Clonal foraging in Trifolium repens
CHARLES H. WALKER
Department of Plant Biology, Southern Illinois University, Carbondale, Illinois 62901, USA
Summary
1 Foraging by means of elongating stolon internodes and petioles in low nutrient, high nutrient and control plots was examined in Trifolium repens.
2 Ten individual T. repens patches were sampled in each of the three 5 x 5 m plots.
3 Internode length showed no significant difference between plants growing in the three plots.
4 There was no significant difference in petiole length between the plots due to fertilizer.
5 There was a significant difference in petiole length due to fertilizer x time interaction.
Introduction
Growth form in clonal species range from those that employ a ‘guerilla’ strategy to those that employ a ‘phalanx’ strategy. The ‘guerilla’ species bear widely-spaced ramets, exhibit less branching, and infiltrate the surrounding vegetation. The ‘phalanx’ species present a tight-packed advancing front of ramets that exclude other plants from the clonal territory (Doust 1981).
Glechoma hederacea was shown to exhibit a ‘guerilla’ growth form when nutrients were low or competition was present. When nutritional levels were high, or when there was no competition, G. hederacea displayed the ‘phalanx’ growth form (Slade & Hutchings 1987).
Wijesinghe & Hutchings (1997) showed that G. hederacea was able to forage for nutrients when they were in large patches (707 cm2), but not in smaller patches that had the same absolute quantity of nutrients. Similar results have been obtained in a different experiment. Biomass more than doubled where there was a concentration of soil resources in 11% of the area of a box (625 cm2), in comparison to having the resources spread evenly throughout the box (Birch & Hutchings 1994). These results indicate that there is a lower limit on patch size in which a species can successfully forage for nutrients.
An increase in branching in high resource patches can be the result of more growth. However, rhizome length between parent and daughter ramets is a response to patch quality and should be used as a test for plasticity in ramet placement (Cain 1994).
De Kroon & Hutchings (1995) disagree. They concluded that the response of stolon internode lengths or rhizome length ("spacers") to nutrient availability is usually insignificant. They found 1 out of 5 stoloniferous species and 1 out of 11 rhizomatous species actually shortened their stolon internode length and rhizome length respectively in response to higher levels of resource supply. Trifolium repens actually lengthened its internodes, as well as increased branching with higher levels of resources (de Kroon & Hutchings 1995). Mixed results are sometimes obtained in the same experiment. Rhizome lengths were shorter with increased levels of fertilization in Brachypodium pinnatum, but those in Carex flacca were not significantly different (de Kroon & Knops 1990).
The length of "spacers" can be extremely variable. Some contend that this variation is changed very little by the environment in which the plants are growing (de Kroon, Stuefer, Dong & During 1994).
However, studies have shown that variation in length of "spacers" can be due to environmental factors. Doust (1987) found internodes of Ranunculus repens from grassland plants were significantly longer in a glasshouse experiment than those in the field. She attributed this difference to increased moisture in the glasshouse rather than to light or nutrients. MacDonald & Lieffers (1993) found significant differences between the control side and the cool, competitive side in length of rhizome, longest rhizome, number of rhizomes entering, branch points entry, and internode length.
The complex relationships between different species and how this affects the length of "spacers" has been investigated. Trifolium repens was found to have longer internodes when growing from a site dominated by Holcus lanatus, regardless of whether it was Lolium perenne, Agrostis capillaris, or Holcus lanatus growing nearby as neighbors (Turkington, Hamilton & Gliddon 1991).
Rhizome length or stolon internode length also depends upon other movement parameters, e.g., the time between "moves", frequency distributions for rhizome or stolon internode lengths, number of daughter ramets, etc., and they should be measured (Cain 1994).
Besides movement parameters, other growth factors have been examined. Increased branching has been found to be a more reliable indicator of morphological plasticity in clonal plants. Four of the five stoloniferous species and seven of the eleven rhizomatous species responded in a positive manner to higher levels of resource supply (de Kroon & Hutchings 1995). De Kroon & Hutchings (1995) concluded that the plasticity of orthotrophic stems and roots enable plants to effectively exploit favorable patches. Other studies support this conclusion. The proportion of roots in G. hederacea has been shown to decline more in going from nutrient-rich patches to nutrient-poor patches than do ramets and above-ground biomass (Birch & Hutchings 1994).
Even within the same species there is a great deal of plasticity between genotypes.
In a study of Ambrosia psilostachya, 9 distinct clones from a wide range of soil salinities were collected from natural saline flats in the Great Plains, and potted for an experiment. The clones with the highest degree of salt tolerance had the lowest proportion of emerged shoots in the nonsaline end of their pots (Salzman 1985). This variation within the same species indicates a genetic component to plant foraging as well as plant growth. Similarly, Ranunculus repens transplanted from grassland or adjacent woodland reacted differently from one another in a glasshouse experiment. When subjected to low supplies of resources at the parent rosette, grassland plants consolidated resources in the parent rosette while woodland plants consolidated resources in the daughter ramets (Doust 1987).
Not all species of plants consolidate resources. Tillers of Elymus repens are independent of one another under normal conditions. Defoliation of a new tiller or an old tiller can cause 14CO2 to translocate through the rhizome, old to new and new to old respectively (Kleijn & Van Groenendael 1999). Humphrey & Pyke (1997) found that Elymus repens produced more intravaginal and rhizomal tillers in high nutrient destination cells than in low density destination cells and that nutrient status of the origin cell was not significant to those plants growing in high-nutrient destination cells. Elymus repens was shown to exploit favorable microsites in a heterogenous environment by growing more primary rhizomes into bare quadrants (Kleijn & Van Groenendael 1999).
Stochastic simulation models indicate that the overall success of plant foraging strategies is variable and dependent on a suite of environmental conditions (Cain, Dudle & Evans 1996).
In this experiment the effect of fertilizer on the growth of Trifolium repens was examined. The following 2 hypotheses were tested:
1 that internode length of stolons will be longer in plots fertilized than in those not fertilized, and
2 that petiole length will be greater in plots fertilized than in those not fertilized.
Materials and Methods
THE SPECIES
Trifolium repens is a perennial, clonal herb native to Eurasia. It is commonly planted along with grass seed in lawn seed mixtures from where it has spread to roadsides and other "waste places" throughout temperate North America. It has been listed as an invasive weed by some authors (Uva, Neal & DiTomaso 1997, Stubbendieck, Friisoe & Bolick 1994).
THE STUDY AREA
This field study was conducted at the Chautauqua Road Long-term Field Site. It is located on the south side of Chautauqua Rd, approximately 1000 m west of Southern Illinois University, Carbondale, Illinois.
SAMPLING PROCEDURES
Three plots from within a block of 9 plots were used in this study; each plot measured 5 meter by 5 meters. All 3 plots were mowed last spring. One of the 3 plots was fertilized in the fall of 1998, 1 is fertilized annually, and 1 has received no fertilizer. Spring fertilizer for the plots has been 386 g diammonium phosphate, 304 g potash, and 349 g of ammonium nitrate. Fall fertilization was 350 g of ammonium nitrate per block.
A caliper was used to measure the internode length of the stolons as well as the length of the petioles of each leaf. Ten internodes and ten petioles from each of the 3 plots were sampled over a period of 2 weeks. Two-way analysis of variance (ANOVA) was conducted to test the effect of fertilization on petiole length and stolon internode length.
Log10-transformed end petiole length data were analysed.
Results
The first hypothesis had to be rejected because there was no significant difference in internode lengths between plots that had been fertilized and those that had not been fertilized.
The second hypothesis also had to be rejected, as there was no significant difference in petiole length due to fertilizer treatment. However, there was a significant difference in petiole length between plots due to fertilizer-time interaction (F2, 27 = 10.92l O < 0.0001). Petiole length in the annually fertilized group increased from a mean of 6.78 cm to 12.465 cm by the end of the data collecting period. The plot that had been fertilized in 1998 and the control group increased from 9.33 cm to 13.645 cm and 8.29 cm to 12.65 cm respectively (Fig. 1).
In this experiment the last or end internode showed a trend towards being shorter with increased fertilizer (F2, 27 = 3.03; P = 0.0652). The mean internode length of the group fertilized annually was 2.61 cm, the group fertilized in only the first year 2.54 cm, and the control group was 3.43 cm (Fig. 2).
Discussion
Internode length was not significantly different between the fertilized groups and the control group. In earlier studies (Solangaarachchi & Harper 1987; Thompson & Harper 1988; Caradus & Chapman 1991) stolon internode lengths had lengthened in response to higher levels of resource supply.
The trend towards shorter internodes in the last or end internode of the fertilized groups may have been due to irregular lengthening, i.e., not all of the end internodes were fully lengthened.
The significant difference between plots due to fertilizer-time interaction should be an area for future study. Petioles were slow in lengthening in the annually fertilized group as compared to the other 2 groups. The petioles of the annually fertilized group lengthened in a relatively short period of time (4 days). It is unknown why this occurred.
Valerianella radiata, an annual, grew around the outer edge of the Trifolium repens phalanx in the annually fertilized plot. Competition between these 2 species may be an area for further research.
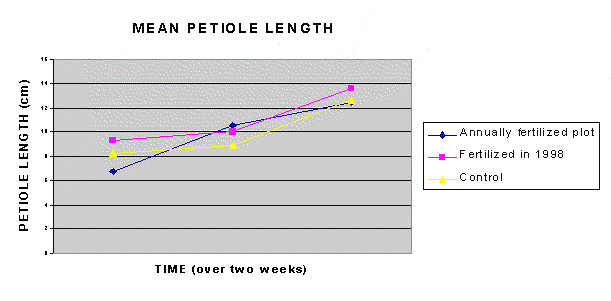
Fig. 1 Mean end petiole length through time. Ten end petioles were measured from each patch in each of the three plots, i.e. the annually fertilized plot, the plot fertilized in 1998, and the control plot. The ten measurements from each of the three plots were averaged to obtain the mean end petiole length.
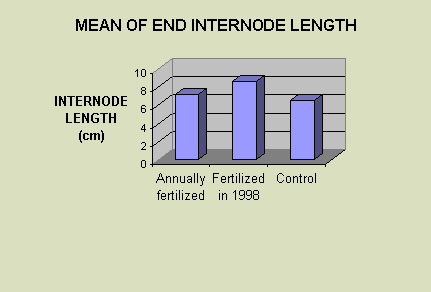
Fig. 2 Mean of end internode length. Ten end internodes were measured from each patch in each of the three plots, i.e. the annually fertilized plot, the plot fertilized in 1998, and the control plot. The ten measurements from each of the three plots were averaged to obtain the mean end internode length.
References
Birch, C.P.D. & Hutchings, M.J. (1994) Exploitation of patchily distributed soil resources by the clonal herb Glechoma hederacea. Journal of Ecology, 82, 653-664.
Cain, M.L. (1994) Consequences of foraging in clonal plant species. Ecology, 75, 933-944.
Cain, M.L., Dudle, D.A. & Evans, J.P. (1996) Spatial models of foraging in clonal plant species. American Journal of Botany, 83, 76-85.
Caradus, J.R. & Chapman, D.F. (1991) Variability in stolon characteristics and response to shading in two cultivars of white clover (Trifolium repens L.). New Zealand Journal of Agricultural Research, 34, 239-247.
Doust, L.L. (1981) Population dynamics and local specialization in a clonal perennial (Ranunculus repens). Journal of Ecology, 69, 743-755.
Doust, L.L. (1987) Population dynamics and local specialization in a clonal perennial (Ranunculus repens). III. Responses to light and nutrient supply. Journal of Ecology, 75, 555-568.
Humphrey, L.D. & Pyke, D.A. (1997) Clonal foraging in perennial wheatgrasses: a Strategy for exploiting patchy soil nutrients. Journal of Ecology, 85, 601-610.
Kleijn, D. & Van Groenendael, J.M. ((1999) The exploitation of heterogeneity by a clonal plant in habitats with contrasting productivity levels. Journal of Ecology, 87, 873-884.
de Kroon, H. & Hutchings, M.J. (1995) Morphological plasticity in clonal plants: the foraging concept reconsidered. Journal of Ecology, 83, 143-152.
de Kroon, H. & Knops, J. (1990) Habitat exploration through morphological plasticity in two chalk grassland perennials. Oikos, 59, 39-49.
de Kroon, H., Stuefer, J.F., Dong, M. & During, H.J. (1994) On plastic and non-plastic variation in clonal plant morphology and its ecological significance. Folia Geobotanica & Phytotaxonomica, 29, 123-138.
MacDonald, S.E. & Lieffers, V.J. (1993) Rhizome plasticity and clonal foraging of Calamagrostis canadensis in response to habitat heterogeneity. Journal of Ecology, 81, 769-776.
Salzman, A.G. (1985) Habitat selection in a clonal plant. Science, 228, 603-604.
Slade, A.J. & Hutchings, M.J. (1987) The effects of nutrient availability on foraging in the clonal herb Glechoma hederacea. Journal of Ecology, 75, 95-112.
Solangaarachchi, S.M. & Harper, J.L. (1987) The effect of canopy filtered light on the growth of white clover Trifolium repens. Oecologia, 72, 372-376.
Stubbendieck, J., Friisoe, G.Y.& Bolick, M.R. (1994) Weeds of Nebraska and the Great Plains, pp. ?. Nebraska Department of Agriculture, Bureau of Plant Industry, Lincoln, Nebraska.
Thompson, L. & Harper, J.L. (1988) The effect of grasses on the quality of transmitted radiation and its influence on the growth of white clover Trifolium repens. Oecologia, 75, 343-347.
Turkington, R., Hamilton, R.S. & Gliddon, C. (1991) Within-population variation in localized and integrated responses of Trifolium repens to biotically patchy environments. Oecologia, 86, 183-192.
Turkington, R. & Klein, E. (1991) Integration among ramets of Trifolium repens. Canadian Journal of Botany, 69, 226-227.
Uva, R.H., Neal, J.C. & DiTomaso, J.M. (1997) Weeds of the Northeast, pp. ?. Cornell University Press, Ithaca, New York.
Wijesinghe, D.K. & Hutchings, M.J. (1997) The effects of spatial scale of environmental heterogeneity on the growth of a clonal plant: an experimental study with Glechoma hederacea. Journal of Ecology, 85, 17-28.