Filogenia
y Sistemática
Los elementos biogeográficos
La pregunta motriz de la biogeografía es: ¿Cómo se
distribuyen las especies sobre la Tierra?
La observación de los patrones de ocupación
espacial del planeta por las diversas especies de animales y plantas dieron
origen a concepciones, en algunos casos antagónica, acerca del origen y
evolución de la vida sobre el planeta.
Linneo (mediados del siglo XVIII) propuso la
hipótesis de un centro de dispersión nico "Una sola isla" poblada de
especies distintas a partir de la cual se dispersarían luego de que los continentes
sumergidos aparecieran en su forma actual. Esta concepción tiene notables
raíces judeo-cristianas en particular si se la compara con la historia de Noé y
el arca en el monte Ararat.
Sin embargo, y a pesar de las buenas intenciones de
Linneo, esta concepción requiere de un par de situaciones hipotéticas:
a. Las especies aparecen en un lugar particular.
b. Las especies se dispersan a partir de dicho
lugar.
Pero si las especies se originan en un mismo lugar,
cómo es posible que haya diversidad de especies si están sometidas a
condiciones ambientales y estructurales similares?. Dado que Linneo había
propuesto con anterioridad que diferentes especies correspondían a su vez a
diferentes ambientes, la única alternativa era que el supuesto lugar original
tuviera una particular variabilidad de ofertas ambientales lo cual fue muy
criticado.
Buffon (siglo XVIII) encontró por su parte que
áreas con condiciones físicas aparentemente semejantes no tenían prácticamente
especies en común. Por ejemplo, si se compara la fauna del trópico de Africa
con la del trópico asiático o americano, se observará que el número de especies
comunes es muy bajo.
Esta "anomalía" conocida como Ley de
Buffon representa uno de los primeros antecedentes del concepto de
convergencias evolutivas y de especies homólogas.
Buffon concluyó, por su parte, que las aparentes
semejanzas probablemente no eran tales sino que a lo mejor algún factor crítico
era pasado por alto; con lo cual dejaba intacta la concepción original de
Linneo.
Más adelante, De Candolle (inicios del siglo XIX)
identificó el concepto de endemismo, lo que el llamaba especie aborigen, y las
relaciones entre la geografía de un lugar y la distribución de especies. Su
concepto de endemismo, sin embargo, incluía además la noción de especie típica
o característica de una geografía particular.
Darwin y Wallace (siglo XIX) establecieron
independientemente las bases para la biogeografía contemporánea.
Darwin basó sus apreciaciones en tres hechos
registrados por él mismo (1859):
1.Las semejanzas y diferencias entre colectivos de
especies no pueden ser explicadas completamente por factores climáticos y
físicos en general.2.Las barreras para la dispersión están relacionadas con las
diferencias que existen entre los lugares mismos. 3.La similitud de los
componentes de un mismo continente o área geográfica, aunque las especies
mismas sean diferentes.
Wallace, por su parte, dividió el planeta en
regiones biogeográficas que habrían de perdurar hasta nuestros días:
Paleártica, Neártica, Neotropical, Etiópica, Oriental y Australiana.
Henig (~1950) estableció las bases de la llamada
sistemática filogenética. A través de ésta se postula que la sistemática debe
reflejar las relaciones filogenéticas de parentesco (relaciones genealógicas).
Henig diferencia estas relaciones de parentezco de las relaciones de semejanza
(fenéticas) que forman la base de un tipo de taxonomía llamada taxonomía
fenética la que establece relaciones entre organismos en base a su similitud.
La sistemática filogenética (también denominada
cladista) establece las relaciones a partir de caracteres homólogos derivados
(sinapomorfías) evitando los caracteres homólogos antiguos (simplesiomorfías) o
bien las evoluciones paralelas o convergentes (homoplasias).
Finalmente, Wegener postuló alrededor de 1915 una
controvertida teoría que señalaba que la superficie del planeta no era fija,
sino que los continentes se desplazaban sobre una fluida capa interior del
planeta. Si bien Wegener apoyó su propuesta en evidencia fósil, como el caso del
reptil paleozoico Mesosurus sp. cuyos restos han sido encontrados a ambos lados
del oceáno Atlántico y cuyo estudio anatómico refleja una muy limitada
capacidad de dispersión propia, la hipótesis de la tectónica de placas no fue
bien recibida por sus contemporáneos.
Recién hacia fines de los 60 e inicios de los 70 se
consolidó la teoría y se propusieron alternativas para una dispersión más bien
pasiva de especies a bordo de continentes móviles .
La propuesta más aceptada hoy en día señala que en
un principio se contaba con una gran superficie de agua y dos grandes
continentes: Godwana al sur y Laurasia al norte del ecuador. La primera se
habría dividido en Eurasia y América del Norte, y la segunda en América del
Sur, Antártida, Australia, Africa e India, la que finalmente colisionó con
Eurasia para llegar a su ubicación actual.
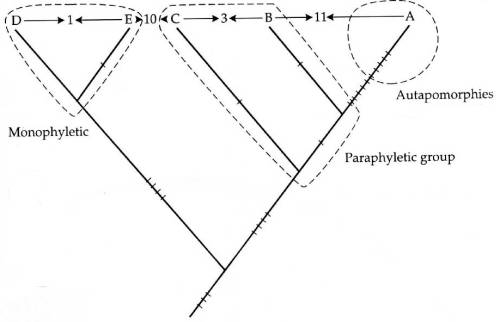
Debe reflejar la historia evolutiva
Solo se debe reconocer grupos taxonómicos que son monofiléticos, que
1.Tienen un ancestro en común y que
2.Incluyen todos los decendentes de este ancestro
en común
plesiomorfia: un carácter ancestral
apomorfia: un carácter derivado
sinapomorfia: un carácter derivado compartido entre
dos o más de dos taxones
autapomorfia: un carácter derivado que se encuentra
en un solo taxón o línea evolutiva
grupo monofilético: un taxón que incluye todos los
miembros de un clado o todos los descendientes de un taxón ancestral; ejemplos
incluyen: las plantas verdaderas (Reino Plantae), los hongos verdaderos (Reino
Fungi o Mycota) y todas las divisiones estudiadas en este curso excepto las
Divisiones Bryophyta, Chlorophyta y quizás Pterophyta
grupo parafilético: un grupo de organismos que
incluye algunos pero no todos los miembros de un clado, usualmente debido a la
exclusión del clado de taxones que poseen autapomorfias notables; ejemplos
incluyen: las dicotiledóneas (Clase Magnoliopsida), las gimnospermas, los
pteridófitos, los briófitos (División Bryophyta) y las algas verdes (División
Chlorophyta)
CRITERIOS DE HUTCHINSON PARA LAS ANGIOSPERMAS
LAS
PLANTAS CRIPTÓGAMAS Y FANERÓGAMAS
Los
términos «planta» y «animal» se equipararon originariamente con los dos grupos
sistemático taxonómicos tradicionales de los seres vivos («regnum
vegelabile» y «regnum animale»). Hoy sabemos que se trata de tipos
de organización diferentes en cuanto a la fisiología de la nutrición y no de
grupos de parentesco «naturales». Así pues, el «reino vegetal» no constituye
tampoco ninguna línea de descendencia y, por consiguiente, ningún taxón. Por el
contrario, el estudio de la constitución ultraestructural de la célula ha
puesto de manifiesto en los organismos la existencia de dos planos
estructurales básicos y diferentes, no relacionados entre sí por formas
intermedias: protocitos y eucitos. Estos dos planos estructurales de la célula
se consideran de forma muy general característicos de los dos táxones básicos y
«naturales» de los seres vivos, clasificados como «reinos»: procariontes
y eucariontes.
En
general son organismos inmóviles con paredes celulares sólidas que sólo pueden
tomar el alimento disuelto (o gaseoso). quedan excluidos los animales uni y
pluricelulares (protozoos y metazoos = zoobiontes). Además, debemos tener en
cuenta que la delimitación entre plantas, hongos y animales en los eucariotas
de niveles inferiores de organización tropieza con dificultades (e.g., en los
mixófitos y en los euglenófitos.
Dentro
de los «reinos» procariota y eucariota, ordenamos como «divisiones» comunidades
de descendencia muy amplias, pero claramente coherentes, o sea, derivadas de un
grupo de antepasados común y «monofilético». Pero como las relaciones de
parentesco de estas divisiones a menudo no resultan claras, se ordenan en
primer lugar según la fisiología característica de su nutrición en dos grupos
fundamentales: autótrofos y heterótrofos. Estos indican tan sólo grupos con un
nivel dé desarrollo comparable (y no necesariamente con un origen común) y son
apropiados para una mejor exposición.
En
un tipo de organización (de cualquier amplitud) incluimos grupos de
organismos que concuerdan básicamente en características de su organización,
tanto externas (i.e., morfológicas), como internas (i.e., anatómicas y
citológicas). Los tipos de organización corresponden muchas veces a niveles
de desarrollo y, como tales, son adaptaciones que a menudo se han producido
independientemente, en respuesta a condiciones ambientales determinadas o al
desarrollo progresivo general de la organización. Caracterizan también un
determinado grado de desarrollo filogenético dentro de una serie progresiva.
Por esto, tales niveles de desarrollo o tipos de organización .(e.g.,
talófitos, líquenes, plantas con semilla, angiospermas simpétalas) comprenden a
veces grupos totalmente heterogéneos en cuanto al parentesco, y dividen, no en
raras ocasiones, conjuntos filogenéticos que, en realidad, desde el punto de
vista del parentesco y la sistemática, deberían estar más cerca. (Así, e.g.,
la unión de los clorófitos con los rodófitos y otros grupos al tipo de
organización de las algas eucarióticas oscurece la estrecha relación
filogenética de los clorófitos, briófitos, pteridófitos y espermatófitos.)
La
evolución de los organismos fue de todos modos un proceso muy complejo y se
describe sólo de forma muy insuficiente en el marco de una obligada disposición
lineal de los grupos taxonómicos y a través de un solo principio de
organización.
Dentro
de los procariotas, y de acuerdo con esta división de tipos de organización, se
distinguen los bacterios, con los característicos arquebacterios, los
eubacterios y las algas procarióticas, dentro de los eucariotas, por un lado
los mixófitos heterótrofos (A) y los hongos sensu stricto (B), y, por
otro, los líquenes simbióticos (D) junto con los briófitos y las plantas
vasculares (E). Los eucariotas inferiores (A, B, C y D) están estrechamente
relacionados entre sí y con los protozoos, y pueden incluirse en el subreino
protobiontes ~protobionta). A partir de ellos se han originado no sólo
los animales pluricelulares (subreino metazoos -metazoo = zoobiontes
-zoobionta), sino también las verdadderas plantas terretres o embriófitos
(E), que se consideran muchas veces como integrantes de otro subreino de los eucariotas:
los cormobiontes (cormobionta).
La
historia del sistema del reino vegetal se caracteriza por la variación de los
criterios fundamentales. El más conocido de los sistemas artificiales es
el sistema sexual establecido por Linné en 1735. Linné oponía a 23 clases de
plantas con flores una 24ta clase, «Cryptogamia», en la que incluía no
sólo los helechos, musgos, algas y hongos, entonces aún poco conocidos, sino
también algunas plantas superiores con flores difíciles de reconocer (Ficus,
Lemna) y asimismo los corales y las esponjas. Dentro de las plantas con
flores (Phanerogamia) establecía subdivisiones basadas sobre todo en la
disposición de los sexos en las flores y en el número, concrescencia, inserción
y relaciones de longitud de los estambres. Las criptógamas se pueden llamar hoy
día «plantas esporíferas», pues en ellas los nuevos individuos se desarrollan a
partir de gérmenes generalmente unicelulares (esporas); las fanerógamas podrían
designarse como plantas floríferas (antófitos) o, mejor, seminíferas
(espermatófitos).
Ya
el mismo Linné había intentado establecer un sistema natural de las plantas,
pero los creadores de los primeros sistemas formales importantes fueron
A. L. de Jussieu (1789), A. P. de Candolle (1819), St. Endlicher (1836) y otros.
Después de que se impuso la teoría de la descendencia, los sistemas de A. Braun
(1864), G. Bentham yJ. D. Hooker (1862-1883), A. Eichler (1883) y especialmente
la ordenación de A. Engler, todavía muy empleada en la actualidad, quedaron
limitados a la utilización taxonómica de los niveles de organización y de los
grados de desarrollo. El primer sistema realmente filogenético fue
ideado por R. von Wettstein (1901-1908). Los sistemas de uso corriente en el
momento actual representan distintas etapas en el camino que conduce de la
agrupación formal a la filogenética y a la sintética.
Si
se observan los diversos sistemas actuales, a menudo opuestos, se ve que las
diferencias que los separan son aún muy profundas. Ello muestra hasta qué punto
la sistemática y la taxonomía modernas adolecen aún de falta de base y cuán
necesarias son todavía nuevas investigaciones fundamentales antes de que se
pueda dar por obtenida una sistematización generalmente aceptable. Tampoco la
división que aquí se expone representa otra cosa que un intento de representar
de manera sucinta las principales relaciones existentes; algunas
simplificaciones se proponen en atención a la finalidad didáctica de esta guía.
Hasta
el momento se conocen más de 400 000 especies de plantas vivas. Unos dos
tercios de las mismas pertenecen a los espermatófitos (unas 800 gimnospermas y
240 000 angiospermas), unas 10 000 corresponden a los pteridófitos y 24 000 a
los briófitos. Se calcula que, entre los protobiontes, el número de especies
descritas de algas es de unas 23 000, el de hongos es de unas 100 000 y el de
Iiquenes unas 20 000. Hay que evaluar, finalmente, las especies de bacterios en
unas 1700 y las de cianofíceas en 2000. Si se tiene en cuenta el gran número de
especies nuevas que se describen cada año (especialmente de hongos y de
angiospermas) no parece impropio suponer que el inventario completo del reino
vegetal, que aún estamos muy lejos de haber concluido, superará ampliamente el
medio millón de especies.
CUADRO
EXPLICATIVO DE LA UBICACIÓN DE LAS CRIPTÓGAMAS
Como
ya se explicó anteriormente las criptógamas son una agrupación artificial,
desarrollada a fin de poder ubicar a todos aquellos seres que no presentaban
flores, allá por la época de Linné; sin embargo, como herramienta didáctica aún
se conserva a fin de agrupar a organismos de diversos reinos, según Witaker.
Como
se puede apreciar en el cuadro, dentro de la denominación “criptógama” se
incluyen diversos tipos de individuos que han sido clasificados en DIVISIONES,
desde las Bacteriophytas hasta las Pteridophytas.
|
TAXÓN |
Divisiones |
REINOS (Witaker) |
Linne |
|
Bacterias |
Bacteriophyta |
MONERA |
CRIPTÓGAMAS |
|
Algas Verde Azuladas |
Cianophyta |
||
|
Hongos |
Fungi (Myxomycota, Oomycota,Eumycota,, Líquenes) |
FUNGI |
|
|
Algas Rojas |
Rhodophyta |
PROTISTA |
|
|
Algas Pardas |
Phaeophyta |
||
|
Flagelados |
Xantophyta |
||
|
Dinoflagelados |
Pyrrhophyta |
||
|
Diatomeas |
Bacillariophyta |
||
|
Euglenas |
Euglenophytas |
||
|
Charas |
Charophyta |
||
|
Algas Verdes |
Clorophyta |
||
|
Musgos, Hepáicas |
Briophyta |
PLANTAE ó VEGETAL |
|
|
Helechos, Colas de Caballo |
Pteridophyta |
||
|
Plantas con Flores y semillas |
Spermatofitas |
FANERÓGAMAS |
Debido
a que ya en prácticas anteriores se han estudiado a los seres unicelulares –
que conforman gran parte de las criptógamas nos detendremos en aquellos grupos
que aún faltan por estudiar.
Las Rodófitas: están formadas por las algas
rojas , debido a la presencia del pigmento ficoeritrina., además de la
clorofila Estas algas pueden ser microscópicas, en una etapa de su ciclo
biológico o durante todo éste. Las más grandes se pueden encontrar en las
zonas al nivel de la superficie marina,
pegadas a las rocas o sobre otros organosmos como caracoles o conchas de
bivalbos. Son utilizadfos en la alimentación humana con el nombre de “yuyos de
mar” o “cochayuyo” (varias especies de Porphyra y Gigartina)Imagen
de varias especies de Gigartina)
Las
Phaeophytas o algas pardas
deben su color a la presencia de caroteno, violaxantina y fucoxantina, además
de la clorofila. Viven sobre rocas a bajas profundidades o flotando libremente,
como representante más característico tenemos a Macrocystis integrifolia,
que es una alga que puede medir muchos metros de longitud, su cuerpo está
constituido por una especie de “rama” que se produce de una especie de rizoma
basal, con una eje cilíndrico del que se producen estructuras laminares a
manera de hojas, que en su base presentan un pneumatóforo globoso, el cual es
una estructura llena de aire lo que le permite flotar (imagen de Macrocystis
integrifolia)
Las
Clorophytas o algas verdes: se caracterizan
por que el color que sobresale es el de la clorofila, pueden tener diversa
morfología desde laminar hasta filamentosa e incluso ser unicelular, ser marina
o de agua dulce. la más conocida dentro de las marinas es la llamada “lechuga
de mar” (Ulva lactuca)
Las
Charas o Charophytas son algas dulceacuícolas
que viven en el fondeo de lagos, pantanos o lagunas, presentan una morfología
más compleja que las anteriores, pues se puede distinguir ya una morfología con
rizoide, nudos y entrenudos, demás de una disposición muy ramificada formando
verticilos de sus “ramas”. Presentan reproducción asexual por medio de
bulbillos que se forman en la base de sus rizoides y sexual con órganos
reproductores están separados en masculinos (globos anteridiales) y femeninos
(núculas) , produciendo anterozoides (gametos masculinos) y arquegonios
(gametos femeninos)
En
la figura de la izquierda se observa una rama de Chara, en la figura de
la derecha una aproximación al globo anteridial (superior) y a la núcula
(inferior).
BRIOFITAS
Las Briofitas, conforman un amplio grupo formado por unas 22.000 especies de plantas
pequeñas que crecen habitualmente en zonas húmedas sobre el suelo, troncos de
árboles y rocas. Los briofitos son plantas embrionarias no vasculares (sin
vasos conductores) que incluyen musgos, hepáticas, y antocerotas. En su nivel de organización, los briofitos se
sitúan entre las algas verdes o clorofitos, de las que con gran probabilidad
descienden, y las Pteridofitas. Los briofitos son similares a las plantas superiores
(pteridofitas y spermatofitos) en las que el óvulo fertilizado se desarrolla en
un embrión, una masa celular dependiente del gametofito (la forma sexual). Sin
embargo, el briofito embrionario se desarrolla en esporofitos (formas
asexuales) que, a diferencia de las plantas superiores, permanecen casi por
completo dependientes de los gametofitos y carecen de hojas, tronco o raíces.
Verdaderos tejidos de conducción, como los presentes en los helechos y plantas
superiores, no existen en los briofitos.
Algunas especies de briofitos son acuáticas, y
otras pueden sobrevivir en zonas áridas y secas. Aunque su tamaño varía desde
el microscópico a los 30 cm, el briofito medio tiene una longitud entre 1,2 y 5
cm, y varía de verde a negro y a casi incoloro. Los briofitos más primitivos,
las hepáticas, tienen cuerpos planos, algunas veces sólo del espesor de una
célula. Los musgos tienen un cuerpo central que recuerda a un tallo (cauloide)
del que se desprenden pequeñas hojas (filoides)y que se prolonga en unas
estructuras del tipo de las raíces denominadas rizoides. Sin embargo, los
briofitos, más que a través de estas estructuras, absorben el agua directamente
de la base sobre la que crecen o del aire.
Todas las especies de briofitos (al igual que las
pteridofitas y spermatofitas) se caracterizan por la alternancia de generaciones. El embrión de la forma sexual madura a una forma
asexual pequeña, que permanece unida y dependiente de ella. La forma asexual
produce esporas, similares a las de las plantas inferiores, que son diseminadas
por el viento y otros factores para producir nuevas formas sexuales. Los
órganos sexuales de los briofitos son multicelulares.
ciclo biológico en:
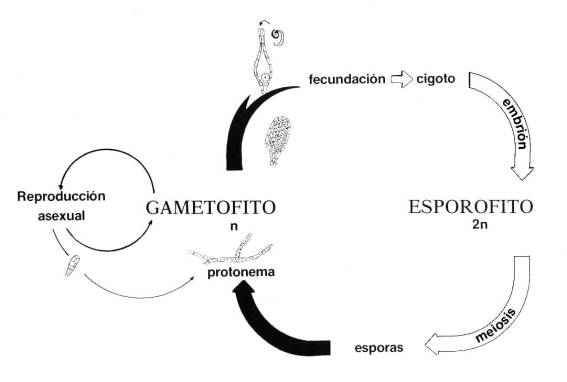
Más información sobre la clasificación de las
briofitas en: http://www.briolat.org/briofitas/
Las
Pteridofitas, son las criptógamas
más evolucionadas, pues presentan un cuerpo ya con raíz, tallo y hojas, lo cual
se denomina en conjunto CORMO. Son del reino vegetal (al igual que las fanerógamas,
que veremos luego), pues a diferencia de sus antecesores en su ciclo de vida
producen EMBRIÓN. Existen grupos ya extintos pero los que llaman más la
atención son el de los las “colas de caballo y el de los “helechos verdaderos”
o
Las colas de caballo
pertenecen al género Equisetum (en Chanchamayo: Equisetum giganteum y
crecen en las riberas rocosas de los ríos).Presenta rizoma perenne, que puede
estar a considerable profundidad de la superficie del suelo. Presentan ejes
estriados de ramificación verticilada, los nudos presentan anillos cerrados de
haces conductores denominado sifonostela, se encuentran separados entre sí por
entrenudos, en los entrenudos los anillos se descompone en cordones vasculares.
Hacía la medula es común hallar bandas de Caspary protegiendo a los haces
conductores.Las hojas son muy pequeñas y pierde la clorofila muy pronto, por lo
que la acción central de la misma está dada en el tallo. Los esporangios forman estróbilos o piñas, los que
nos recuerdan los conos de las gimnospermas. Los esporangios tienen una pared
pluriestratificada, presentando dehiscencia longitudinal. Las esporas presentan
hapterios que son producidos por la propia pared de la espora, las cuales se desarrollan como estructuras alargadas
que envuelven a las esporas y que les facilita la diseminación, actuando como
resortes. Son heterosporeos.
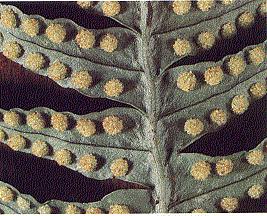
Los “helechos verdaderos” tienen tallos que
producen hojas compuestas denominadas frondes. Los esporangios se
agrupan formando estructuras protegidas por un tegumento denominadas soros.
Puede que tengan esporas de igual morfología (isosporeos) o diferentes
(heterosporeos). Existen helechos de
ambientes abiertos, con el género Adiantum
que presentan 56 especies en el Perú. las plantas de este genero son conocidos
como "culantrillo", sus hojas no se enrollan sobre sí durante su
desarrollo, presentan esporangios marginales a la hoja y libres , son
isosporeos. Entre los helechos de ambientes sombríos. tenemos como
representante a Polypodium, que es el helecho típico, de frondes que se van
desenrollando conforme crecen, tenemos especies herbaceas como el anterior y
arbóreas como Cyathea, de clima tropical, los soros se encuentran en el envés
como puntos grandes. Son isosporeos, los gametos se producen en la parte posterior
del protalo.
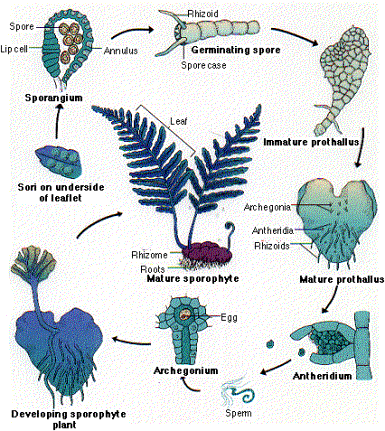
Mayor información sobre pteridofitas en: http://home.frognet.net/~jaknouse/ferns.htg/d_pter.html
Las Plantas Fanerógamas
(Spermatophyta)
Los
espermatofitos presentan alternancia de generaciones heteromorfa, con
gametofito y esporofito, como los briofitos y los pteridofitos y, asimismo,
alternancia de fases nucleares, como haplofase y diplofase. En los grupos de
mayor primitivismo todavía se reconocen con claridad, el gametofito femenino
arquegonios mientras que los anteridios del gametofito femenino están muy
reducidos. Al igual que en los pteridófitos actuales, el esporófito presenta la
diferenciación característica en raíz y vástago, y, a su vez, este último se
compone de ejes caulinares y hojas. Los espermatófitos pertenecen alos embriófitos o cormobiontes = corniófitos.
Sólo en 1851 Wilbelm Hofmeister
reconoció la alternancia de generaciones “disimulada”, que se da en los esper matótitos y con ello
apreció su estrecha afinidad respecto a bnófitos y ptendótitos. Entonces ya se
usaban términos particulares para denominar los órganos reproductores de los
espermatófitos y, aunque luego se haya establecido claramente la homología de
dichos órganos con los correspondientes de los pteridófitos, se han mantenido
simultáneamente basta el momento los dos sistemas de terminología Para las células y órganos reproductores
femeninos, en el dominio lingíiístico alemán se usó, en general, hasta el
momento, el prefijo griego 'rnacro-~, pero actualmente se prefiere el prefijo
también griego 'mega-~, de uso internacional.
Igual que los
pteridófitos superiores, heterospóreos, los espermatófitos forman también, tras
la meiosis, micrósporas (granos de polen uninucleados = células polínicas) y megásporas (= células del saco embrionario uninucleado). La reducción de los gametófitos o protalos
masculinos y femeninos (grano de polen o tubo polínico y saco embrional
pluricelulares) ha llegado tan lejos que dichos gametófitos no son visibles
externamente y muchas veces deben nutrirse a expensas del esporófito. Es
particularmente importante el hecho de que la megáspora al madurar no abandone
el megasporangio (= nucela del primordio seminal) y no se separe,
por lo tanto, de la planta madre esporófita. Así pues, el gametofito ftmenino (= saco embrionario) con las ovocélulas (a veces contenidas todavía en
arquegonios) se forma también sobre la planta madre. Mientras tanto, en los microsporangios
(= sacos polínicos) van madurando las micrósporas (= granos de polen
uninucleados). En este momento empieza ya el desarrollo del gametofito masculino, mediante, por lo menos, una división celular. Los granos de polen pluricelulares son conducidos hasta las inmediaciones de los megasporangios
y de los gametófitos femeninos (polinización) y, a continuación, forman un tubo polínico con espermatozoides (= espermios), o, en los casos comunes, con células
espermáticas desprovistas de flagelos. Se produce después la ftcundación de
la ovo-célula y el desarrollo del zigoto para dar el embrión. Al
mismo tiempo, junto al esporófito materno, se ha formado, a partir de la
cubierta del megasporangio (los 1-2 tegumentos del primordio seminal), una envoltura
(episperma, testa) en torno al embrión y a su tejido nutricio (endosperma): de este modo se ha formado, en lugar de la megáspora, un nuevo órgano
de multiplicación, la semilla. Estos cambios respecto a lo que sucedía en los
pteridófitos determinan que el proceso de la fecundación se haga independiente
del agua atmosférica y dan a los jóvenes esporófitos mejores posibilidades en
el inicio de su vida.
En cuanto a esporangios, encontramos en los espermatófitos megasporangios
con envoltura estéril [primordios
seminales, formados por nucela y tegumento(s)~ y grupos de microsporangios (grupos de sacos
polínicos). Estos elementos
morfológicos fundamentales de los órganos reproductores se insieren, aislados o
en grupos de pocos o muchos elementos, en soportes simples o ramificados de
modo más o menos complejo, los cuales pueden recibir los nombres de mega y
microsporofilos o carpelos
y estambres respectivamente.
Casi siempre los esporofilos se
sitúan, en los espermatófitos, sobre braquiblastos
de crecimiento limitado:en este caso se
habla de flores. Por ello, los espermatófitos también pueden recibir
el nombre de «antófitos» («anthophyta», plantas con flores). Las flores pueden ser unisexuales o
hermafroditas, según contengan sólo micro o megasporofilos o bien
contengan los dos tipos juntos. Sobre todo en las flores hermafroditas se
constituye muchas veces una envoltura fioral
(perianto). La disposición de los
micro y megasporofilos en las flores, que quedan separadas del cuerpo
vegetativo, facilita la polinización cuando la flor alcanza su pleno desarrollo
(antesis). En las gimnospermas («Gimnospermae»),
el polen es transportado sobre los
primordios seminales, en las angiospermas («Angiospermae»), sobre los
estigruas de los carpelos, lo cual hace posible la fecundación y el desarrollo
de las semillas. Los órganos que
contienen las semillas que van madurando o están maduras y contribuyen a la
diseminación de las mismas se llaman frutos.
Incluso en el embrión de los
espermatófitos primitivos se reconoce un polo caulinar y un polo radical
opuestos entre sí a partir de los
cuales se desarrollan el vástago primario y la raíz principal. Son
características del tallo de los espermatófitos la ramificación axilar, la
posesión de eustela y la capacidad de efectuar crecimiento secundario en
grosor. Primariamente se trata, pues, de plantas leñosas dotadas de un sistema
de absorción y conducción del agua
Los espermatófitos predominan en las
flores terrestres del mundo desde principios del Mesofítico (Pérmico superior).
Aunque puedan apreciarse inicios de constitución de caracteres análogos en distintos
grupos de pteridófitos superiores (e.g., reducción de los gametófitos,
heterosporia, grupos de esporofilos semejantes a flores, incluso formación de
semillas), el origen de los espermatófitos no puede suponerse posterior al
Devónico y tiene que haber arrancado de antepasados semejantes a psilófitos. El
desarrollo del grupo tiene que haber transcurrido paralelo a la expansión de
los pteridófitos.
Las
plantas con Flores (Antofitas, Antos flor fito, planta),
conocidas por Linné como Fanerógamas (Fanero, conspicuo, gama
unión), debido a que sus órganos reproductores, están al descubierto lo que
permite analizar su sexualidad o unión entre sexos, todo lo contrario de las
criptógamas (cripto, oculto). También son denominadas Spermatofitas (sperma,
semilla, fito, planta), pues son los vegetales que luego de la
fecundación, el embrión se va proteger dentro de la semilla, la que
posteriormente constituirá su primera fuente de nutrientes.
En
forma general las plantas con flores, según uno de los primeros sistemas de
clasificación propuesto por A. Engler a finales del siglo XIX, se subdividen en
dos grandes grupos: Las Gimnospermas (aquellas tienen sus semillas al
descubierto) y las Angiospermas (aquellas que protegen dentro de una estructura
su semilla). Estas últimas según Engler se dividen en: Dicotiledóneas (con 2
cotiledones) y Monocotiledóneas (con 1 cotiledón).
Tomando
como base los principios bioquímicos que constituyen a las plantas con flores,
en la década de 1950, el botánico A. Cronquist, propone la moderna clasificación, en la que las Gimnospermas
están divididas en 2 grupos: Coniferophyta y Cycadophyta, mientras que las Angiospermas,
se mantienen en un solo grupo denominándose ahora Magnoliophyta, el cual
conserva la división propuesta por Engler en Dicotiledóneas y Monocotiledóneas,
llamándose ahora Magnoliopsida y Liliopsida, respectivamente, según lo podemos
observar en el siguiente cuadro.
|
Taxón |
Engler |
Cronquist (Divisiones) |
REINO (Witaker) |
Linné |
|||
|
Pinos, Cipreses, Ginkgos, Podocarpos, Efedras |
Gimnospermas |
Coniferophyta |
Spermatophyta |
PLANTAE ó VEGETAL |
FANERÓGAMAS |
||
|
Cycas |
Cycadophyta |
||||||
|
Magnolias, Casuarinas, Castaños, Malvas, Algodones, Girasoles, Tulipán
Africano, Euforbias, Manzano, Fresas, Margaritas, Cítricos, Floripondio, Papa |
Clase Dicotiledóneas |
Angiospermas |
Clase Magnoliopsida |
Magnoliophyta |
|||
|
Grases, Gramíneas (maíz, trigo, cebada), Azucenas, Achira, orquídeas,
bromelias, piñas |
Clase Monocotinedoneas |
Clase Liliopsida |
|||||
Las
flores de los espermatófitos sirven para la reproducción sexual. Las flores son
conjuntos de hojas especializadas para la reproducción, estás tienen
crecimiento limitado provistos de órganos reproductores masculinos y femeninos
o sólo uno de ellos en el caso de las flores unisexuales.
En
los cicadofitos se puede seguir paso a paso el desarrollo de las flores:
primero dispersos sobre el eje principal que sigue creciendo, (e.g., en los
estróbilos de megasporofilos de Cycas,), esporofilos dispuestos helicoidalmente
y en gran número sobre ramas laterales de crecimiento limitado (. Ello
corresponde a una progresiva división del trabajo entre vástagos reproductores
(flores) y vástagos vegetativos (tallos, hojas)., como la que ya se observa
también en los pteridófitos (e.g., en Lycopodium, Selaginella o Equisetum). En
el curso del desarrollo evolutivo de las flores de los espermatofitos, el
número de piezas florales, inicialmente muy elevado, ha disminuido en muchas
ocasiones.
Las
flores de los espermatófitos son originariamente unisexuales (masculinas sólo
con estambres, o femeninas, sólo con carpelos); las flores hermafroditas
(bisexuales, con estambres y carpelos) aparecieron más tarde
Lo
que nos lleva a pensar de la posible evolución de las plantas con flores a
partir de las Pteridofitas.
Inicialmente
las flores, relativamente grandes, se presentan aisladas en distintos grupos de
espermatófitos. En relación con la tendencia general a la reducción del tamaño
de las flores se constituyen muchas veces en compensación, inflorescencias.
Las
Gimnospermas, son plantas que en su gran mayoría son árboles, llegando a
medir más de 100 metros de altura como es el caso de algunos Pinos y Cipreses,
además de ser muy longevos llegando a vivir miles de años (se ha registrado la
edad de un pino hasta en 4,000 años). Las hojas pueden ser escamosas (como en
los cipreses), aciculares (en forma de agujas, como en el pino), lanceoladas,
con nervaduras, bifurcadas (dicótomas); o en el caso de las Cycas con un plan
estructuras pennado (como un peine) Sus flores son muy sencillas, se agrupan en
inflorescencias en forma de conos y están reducidas a simples hojas que pueden
producir ya sea sacos con polen o polínicos (flores masculinas) u óvulos
(flores femeninas)
Las
Coníferas (Coniferophyta), pueden ser árboles (o arbustos (Ephedra),
sus hojas pueden ser bilobadas (Ginkgo), escamosas, aciculares o lanceoladas;
los conos son pequeños, presentándose tanto los sacos polínicos o los óvulos
–sea que se trate de flores masculinas o femeninas- en la cara ventral o haz de
las hojas florales. Las inflorescencias son sólo masculinas o femeninas. Una
vez que se ha producido la fecundación los conos femeninos se cierran y toman
una apariencia leñosa como en los pinos y cipreses, una vez que las semillas
están maduras los conos se abren y las dejan salir para su diseminación. Las
semillas pueden presentar tegumentos a manera de alas (eg. Pinos) lo que
le permite ser diseminado por el viento (anemocoria).
Las
Cycas (Cycadophyta) son gimnospermas arbóreas, sus hojas tienen
un plan estructural pennado y salen junto con las flores del extremo superior
del tronco (estípite). Las flores masculinas y femeninas se agrupan en inflorescencias
(conjunto de flores), sobre un eje que nos recuerda los conos de las coníferas,
pero son mucho más grandes.
Las
Angiospermas (Magnoliophyta) Pueden ser árboles, arbustos o hierbas; con hojas de diversa
morfología, y nervaduras predominando las reticulares (en forma de rejilla) o
las paralelas. Ya presentan las hojas que producen óvulos (hojas carpelares)
protegidos por una cubierta (Pistilo) que desarrolla estructuras
especializas conocidas como estigma (para captar el polen), estilo
(tubo compacto que une al estigma con el ovario) y el ovario (donde se
encuentran los óvulos). Las hojas que producen polen se han modificado en estambres,
encontrándose los sacos polínicos en las llamadas anteras, que
generalmente están sobre un filamento. Las flores tienen una envoltura
especializada (envoltura floral) que
protege a los órganos reproductores (pistilos y estambres). La envoltura floral
puede estar formada por dos grupos diferentes de hojas: el cáliz , que está
formado por los sépalos, que por lo general son de color verde; y la corola,
que está formada por los pétalos que son de vivos colores y tienen especial
aroma, La envoltura floral que presenta cáliz y corola se denomina perianto..
Cuando no se pueden diferenciar entre sépalos y pétalos, es
decir, cuando todos los miembros de la envoltura floral tienen igual color,
consistencia y textura, esta envoltura toma el nombre de Perigonio y
cada pieza toma el nombre de tépalo.
Las
Dicotiledóneas se Diferencian de las Monocotiledóneas, por las siguientes
características:
|
MAGNOLIOPSIDA |
LILIOPSIDA |
|
dos cotiledones |
un cotiledón |
|
raíz axonomorfa (y otras) |
raíz fibrosa o fasciculada |
|
hojas reticulinervias (en las nervaduras más pequeñas) |
hojas paralelinervias (en las nervaduras más pequeñas) |
|
presenta cambium |
no presenta cambium |
|
polen con apertura diversa |
polen con una sola apertura (monocolpado ) |
|
nº diverso de piezas florales |
flores por lo general trímeras, es decir, el cáliz tiene 3 sépalos, y
la corola 3 pétalos; o le perigonio 3 + 3 tépalos |