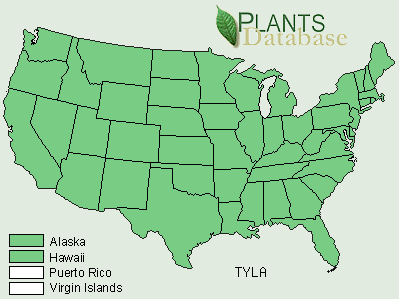
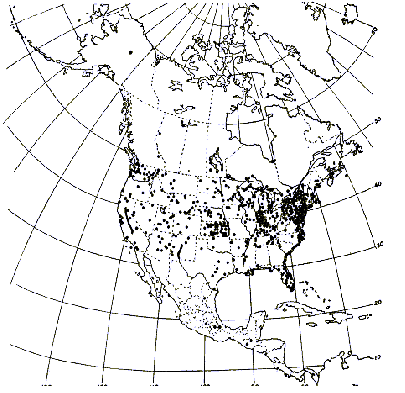
Typha domingensis has been reported from the 25 states in the southern half of the United States (Figures 3 and 4) and is restricted to southern latitudes.
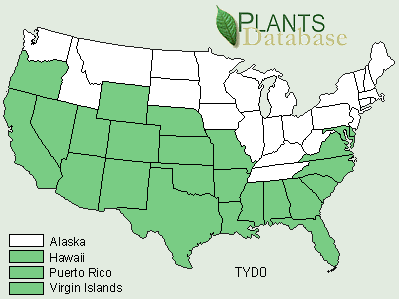
FIGURE 3. Distribution of Typha domingensis Pers., the southern cattail (USDA, NRCS, 2001).
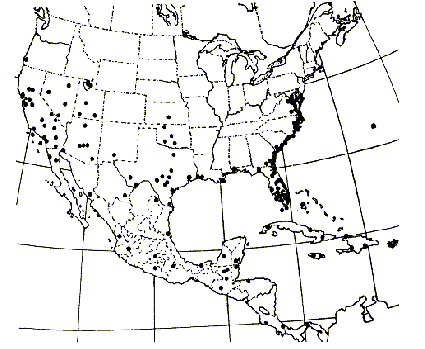
FIGURE 4. Distribution of Typha domingensis Pers., the southern cattail (Hotchkiss and Dozier 1949).
Typha angustifolia has been spreading across North America since the early 1800s via canals, railroads and highways and is considered by some to be an introduced European species (Stuckey 1987). At its northern range in central Canada, winter temperatures range from 14° to -13°C (Smith 1967). Some consider both T. angustifolia and T. domingensis to be subject to K-selection as they are restricted to more of a southern distribution than is T. latifolia (McNaughton 1975). Others consider T. angustifolia and T. domingensis to be specialized for climate, whereas T. latifolia is specialized for competitive ability in freshwater sites (McNaughton 1979).Typha x glauca and T. angustifolia have been reported from 31 and 40 different states respectively (Figures 5, 6, and 7).
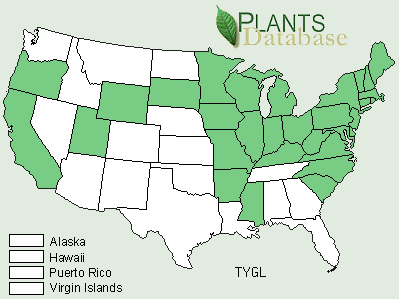
FIGURE 5. Distribution of Typha x glauca Godr. (pro sp.) [T. angustifolia or T. domingensis x T. latifolia] (USDA, NRCS, 2001).
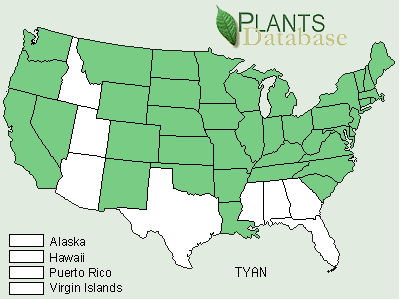
FIGURE 6. Distribution of Typha angustifolia L., the narrowleaf cattail (USDA, NRCS, 2001).
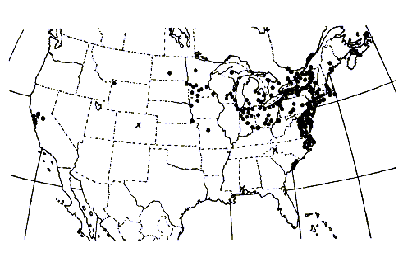
FIGURE 7. Distribution of Typha angustifolia L., the narrowleaf cattail (Hotchkiss and Dozier 1949).
Some authors believe that the major threat to T. latifolia from T. x glauca is vegetative displacement and not introgression (Kuehn, Minor, and White 1999). The invasiveness of T. x glauca can be explained by the introgression/hybrid speciation hypothesis where a new taxon and one already existing in an area produce a hybrid that can grow under conditions that is not favorable to either parent (Galatowitsch et al. 1999). Typha x glauca is best adapted to conditions where the water level fluctuates rapidly (Smith 1967). Typha x glauca has supplanted T. latifolia as a result of drawdown operations on the Agassiz National Wildlife Refuge in Minnesota (Harris and Marshall 1963). At this Refuge T. latifolia was found to be intolerant of reflooding, while the number and sites of T. x glauca growing in 60 cm of water remained unchanged through five years of flooding. Typha x glauca has been shown to maintain high stand biomass from 0-100 cm of water depth (Waters and Shay 1992).
Typha domingensis is the most tolerant of salinity, followed by T. angustifolia, T. latifolia, and T. x glauca (Hotchkiss and Dozier 1949). McMillan (1959) obtained different results finding T. x glauca to be more vigorous and taller than T. latifolia, but exhibiting greater curling of the leaves than T. angustifolia in salinity experiments. Germination rates for T. latifolia are not affected by extreme ranges of pH found in marsh water (Rivard and Woodard 1989). Stands of T. angustifolia, T. x glauca, and T. latifolia have been observed growing in soils with pHs ranging from 4.1 to 8.1 (Segadas-Vianna 1951).
Physiological and morphological characteristics of Typha
Typha is a clonal dominant that is in the matrix functional group or guild that vegetatively covers entire gaps in the landscape (McJannet, et al. 1995). In these emergent perennial aquatic plants, the ramet is the unit of vegetative growth and consists of the submerged rhizome, associated roots, and the shoot (Dickerman and Wetzel 1985). Net productivity is determined by the leaf production rate and not by the photosynthetic rate (McNaughton 1974). The main meristem is basal and produces the leaves and inflorescences and it usually dies after flowering (Grace and Harrison 1986). The whole ramet dies after flowering in T. latifolia (Grace and Wetzel 1981a), but in T. angustifolia flowering consumes only the bud with rhizomes producing leaves and inflorescences the following spring (Grace and Wetzel 1982). Ecotypes native to areas with short growing seasons translocate photosynthates to their rhizomes under 16-hr/30° - 15° conditions while those from areas with long growing seasons did not (McNaughton 1974). McNaughton (1974) suggests that canopy differentiation has a definite photoperiodic element, indicating that it is under hormonal control. Growth of Typha in the spring is rapid with deep-water shoots sprouting before those found in shallow water, but growth can be curtailed by frost (Grace and Harrison 1986).
Typha latifolia has large amounts of aerenchymatic tissue and intracellular air spaces (Bieboer 1984) that enable it to survive in anaerobic conditions. Leaf stomates are at least partially open both day and night insuring that there is a constant supply of oxygen available for diffusion (Sale and Wetzel 1983). Sometimes Typha is not rooted in the mineral soil, but floats as a fibrous mat due to trapped gases and the lower specific gravity of rhizomes as compared to water (Krusi and Wein 1988). Floating leaves develop before erect leaves in both T. angustifolia and T. latifolia when they germinated underwater (Grace 1985).
Germination
All four species of Typha are classified as obligate wetland species (Reed 1988). High temperature (35° C), low O2 concentration, and long light exposure are the conditions necessary for germination, and are known to exist on bare mud flats where Typha seedlings grow in nature (Bonnewell et al. 1983). Studies in the Everglades have shown that the whole seeds of Typha make up only 5.2% of the total number of whole seeds and only 2.4% of the total number of intact seeds in the seedbank (van der Valk and Rosburg 1997).
Failure of seed germination in field trials with the seed from the same cattail spike germinating under greenhouse conditions might indicate the effect of inhibitors that are produced by adult cattail plants (Bedish 1967). High amounts of water, at least in the spring, are necessary for the establishment of cattail stands (Segadas-Vianna 1951). Typha x glauca plants growing in 2.54 cm of water have a faster growth rate and a greater amount of growth than plants growing in dry, saturated or under 15.24 cm of water (Bedish 1967). Germination of T. domingensis and T. latifolia has been shown to start within three days, and to peak after seven days with a germination rate of 22-40% (Stewart et al. 1997). Stewart et al. (1997) also found that the seed germination rate of T. latifolia was equal to or greater than that of T. domingensis at low, medium, and high nutrient levels. His germination experiments showed that T. domingensis had its lowest germination rate under conditions with zero nutrients, while for T. latifolia it was with high nutrients.
Typha latifolia can allelochemically inhibit germination of other species, as well as itself, through green leaf extracts exuded into the water of marshes (Rivard and Woodard 1989). In another experiment soil extracts and leaf, rhizome, and root extracts from T. latifolia showed no inhibitory effects on the germination and growth of lettuce, radish, cucumber, and tomato (Bonasera et al. 1979). Rivard and Woodard (1989) found that inhibition could be countered with controlled burning that increases light and deposits ash on marsh seedbeds. The lack of litter in a T. angustifolia dominated brackish tidal marsh has been shown to increase sediment temperatures by ≈3°C with an increase in mineralization of organic matter below the surface of the sediment (Jordan et al. 1989).
Seed production
Typha produces prodigious amounts of powdery pollen and is wind-pollinated (Smith 1967). Typha latifolia produces pollen that is in the form of tetrads, T. angustifolia in the form of monads, and T. x glauca in the form of monads, dyads, triads, and tetrads (Smith 1967). In Typha flowering is determined by photoperiod, occurring immediately after the formation of floral initials, and ramets must be of a minimum size and/or weight (Grace and Wetzel 1982).
One-seeded fruits with numerous gynophore hairs are produced in the second year following germination, and are dispersed by wind (Smith 1967). The fruit is classified as a follicle as there is a single lengthwise suture (Stewart et al. 1997). When the fruit comes into contact with water the pericarp opens up along the suture, releasing the seed that sinks with the pointed, posterior portion aimed downward (Grace and Harrison 1986). Typha seeds are 1-1.5 mm long and are readily buried during sedimentation and sediment mixing (Grace 1984). Under optimum conditions the seeds of T. domingensis and T. latifolia have a germination rate between 50-100% (Grace 1985).
In an open marsh biotype of T. latifolia, nonflowering ramets allocated 10-15% more to vegetative reproduction than did flowering ramets (Grace and Wetzel 1981a). Grace and Wetzel (1982) found that allocation to sexual structures in Typha decreased with increasing water depth and was related to conserving biomass in deep water. The inflorescence of T. latifolia has a fixed dry mass of 30 g (Grace and Wetzel 1981a) and usually remains standing for up to two years in wetlands before collapsing (Mason and Bryant 1975). Grace and Wetzel (1981a) found that rhizome size is related to flowering since most of the biomass for inflorescence production is stored in the rhizome. They also concluded that ramets that make a bigger allocation for root biomass instead of rhizomes would have a greater potential for nutrient uptake, since root biomass correlates roughly with root surface area. In a nutrient poor environment one would expect to find a larger allocation to roots rather than to rhizomes.
Smith (1967) found that seed production in the T. latifolia x T. domingensis and T. latifolia x T. angustifolia hybrids was 0-25% of that of its parents. Morphologically hybrids are almost always intermediate between the parent species.
Tubificid worms and litter
Tubificid worms with their mixing of sediments have significant effects on germination, survival, and height of T. domingensis (Grace 1984). Large-mesh bags containing the litter of T. angustifolia were found to contain 0-260 members of the Tubificidae per square meter in a Norfolk, U.K. study (Mason and Bryant 1975). Typha litter in small-mesh bags took longer to decompose than in large-mesh bags (Figure 8). Most of the P, Ca, K, and Mg leached out of the bags in the first month, but the N level increased due to microbial activity (Mason and Bryant 1975).
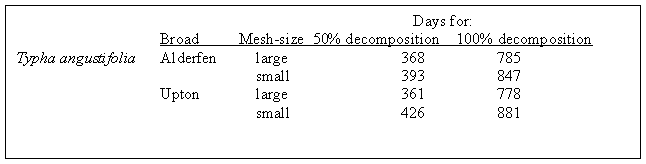
FIGURE 8. The estimated times for 50% and 100% decomposition of Typha angustifolia litter (Mason and Bryant 1975).
Competition studies
1. Typha angustifolia and T. latifolia
T. angustifolia is usually found in deeper water and T. latifolia in shallow water where the two species occur together (Grace and Wetzel 1981b). Grace and Wetzel (1981b) in their interaction experiments found T. latifolia to be a stronger competitor than T. angustifolia. This was attributed to the ability of T. latifolia to clone rapidly, with a larger allocation of resources for leaves and a higher leaf surface area than T. angustifolia. Almost 20% of the biomass of T. angustifolia was allocated to sexual structures, while in T. latifolia this allocation was less than 5 % (Grace and Wetzel 1982). The leaves of T. latifolia increase in height with increasing water depths, but were 23.1% lower than those of T. angustifolia (Grace and Wetzel 1982). This comparison was made in the transition zone between the two species in a water depth of 50 cm.
Typha x glauca plants exhibit the full range of leaf widths observed in the parents (Kuehn et al 1999). Except for spike length, which is the same as T. angustifolia, measurements for spike width, spike interval, and stigma width in T. x glauca are intermediate to the parental species (Kuehn and White 1999). There is considerable overlap in the range of values for these five characters, making accurate identification possible for only extreme specimens of T. angustifolia and T. latifolia (Kuehn and White 1999).
Typha latifolia produces almost twice as many lateral rhizomes per parent ramet as does T. angustifolia, but these are smaller and usually die the following year (Grace and Wetzel 1982). Lateral rhizomes of T. angustifolia are larger and they can regrow for several years. Large rhizomes and tall leaves have enabled T. angustifolia to grow in deeper water and to escape competition with T. latifolia.
2. Typha latifolia and Typha domingensis
Typha domingensis outcompetes T. latifolia when they are started together from seed, especially at higher densities and deeper water (Grace 1987). Typha domingensis has a higher percent germination rate than T. latifolia (Grace 1985). When planted in monocultures at the same time, T. domingensis outyields T. latifolia by more than five-fold (Grace 1987). Grace (1987) also found that T. latifolia has triple the yield of T. domingensis if planted a week earlier. Further he showed that when planted in mixtures, the relative yield of T. domingensis was negatively affected by T. latifolia, dropping to 38 % of its yield in monoculture. The growth of T. latifolia was largely unaffected by T. domingensis. In an earlier experiment the biomass of T. domingensis was reduced by 73.6%, while the biomass of T. latifolia was reduced by 30.7% when they were both grown together (Grace 1985). The competitive superiority of T. latifolia over T. domingensis may be due to its greater capacity for obtaining N (Grace 1988).
In water depth experiments T. latifolia was found to flower only five cm above the water, while T. domingensis flowered when growing at depths of five, 22, 42, 58,70, 83, 95, 105, 110, and 115 cm below the surface of the water (Grace 1989). Typha domingensis may have the ability to supply more oxygen to its rhizosphere than does T. latifolia (Grace 1988). Grace (1989) found that eventually T. latifolia died out when growing in depths greater than 95 cm. A comparison of realized distributions for both juvenile and adult distributions has shown a distinct segregation with T. latifolia dominant above the waterline and T. domingensis dominant below the waterline (Grace 1985). Grace (1985) found the competitive ability of T. domingensis declining in relation to T. latifolia as they approached maturity and he formed three hypotheses for this decline.
1. In the Flowering Effect the allocation of resources for flowering is greater for T. domingensis than it is for T. latifolia and this reduces its competitive ability.
2. With the Growth and Development effect morphological and physiological changes lead to changes in relative competitive ability.
3. In the Limitation Change Effect light or some other factor becomes progressively more limiting and changes the mechanism of competition.
At Par Pond of the Savannah River Nuclear Power Plant T. latifolia has been shown to grow where water temperatures reach 35°C during the summer, while T. domingensis disappears from the shoreline community when water exceeds 30°C (Liu et al. 1978). Liu et al. (1978) found that even though the electrophoretic isozyme patterns of the enzyme malate dehydrogenase (MDH) were identical between T. latifolia and T. domingensis, the biochemical properties of their MDH were different. All six of the isozymes of T. domingensis decayed at 50°C, while with T. latifolia only three of its six isozymes decayed when incubated at this temperature. What connection there is between the three heat stable isozymes in the MDH of T. latifolia and its ability to survive in water that is 35°C is uncertain.
Nutrients and Typha
On islands in the Atchafalaya Delta in Louisiana, as well as in other undisturbed locales, T. domingensis occurs in relatively small but dense stands (Shaffer et al. 1992). Shaffer et al (1992) have found that it is only through local erosion or accretion (≈ 4 cm sediment/yr.) that T. domingensis shifts in elevation and the area that it covers has remained constant. T. domingensis, and to a lesser extent T. latifolia, is expanding its range in the Florida Everglades due to an environment that is changing from low to high nutrient conditions (Davis 1991). The areas of soil phosphorus enrichment in a portion of the Everglades west of Fort Lauderdale closely correspond with areas of cattail encroachment (DeBusk et al 1994). The accumulation of P in the Everglades is biologically mediated with about 75% associated with organic matter (Koch and Reddy 1992). Typha domingensis has been shown to accumulate more P per square meter than either Cladium jamaicense or Eleocharis interstincta (Newman et al. 1996). Strong correlations have been made between dilute acid soluble P in hydrosoils and dissolved P in water with the shoot standing crop of T. latifolia in six different physiographic and geographic locations along the Gulf Coast (Boyd and Hess 1969). Other elements were not correlated with shoot standing crop in this study. Species like Cladium jamaicense have a low nutrient threshold and they lose their competitive capability when crossed. T. domingensis had greater growth than either Cladium jamaicense or Eleocharis interstincta when grown with higher levels of phosphorus and nitrogen (Newman et al. 1996).
Typha x glauca has a N (% dry weight) content of 1-2 % with a phosphorus to nitrogen ratio of 1:1 (McJannet, et al. 1995). When Typha plants have reached their full capacity for taking up nutrients, the excess nutrients pass downstream opening up new areas for Typha to invade (Davis 1991). An increase in P and N in the waters of two different Norfolk broads in early fall indicates that a sizeable portion of nutrients are leached out at senescence (Mason and Bryant 1975). The facultative anaerobe Bacillus has been shown to have an associative relationship with the roots of T. latifolia, fixing nitrogen while root exudates provide carbon substrates to the bacteria (Bieboer 1984). Bieboer calculated that an average cattail stand with a density of 51 plants could fix 18 kg N2 ha-1 yr-1 which represents 8.2% of the 220-kg N2 ha-1 present in the standing crop.
Typha domingensis had a higher transpiration rate than did Cladium jamaicense and that difference was more pronounced in nutrient-rich sites (Koch and Rawlik 1993). As T. domingensis expands its range, it could have a significant effect on water loss in the Everglades and in other wetland ecosystems where it is expanding its range.
Control of Typha
Typha latifolia shoots have greater biomass in burned areas than they do in unburned areas (Smith and Kadlec 1985). Smith and Kadlec (1985) also reported preferential grazing in the burned areas, reducing the total annual production of Typha latifolia. Typha x glauca is most susceptible to injury from burning after the period of rapid growth in mid-July (Canada), but before the plants reach their maximum height in mid-August (Krusi and Wein 1988). At this stage of development most of the reserves of the plant are above ground. Chlorophyll a and b, nitrogen, phosphorus, and potassium in above ground biomass were at peak levels on April 23rd and then declined throughout the rest of the growing season (Boyd 1971). Initial shoot growth of T. latifolia was on March 24th in this study that was conducted in South Carolina.
Drainage can also decrease vigor and diminish the reserves of Typha because shoots emerge 1-2 weeks later, there is a faster rate of senescence, final shoot heights decrease by 10% and shoot density decreases by 10% as compared to the control (Krusi and Wein 1988).
When Typha was cut below water 95-100% of the belowground biomass died as compared to 15-20% for the uncut controls (Sale and Wetzel 1983). Sale and Wetzel (1983) found that three cuts made underwater during the growing season resulted in an oxygen shortage with ethanol being produced, accompanied by extensive tissue breakdown, and ultimately in almost complete death of the belowground biomass.
Conclusions
Typha angustifolia has expanded its range from 1949 to 2001 (Figures 1 and 2). It has been reported from 10 new states including Kansas, Kentucky, Louisiana, Montana, Nevada, New Mexico, Oklahoma, Oregon, Oklahoma, Tennessee, and Washington.
Typha domingensis has been reported from five new states that includes Arkansas, Colorado, Missouri, Nebraska, and Wyoming (Figures 3 and 4). Both T. angustifolia and T. domingensis may have been present in these new states in 1949, but had not been collected in those states up until that time. Typha latifolia was present in all 50 states in 1949 as well as 2001.
Competition studies have demonstrated that T. latifolia is a stronger competitor than either T. angustifolia or T. domingensis due to its ability to clone rapidly. It is also able to tolerate a wider range in temperature than either T. angustifolia or T. domingensis
The distribution of T. x glauca was not mapped by Hotchkiss and Dozier in 1949 and its identification and distribution remains problematic. T. x glauca is a hybrid that can exhibit the full range of leaf widths, spike width, spike interval, and stigma width found in T. angustifolia and T. latifolia. Differentiating the two parent species is difficult unless the plants exhibit the extreme leaf width, be at the large end of the range for the interval between staminate and pistillate portions of the spike. Only electrophoretic isozyme pattern tests of malate dehydrogenase and other enzymes will be conclusive for the identification of species that are intermediate in character traits.
Typha angustifolia will continue to expand into areas currently populated by T. latifolia where it will occupy the deeper water portions of the habitat. Typha angustifolia will probably never expand its range to the Arctic Circle as has T. latifolia, but the potential exists for T. x glauca to survive at latitudes further north than T. angustifolia.
Typha domingensis is a southern species that has not expanded very much over the last 52 years since it is limited by colder conditions. Unless agricultural runoff of chemical fertilizers is halted, it will continue to expand its range in the Florida Everglades and other similarly effected wetlands.
The low number of Typha seeds in the seedbank of the Everglades could be due to a number of different factors, including low survival rate, low seed production, or not enough time has passed for Typha seeds to accumulate.
Typha can be controlled through burning after the period of rapid growth in summer, draining of the wetland, or cutting plants below water. These techniques can be best employed in areas where water level can be controlled.
LITERATURE CITED
Bedish, J. W. 1967. Cattail moisture requirements and their significance to marsh management. American Midland Naturalist 78: 288-300.
Bieboer, D. D. 1984. Nitrogen fixation associated with natural and cultivated stands of Typha latifolia L. (Typhaceae). American Journal of Botany 71: 505-511.
Bonasera, J., Lynch, J., and Leck, M. A. 1979. Comparison of the allelopathic potential of four marsh species. Bulletin of the Torrey Botanical Club 106: 217-222.
Bonnewell, V., Koukkari, W. L., and Pratt, D. C. 1983. Light, oxygen, and temperature requirements for Typha latifolia seed germination. Canadian Journal of Botany 61: 1330-1336.
Boyd, C. E. 1971. Further studies on productivity, nutrient and pigment relationships in Typha latifolia populations. Bulletin of the Torrey Botanical Club 98: 144-150.
Boyd, C. E. and Hess, L. W. 1970. Factors influencing shoot production and mineral nutrient levels in Typha latifolia. Ecology 51: 296-300.
Davis, S. M. 1991. Growth, decomposition, and nutrient retention of Cladium jamaicense Crantz and Typha domingensis Pers. in the Florida Everglades. Aquatic Botany 40: 203-224.
DeBusk, W. F., Reddy, K. R., Koch, M. S., and Wang, Y. 1994. Spatial distribution of soil nutrients in a northern Everglades marsh: Water Conservation Area 2A. Soil Science Society of America 58: 543-552.
Dickerman, J. A. and Wetzel, R. G. 1985. Clonal growth in Typha latifolia: population dynamics and demography of the ramets. Journal of Ecology 73: 535-552.
Galatowitsch, S. M., Anderson, N. O., and Ascher, P. D. 1999. Invasiveness in wetland plants in temperate North America. The Society of Wetland Scientists 19: 733-755.
Grace, J. B. 1984. Effects of tubificid worms on the germination and establishment of Typha. Ecology 65: 1689-1693.
Grace, J. B. 1985. Juvenile vs adult competitive abilities in plant: size-dependence in cattails (Typha). Ecology 66: 1630-1638.
Grace, J. B. 1987. The impact of preemption on the zonation of two Typha species along lakeshores. Ecological Monographs 57: 283-303
Grace, J. B. 1988. The effects of nutrient additions on mixtures of Typha latifolia L. and Typha domingensis Pers. along a water-depth gradient. Aquatic Botany 31: 83-92.
Grace, J. B. 1989. Effects of water depth on Typha latifolia and Typha domingensis. American Journal of Botany 76: 762-768.
Grace, J. B. and Harrison, J. S. 1986. The biology of Canadian weeds. Canadian Journal of Plant Science 66: 361-379.
Grace, J. B. and Wetzel, R. G. 1981a. Phenotypic and genotypic components of growth and reproduction in Typha latifolia: experimental studies in marshes of differing successional maturity. Ecology 62: 789-801.
Grace, J. B. and Wetzel, R. G. 1981b. Habitat partitioning and competitive displacement in cattails (Typha): experimental field studies. The American Naturalist 118: 463-474.
Grace, J. B. and Wetzel, R. G. 1982. Niche differentiation between two rhizomatous plant species: Typha latifolia and Typha angustifolia. Canadian Journal of Botany 60: 46-57
Harris, S. W. and Marshall, W. H. 1963. Ecology of water-level manipulations on a northern marsh. Ecology 44: 331-343.
Hotchkiss, N. and Dozier, H. L. 1949. Taxonomy and distribution of North American cat-tails. The American Midland Naturalist 41: 237-253.
Jordan, T. E., Whigham, D. F., and Correll, D. L. 1989. The role of litter in nutrient cycling in a brackish tidal marsh. Ecology 70: 1906-1915.
Koch, M. S. and Rawlik, P. S. 1993. Transpiration and stomatal conductance of two wetland macrophytes (Cladium jamaicense and Typha domingensis) in the subtropical Everglades. American Journal of Botany 80: 1146-1154.
Koch, M. S. and Reddy, K. R. 1992. Distribution of soil and plant nutrients along a trophic gradient in the Florida Everglades. Soil Science Society of America 56: 1492-1499.
Kruse, B. O. and Wein, R. W. 1988. Experimental studies on the resiliency of floating Typha mats in a freshwater marsh. Journal of Ecology 76: 60-72.
Kuehn, M. M. and White, B. N. 1999. Morphological analysis of genetically identified cattails Typha latifolia, Typha angustifolia, and Typha x glauca. Canadian Journal of Botany 77: 906-912.
Kuehn, M. M, Minor, J. E., and White, B. N. 1999. An examination of hybridization between the cattail species Typha latifolia and Typha angustifolia using random amplified polymorphic DNA and chloroplast DNA markers. Molecular Ecology 8: 1981-1990.
Lee, D. W. and Fairbrothers, D. E. 1969. A serological and disc electrophoretic study of North American Typha. Brittonia 21, No. 3: 227-243.
Liu, E. H., Sharitz, R. R., and Smith, M. H. 1978. Thermal sensitivities of malate dehydrogenase isozymes in Typha. American Journal of Botany 65: 214-220.
Mason, C. F. and Bryant, R. J. 1975. Production, nutrient content and decomposition of Phragmites communis Trin. and Typha angustifolia L. Journal of Ecology 63: 71-95.
McJannet, C. L., Keddy, P. A., and Pick, F. R. 1995. Nitrogen and phosphorus tissue concentrations in 41 wetland plants: a comparison across habitats and functional groups. Functional Ecology 9: 231-238.
McMillan, C. 1959. Salt tolerance within a Typha population. American Journal of Botany 46: 521-526.
McNaughton, S. J. 1970. Fitness sets for Typha. The American Naturalist 104: 337-341.
McNaughton, S. J. 1974. Developmental control of net productivity in Typha latifolia ecotypes. Ecology 55: 864-869.
McNaughton, S. J. 1975. r- and K-selection in Typha. American Naturalist 109: 251-261.
Newman, S., Grace, J. B., Koebel, J. W. 1996. Effects of nutrients and hydroperiod on Typha, Cladium, and Eleocharis: implications for Everglades restoration. Ecological Applications 6: 774-783.
Reed, P. B. 1988. National list of plant species that occur in wetlands: National summary. Biological Report, 88(24), Washington, D.C.
Rivard, P. G. and Woodard, P. M. 1989. Light, ash, and pH effects on the germination and seedling growth of Typha latifolia (cattail). Canadian Journal of Botany 67: 2783-2787.
Sale, P. J. M. and Wetzel, R. G. 1983. Growth and metabolism of Typha species in relation to cutting treatments. Aquatic Botany 15: 321-334.
Segadas-Vianna, F. 1951. A phytosociological and ecological study of cattail stands in Oakland county, Michigan. Journal of Ecology 39: 316-329.
Shaffer, G. P., Sasser, C. E., Gosselink, J. G., and Rejmánek, M. 1992. Vegetation dynamics in the emerging Atchafalaya Delta, Louisiana, USA. Journal of Ecology 80: 677-687.
Sharitz, R. R., Wineriter, S. A., Smith, M.H. and Liu, E. H. 1980. Comparison of isozymes among Typha species in the eastern United States. Journal of Botany 67: 1297-1303.
Smith, L. M. and Kadlec, J. A. 1985. Fire and herbivory in a Great Salt Lake marsh. Ecology 66: 259-265.
Smith, S. G. 1962. Natural hybridization among three species of cattail (Typha) in California. American Journal of Botany 49: 678.
Smith, S. G. 1967. Experimental and natural hybrids in North American Typha (Typhaceae). The American Midland Naturalist 78: 257-287.
Stewart, H., Miao, S. L., Colbert, M., and Carraher Jr., C. E. 1997. Seed germination of two cattail (Typha) species as a function of everglades nutrient levels. Wetlands 17: 116-122.
Stuckey, R. L. and Salamon, D. P. 1987. Typha angustifolia in North America: a foreigner masquerading as a native. American Journal of Botany 74: 757
USDA, NRCS. 2001. The PLANTS Database, Version 3.1 (http://plants.usda.gov). National Plant Data Center, Baton Rouge, LA.
Van der Valk, A. G. and Rosburg, T. R. 1997. Seed bank composition along a phosphorus gradient in the northern Florida everglades. Wetlands 17: 228-236.
Waters, I. and Shay, J. M. 1990. A field study of the morphometric response of Typha glauca shoots to a water depth gradient. Canadian Journal of Botany 68: 2339-22342.
Waters, I. and Shay, J. M. 1992. Effect of water depth on population parameters of a Typha glauca stand. Canadian Journal of Botany 70: 349-351.