 The Genus Cymbidium
The Genus Cymbidium
The Genus Cymbidium
The Genus Cymbidium| In preparing a monograph, one of the tasks of the
botanist is to decide what to include and what to leave out. Inevitably,
in the past, a number of species have been given the name Cymbidium but are
now known to be better placed in other genera. Similarly, species which have
not yet been transferred to Cymbidium may have to be sought in other genera.
Before this can be done, the exact characters of the genus, features by which
it can always be recognised, need to be elucidated and described. The characters
traditionally used are those of vegetative and floral morphology. They are
described briefly below. |

|

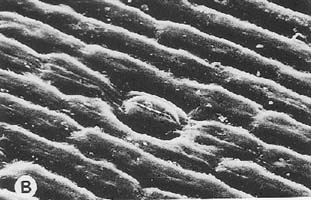
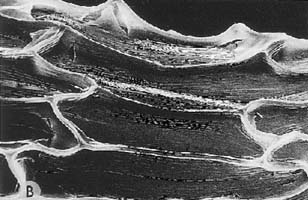
| In the subgenus Cyperorchis and in Cymbidium dayanum,
the subepidermal layer is lacking and the fibrous strands are isolated from
each other. These species also have smooth epidermal cells. In subgenus Jensoa,
however, where the bundles of fibres near the lower surface are also isolated,
the epidermal cells have a conspicuous papillose surface. The accompanying
photograph shows part of a leaf of a member of the subgenus Cyperorchis.
The small strands of fibres along the margins and the larger bundles surrounding
the vascular tissues of the leaf are similarly arranged in all the species
of this subgenus. |
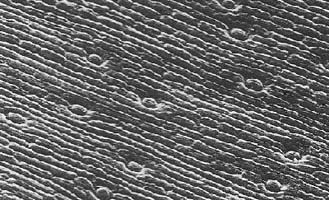
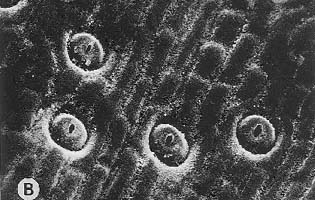
 showing crowded stomata (magnification x80)
showing crowded stomata (magnification x80) |
|
|

|
The first of the large-flowered cymbidiums to flower
in English glasshouses was C. iridoides (syn. C. giganteum). It was introduced
from the tropical parts of the Himalaya range and plants grew well at Chatsworth.
The dull greenish brown flowers rarely open fully and are shorterlived than
those of some other species. The plants are relatively large and the combination
of these features has made this species less popular with modern growers. |
