalanus) includes the red and black oaks. The fruit in this group matures at the end of the second year, stigmas are elongate, and leaves are
bristle-tipped (Daghlian & Crepet, 1983).
Protobalanus represents fossil species that are intermediate between Leucobalanus and Erythrobalanus. Leucobalanus (Lepidobalanus) includes the white oaks.
Their fruit matures at the end of the first year, stigmas are sessile, and leaves are not bristle-tipped (Daghlian & Crepet, 1983). Cyclobalanopsis is now
considered to be Quercus and is distributed across eastern Asia and Malaysia.
D) Subfamily Trigonobalanoideae
Trigonobalanus is found in north Borneo, the Celebes, north Thailand, Malaya, and Sarawak (Heywood, 1978). Colombobalanus is now considered to be a
species of Trigonobalanus and is native to Colombia, South America (Crepet & Nixon, 1989a). Likewise Formanodendron is a species of Trigonobalanus
that is native to Thailand and nearby China (Crepet & Nixon, 1989b).
E) NOTHOFAGACEAE
Nothofagus is the only genus in this family and include 35-40 species that are found from South America to Australasia with none crossing Wallace’s Line
(Schuster, 1972). This genus has a wide ecological tolerance, growing in tropical to cold temperate climates (Van Steenis, 1971).
II BIOGEOGRAPHY (See figure 4 for order of Epochs)
SENONIAN EPOCH
Globally the climate was warmer than it is today with both Poles being ice-free and having cool temperate climates. The United States, southern Canada, and
eastern Asia had warm temperate climates, while Europe had a paratropical climate (Scotese, 2001). Castanea and Nothfagus are believed to have originated
prior to the Santonian Age of this epoch (Daghlian & Crepet, 1983). It has been hypothesized that the evolutionary center of origin of Nothofagus was
southern South America. Using Bremer’s (1992) and Hausdorf’s (1998) methods, Swenson, et al. (2000) identified southern South America as the likely
ancestral area and this is supported by the current fossil record. The dispersal of Nothofagus has been tied to the land as the nuts can not tolerate seawater,
will sink in water, and can only be transported 2-3 km by wind (Van Steenis, 1971). From South America, Nothofagus migrated to Antarctica and then to
Australasia in what has been called the "marsupial route" (Schuster, 1972). Migration occurred while South America, Australia, New Zealand, Antarctica, etc.
were still connected with one another as Gondwana, but each of these areas was part of a different tectonic plate that was traveling in a different direction
and at a different rate of speed. The opening of the South Atlantic, due to sea-floor spreading, began 125-130 Ma during the late Hauterivian or early
Barremian Ages and resulted in South America moving away from Africa, Antarctica, and Australia (Raven and Axelrod, 1974). Campanian Age fossils from
Georgia, U.S.A share morphological features with both Nothofagus and Castaneoids (Chrysolepis) of Fagaceae, but they also have unique characteristics,
e.g., very small pollen size with reticulate to microfoveolate exine ornamentation (Herendeen, et al., 1995). The separation of New Zealand and New
Caledonia from New Zealand occurred during this Age (Raven & Axelrod, 1974). The nervation of the Neogene species Quercus nevadensis has shown that
it is closely allied to the Asian species Lithocarpus densiflorus (Axelrod, 1983). A probable connection would have been from east Asia to Europe and
across a much narrower Atlantic Ocean during late Cretaceous times (Axelrod, 1983). Axelrod, (1983) also cites Lithocarpus densiflorus, (California and
Oregon), Chrysolepis chrysophylla (California, Oregon, and Washington), and C. sempervirens (California, Nevada, and Oregon) as being allied to Asian
taxa and thus another Cretaceous link.
SENONIAN-PALEOCENE BOUNDARY
Based on fossil evidence, Castaneoideae and Fagoideae are believed to have diverged from one another some time in the early Tertiary or late Cretaceous
(Crepet & Nixon, 1989b). Fagaceae originated in the montane tropics, migrating across the tropics via the Atlantic into temperate areas (Axelrod, 1983).
PALEOCENE-EOCENE BOUNDARY
Castanopsoidea columbiana fossils from western Tennessee most closely resemble Asian members of Castanopsis (Crepet & Nixon, 1989a). Crepet &
Nixon (1989a) also found fossils of Trigonobalanoidea americana from the same deltaic location that is similar to extant species of Trigonobalanus from
Colombia, South America, and Thailand-China. The climate at this site during the late Paleocene was subtropical which is similar to the current climate of
modern species of Castaneoideae, but dissimilar to the wet montane tropical habitats of extant trigonobalanoid species. There are no extant species of
Trigonobalanus in North America, but there are in Colombia. The discovery of Trigonobalanoidea fossils in North America supports the hypothesis that
trigonobalanoids were once more widely distributed in the Northern Hemisphere and dispersed to South America in the Late Tertiary (Nixon & Crepet, 1989).
EOCENE EPOCH
During the early Eocene the North Pole had a cool temperate climate with the United States and Eurasia with a paratropical climate (University of Rochester,
(2001). The earliest fossils of Quercus, including wood, leaves, and fruits, are known from America and England (Daghlian & Crepet, 1983). Also all of the
modern subfamilies and some extant genera, e.g., Rubrae and Leucobalanus, were recognizable during this epoch (Daghlian & Crepet, 1981). This may be due
to the increased aridity that developed over southwestern Asia and North America during the middle Eocene (Axelrod, 1983) and has persisted to this day.
Fossil Castaneoideae with robust inflorescences from mid-Eocene deposits in Tennessee show that they were essentially like those of modern, insect-pollinated
Castaneoideae and may have been insect-pollinated at well (Crepet & Daghlian, 1980). Nuclear ribosomal DNA sequences of 179 accessions were studied
and results suggest that wind pollination evolved at least 3 times and hypogeal cotyledons once within Fagaceae (Manos, et al., 2001).
EOCENE-OLIGOCENE EPOCHS
Globally the climate was warmer than it is today with ice just beginning to form in Antarctica, tropical rainforests covered India, and warm temperate forests
encompassed most of Australia (Scotese, 2001). The transitionary period between these 2 epochs has been characterized with having had rapid global cooling,
drying and a loss of equibility (Daghlian & Crepet, 1983). It was probably through hybridization that Quercus underwent an adaptive radiation as they
migrated into new habitats that resulted from the change in climate (Daghlian & Crepet, 1983). In addition to cooler and drier conditions, terrain diversified
rapidly in Mexico-Central America, the southwestern United States, the Mediterranean basin, and southeastern Asia (Axelrod, 1983).
OLIGOCENE EPOCH
Eocene sites in Texas were dominated by members of Juglandaceae, but by the middle to late Oligocene Fagoideae had come to dominate (Crepet & Nixon,
1989b). Fossils found at the Texas site were pretrigonobalanoid and could be interpreted as a sister group to either Fageae or Querceae (Crepet & Nixon,
1989b). The climate during this epoch was subtropical to warm temperate with a humid to subhumid moisture regime (Galloway, 1977). Ice covered
Antarctica, but not the North Pole and warm temperate forests covered North America and northern Eurasia (Scotese, 2001). The fossil species Nothofagus
smithtonensis lived during the early Oligocene Epoch in northwest Tasmania, Australia and its reduced and less woody cupule contained only one fruit as do
extant species (Hill, 1994). By the end of this epoch Quercus was well represented in the flora of Colorado (MacGinitie, 1953).
OLIGOCENE-MIOCENE EPOCHS
Quercus is well represented in the flora of Europe during this epoch. Light or day length may be the principal factor that has limited Quercus to latitudes less
than 40º N (Axelrod, 1983). Five different fossil species of Nothofagus, representing 3 of the 4 extant subgenera were present in northwest Tasmania during
this time period (Hill, 2001). The subgenus Lophozonia still occurs in Tasmania, but subgenus Nothofagus is restricted to South America and subgenus
Brassospora to New Guinea and New Caledonia. Hill (2001) attributes the disjunct distribution patterns to extinct climates.
MIOCENE EPOCH
During this epoch Quercus had become a common component of the flora of Japan (Wateri, 1941). The climate during this epoch was somewhat warmer that
it is today. The United Kingdom and northern Europe had a tropical climate and Australia was less arid than today (Scotese, 2001). After increasing in diversity
during the Oligocene, many of the modern species of Quercus were present by the Miocene (Axelrod, 1983). Bi-directional floristic exchange between Asia
and North America occurred via the Bering Land Bridge until intercontinental disjunction during the Serravallian-Tortonian epochs (Axelrod, 1983). The rise of
the Cascade Mountains in Oregon and Washington provided xeric, well-drained sites in which Quercus and other species from subtropical and warm
temperate areas could inhabit (Axelrod, 1983). The fossil species/extant species Quercus shrevoides/Q. shrevei was present in west-central Nevada prior to
the Burdigalian Age of the Miocene Epoch, but because of the rise of the Sierra Mountains dryer conditions prevailed eastward and it shifted from Nevada into
the lower Sierra Nevadas of California. Again, during the Zanclian and Messinian Ages (late Miocene-early Pliocene), Quercus shrevoides shifted westward
to the outer south Coast Ranges of California. Quercus wislizenoides/Q. wislizenii replaced Q. shrevoides as the later migrated westward (Axelrod, 1983).
In addition to responding to increased topographic, climatic, and edaphic diversity, Quercus and other taxa may have undergone rapid speciation when climatic
extremes sharply increased, e.g. 35, 13, and 5 MA (Axelrod, 1983).
PLIOCENE EPOCH
After the northeastward movement of Australasia in the Tertiary Period, there was a large migration of species from Asia (Laurasian origin) into Australasia as
early as the Miocene Epoch(Schuster, 1972).
PLEISTOCENE EPOCH
Quercus has a Holoarctic distribution except for one species that is found in Colombia. It is believed that there was a North America-South America land
connection during the Pleistocene Epoch and to substantiate this, there is a large and sudden occurrence of pollen in the fossil record for that time period
(Crepet & Nixon, 1989a). Fagus is believed to have originated in Asia and migrated bidirectionally to Europe and North America via the Bering Land Bridge
(Manos & Stanford, 2001). Morphological features of extant species of Fagus exhibit a west-east gradient that is the result of area disruptions during the
Pleistocene coupled with post-glacial recolonization of Europe (Denk, et al., 2002). Fossil leaves from Nothofagus beardmorensis were located in the
Transantarctic Mountains of Antarctica and are from the late Pliocene, indicating that the climate was substantially different than it is today (Hill, et al., 1996).
Utilizing growth forms and tree rings in the fossil wood, Francis et al. (1996) concluded that N. beardmorensis was a prostrate, deciduous dwarf tree that
grew where mean annual temperature was about -12° C and summers were short. Sea-floor spreading, the subduction of tectonic plates, the uplifting of
mountains, the emergence of new archipelagos and the closing in of old ones have all changed the physical environment in which plants grow. The expansion or
the contraction of individual species of Fagaceae senso lato has been governed by environmental, geographical, and ecological factors. Each of the individual
species through differing dispersal rates has taken advantage of their environment and those species with similar ecological amplitudes have spread together in
homologous patterns (Van Steenis, 1971).
LITERATURE CITED
Axelrod, D. I. 1983. Biogeography of oaks in the Arcto-Tertiary province. Annals of the Missouri Botanical Garden 70: 629-657.
Bremer, K, Bremer, B, and Thulin, M. 1998. Classification of flowering plants. (http://www.systbot.uu.se/classification/classification98.html)
Crepet, W. L. and Daghlian, C. P. 1980. Castaneoid inflorescences from the middle Eocene of Tennessee and the diagnostic value of pollen (at the subfamily level) in the Fagaceae. American Journal of Botany 67: 739-757.
Crepet, W. L. and Nixon, K. C. 1989a. Earliest megafossil evidence of Fagaceae: phylogenetic and biogeographic implications. American Journal of Botany 76: 842-855.
Crepet, W. L. and Nixon, K. C. 1989b. Extinct transitional Fagaceae from the Oligocene and their phylogenetic implications. American Journal of Botany 76: 1493-1505
Daghlian, C. P. and Crepet, W. L. 1983. Oak catkins, leaves and fruits from the Oligocene Catahoula formation and their evolutionary significance. American Journal of Botany 70: 639-649.
Denk, T., Grimm, G., Stogerer, K., Langer, M., and Hemleben, V. 2002. The evolutionary history of Fagus in western Eurasia: evidence from genes, morphology and the fossil record. Plant Systematics and Evolution 232: 213-236.
Francis, J. E. and Hill, R. S. 1996. Fossil plants from the Pliocene Sirius Group, Transantarctic Mountains: evidence for climate from growth rings and fossil leaves. Palaios 11: 389-396.
Herendeen, P. S., Crane, P. R., and Drinnan, A. N. 1995. Fagaceous flowers, fruits, and cupules from the Campanian (late Cretaceous) of central Georgia, USA. International Journal of Plant Sciences 156: 93-116.
Heywood, V. H. 1978. Flowering plants of the world. Oxford University Press. London.
Hill, R. S. 1994. Nothofagus smithtonensis (Nothofagaceae), a new macrofossil species from Oligocene sediments in northwest Tasmania, Australia, and its phylogenetic significance. Review of Palaeobotany and Palynology 80: 115-121.
Hill, R. S., Harwood, D. M., and Webb, P. N. 1996. Nothofagus beardmorensis (Nothofagaceae), a new species based on leaves from the Pliocene Sirius Group, Transantarctic Mountains, Antarctica. Review of Palaeobotany and Palynology 94: 11-24.
Hill, R. S. 2002. Nothofagus cupules from Oligocene-early Miocene sediments at Balfour, Northwest Tasmania, Australia. International Journal of Plant Sciences 162: 683-690.
Manos, P. S., Zhou, Z. K., and Cannon, C. H. 2001. Systematics of Fagaceae: phylogenetic tests of reproductive trait evolution. International Journal of Plant Sciences 162: 1361-1379.
Manos, P. S. and Stanford, A. M. 2002. The historical biogeography of Fagaceae: Tracking the Tertiary history of temperate and subtropical forests of the Northern Hemisphere. International Journal of Plant Sciences 162: S77-S93.
Missouri Botanical Gardens. 2002. Angiosperm phylogeny. (http://www.mobot.org/MOBOT/Research/APweb/treeapweb2map.html)
Mossa L., Bacchetta, G., and Brullo, S. 1999. Quercus ichnusae (Fagaceae), a new species from Sardinia. Israel Journal of Plant Sciences 47: 199-207.
Nixon, K. C. 1982. In support of recognition of Nothofacaceae Kuprianova. Botanical Society of America Publication 162: 102.
Nixon, K. C. and Crepet, W. L. 1989. Trigonobalanus (Fagaceae): taxonomic status and phylogenetic relationships. American Journal of Botany 76: 828-841.
Poling, Jeff. 1997. Geologic Ages of Earth History. (http://www.dinosauria.com/dml/history.htm)
Raven, P. H. and Axelrod, D. I. 1974. Angiosperm biogeography and past continental movements. Annals of the Missouri Botanical Garden 61: 539-673.
Schuster, R. M. 1972. Continental movements, 'Wallace's line' and Indomalayan-Australasian dispersal of land plants: some eclectic concepts. The Botanical Review 38: 3-86.
Scotese, C. R. 2001. Paleomap Project. (http://www.scotese.com/climate.htm.)
Swenson, U., Hill, R. S., and McLoughlin, S. 2000. Ancestral area analysis of Nothofagus (Nothofagaceae) and its congruence with the fossil record. Australian Systematic Botany 13: 469-478.
University of Rochester. Life in the Eocene epoch. 2002. (http://www.earth.rochester.edu/ees201/Laidlaw/laidlawe2.html)
Van Steenis, C. G. G. J. 1971. Nothofagus, key genus of plant geography, in time and space, living and fossil, ecology and phylogeny. Blumea 19: 65-98.
Wateri, S. 1941. Studies on the fossil woods from the Tertiary of Japan. I. Fossil woods from the River Mabeti, Anati village, Ninohe district, Iwate Prefecture. Japanese Journal of Botany 11: 385-438.
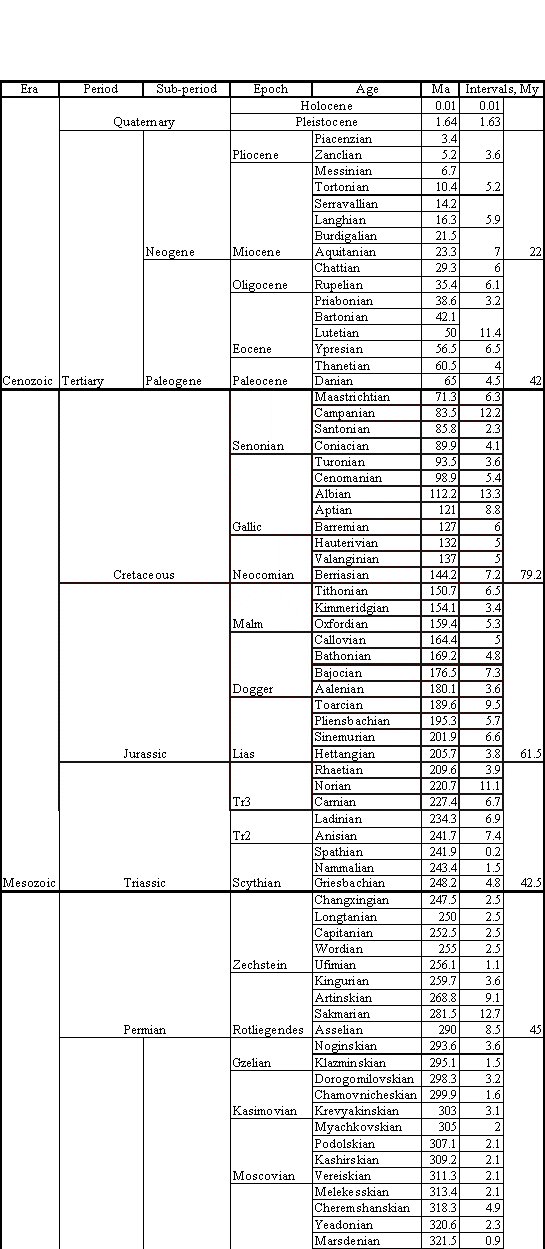
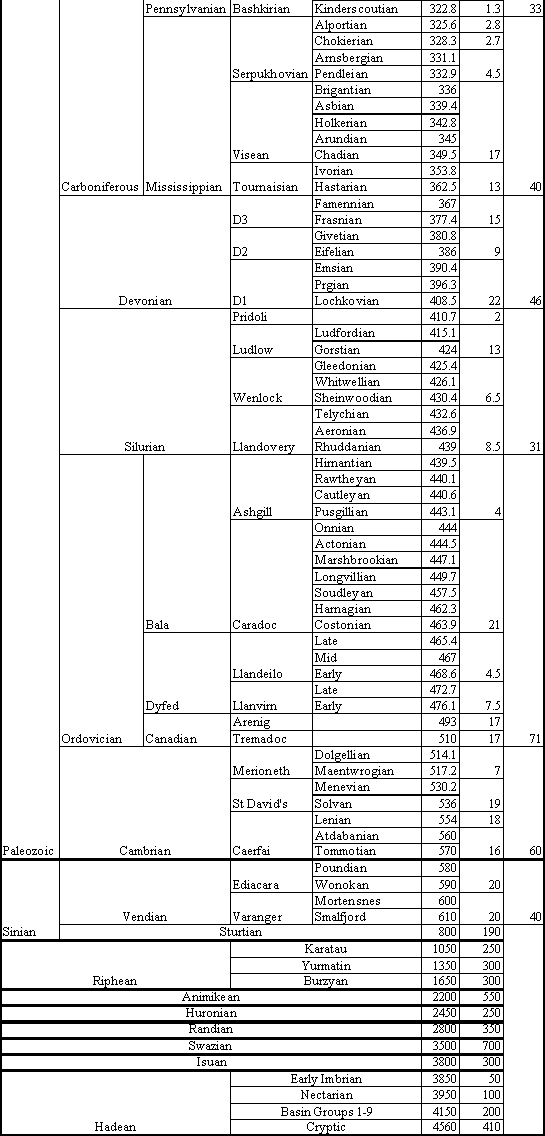
Figure 4. Geologic Ages of Earth History (Poling, 1997). Table is read bottom to top: lower boundaries for Ages are given. Ma = million (years) ago; how long ago the age began. My = million years; how long the age or period lasted.