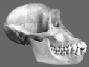
Were Humans Specially Created?
Left: Sts 5:
Australopithecus africanus: "extinct ape"
Right: ER 3733: Homo ergaster: "true" human.
"If we reject the natural explanation of hereditary descent from a common ancestry, we can only suppose that the Deity, in creating man, took the most scrupulous pains to make him in the image of the ape. This, I say, is a matter of undeniable fact -- supposing the creation theory true -- and as a matter of fact, therefore, it calls for explanation. Why should God have thus conditioned man as an elaborate copy of the ape, when we know from the rest of creation how endless are His resources in the invention of types?"
George J. Romanes, 1882
According to young-earth special creationism (YESC), humans do not share a common ancestry with any non-human species. Instead, YESC holds that humans were created instantaneously, along with all other kinds of organisms, about 7,000 years ago. Thus, any similarity humans might have with other species is either entirely coincidental, or based on similar "design objectives" on the part of the creator. YESC is disconfirmed by several independent lines of evidence -- the homonid fossil record, DNA sequence data, chromosomal analyses, and developmental processes -- all of which strongly support the Darwinian thesis that humans are very closely genealogically related to other extant species, such as the chimpanzee. In this essay, we will examine both theories in light of the fossil and genetic evidence.
1. Most special creationists hold that humans were specially created within a few days of the very first living things, as suggested by a literal reading of Genesis 1. The geological and paleontological evidence tells an entirely different story -- whereas single-celled life appears about 4 billion years ago, the first animal forms are found in the fossil record at least as far back as 570 million years ago, the first mammals approximately 200 million years ago, H. sapiens in particular does not appear in the fossil record until a mere 500 thousand years ago. If we think of the past 570 million years as one "day," then the emergence of H. sapiens in the fossil record occured at about 11:58PM. Obviously, this temporal pattern is consistent with evolution, which suggests that H. sapiens originated as a result of a long process of descent with modification, but not with the essentially simultaneous origin of humans and other animals predicted by special creation. This is the most basic imaginable test of YESC as an hypothesis, and it fails the test completely.

| Ardipithecus ramidus is known from about 5.2-5.8 Myr in Awash, Ethiopia (Late Miocene hominids from the Middle Awash, Ethiopia. Nature 2001 Jul 12;412[843]178-81). The earliest australopith, A. anamensis, is known from about 4.17+/-0.03 in Kenya (New specimens and confirmation of an early age for Australopithecus anamensis. Nature 1998 May 7;393[680]62-6). Afarensis is known from around 3.4-3.0My from the Hadar Formation in Ehtiopia. Habilis is known from about 2.3Myr or so, also from Hadar (Am J Phys Anthropol 1997 Jun;103(2):235-62). Ergaster, the first homonid known outside of Africa, appears about 1.9Myr or so (The Human Genus. Science 1999 Apr 2;284[5411]:65-71). Ergaster is now known from Dmanisi in the former Soviet republic of Georgia by about 1.6-1.8My, very close to the time they appear in Africa (Science 288, 1019-1025. May 12, 2000). |
2. Despite creationist claims to the contrary, there exists an excellent series of "intermediate" homonid forms illustrating the emergence of specifically human anatomic features over the past 5 million years or so. In the sequence A. afarensis, A. africanus, H. habilis, H. rudolfensis, H. ergaster, and H. erectus, we see precisely those changes predicted by the theory that humans descended from ape-like ancestors (note: this is not presented as a case of lineal descent, but as an illustration of the distribution of character states in time). Consider the large brain of Homo, for instance. Braincase size in afarensis is about 350-500cc, about the same as chimps. In H. habilis, the earliest known member of the genus Homo, the braincase volume varies between about 500 and 800cc. Falk has argued that Broca's area, a speech processing area located on the left frontal lobe (Falk, p 50), is present in 1470. In early ergaster/erectus specimens, the braincase averages about 900cc, while later erectus averages about 1100cc, with largest erectus at about 1225cc, which is within the human range.
Besides increasing cranial capacity, we can also see other changes occuring in the lineage over time -- the protrusion of the jaw becomes more and more diminished, the teeth, especially the canines, become smaller, the shape of the jaw becomes increasingly parabolic rather than rectangular, the brow ridges become less pronounced, the forehead assumes an increasingly more vertical orientation as the braincase increases in size, and so on. The further back in time the homonid lineage is traced, the more ape-like it appears to be. What's more, there is no obvious dividing line in the homonid lineage, no unbridgable 'genus gap' where "ape" sudddenly becomes "human." The whole sequence is morphologically gradual. For example, although ER 1470 is classed in the genus Homo, but it could just as easily be classed in the genus Australopithecus -- it is purely a matter of definition and nomenclature. In fact, the name Australopithecus habilis was proposed for ER 1470.
|
|
|
|
|
A chimpanzee skull. Not a human ancestor, but the closest living approximation. Braincase about 400cc. |
ER 1813 H. habilis. Considered a member of the ape-kind by all special creationists. Braincase about 510cc. |
ER 1470 H. habilis [or Rudolfensis]. Considered human by some creationists (Lubenow), ape by others (Gish). Braincase about 700cc. |
Turkana Boy. H. erectus. Considered "fully human" by almost all creationists. Other creationists (Hugh Ross [1995, p. 22], John Wiester [1983, p. 173], Percival Davis and Dean Kenyon [1993, p. 10]) assert on the contrary that erectus isn't even related to humans! Braincase about 1000cc. |
H. sapiens. Modern human. Note the many new features -- vertical forehead, jutting chin, reduced jaw protrusion, larger brain case, about 1400cc. |
If you fail to see the huge anatomic gap seperating the human-kind from the ape-kind in this sequence, you're not alone. Depending on which creationist you consult, the non-human/human gap is between either 1813 and 1470, or between 1470 and H. erectus. Either way, such an anatomic "gap" exists only in the imagination of special creationists, and is not supported by the fossils themselves.
If humans were specially created, then they were created as the most recent iteration a series of a progressively more human-like creatures, with the resulting fossil sequence indistinguishable from that predicted by evolution. Afarensis first appears about 4 million years ago (mya), africanus about 3 mya, habilis about 2.5 mya, erectus about 1.8 mya, archaic sapians about 500 thousand years ago (kya), and fully modern humans about 100kya.
1. In the whole 600+ million year history of multicellular life on the surface of planet earth, why do the big-brained, bipedal "apes" (e.g. ER 1470 and the habilines) happen to appear in the fossil record immediately (>1My) before the earliest "true" humans? (whatever "true" means in this case)
2. Given all the variation seen in human populations, why do the earliest "humans" all possess such small cranial volumes (800-1200cc, sloping foreheads, rounded chins, and prominant brow ridges? Why, in other words, do early humans look so apish, and become increasingly more "human" as a function of time?
3. Why, of all the space on planet earth, do these big-brained human-like apes appear in the same geographic place as the earliest, ape-like humans?
3. The degree of genetic relatedness between humans and chimpanzees is very near 100%, and is far greater than would be expected on the basis of either chance or design. DNA hybridization experiments and sequence analysis shows that humans share about 98.5% of their genetic code with bonobo chimpanzees. By way of contrast, humans differ from each other by about 0.5%, and different species of fruit fly differ from each other by about 3.0% percent. In terms of genetic information alone, humans are more closely related to chimpanzees than chimpanzees are to gorillas and other living primates. This overwhelming genetic homology extends not only to functional gene sequences, but also to highly-variable junk DNA regions. Chen FC, and Li WH report:
To study the genomic divergences among hominoids and to estimate the effective population size of the common ancestor of humans and chimpanzees, we selected 53 autosomal intergenic nonrepetitive DNA segments from the human genome and sequenced them in a human, a chimpanzee, a gorilla, and an orangutan. The average sequence divergence was only 1.24% +/- 0.07% for the human-chimpanzee pair, 1.62% +/- 0.08% for the human-gorilla pair, and 1.63% +/- 0.08% for the chimpanzee-gorilla pair. These estimates, which were confirmed by additional data from GenBank, are substantially lower than previous ones, which included repetitive sequences and might have been based on less-accurate sequence data. The average sequence divergences between orangutans and humans, chimpanzees, and gorillas were 3.08% +/- 0.11%, 3.12% +/- 0.11%, and 3.09% +/- 0.11%, respectively, which also are substantially lower than previous estimates. The sequence divergences in other regions between hominoids were estimated from extensive data in GenBank and the literature, and Alus showed the highest divergence, followed in order by Y-linked noncoding regions, pseudogenes, autosomal intergenic regions, X-linked noncoding regions, synonymous sites, introns, and nonsynonymous sites. Genomic divergences between humans and other hominoids and the effective population size of the common ancestor of humans and chimpanzees. Am J Hum Genet 2001 Feb;68(2):444-56.
For example, the protein Cytochrome C is identical in chimps and humans, and both differ from the Rhesus mokey by only one amino acid. Alpha and Beta hemoglobin chains are identical in humans and chimpanzees, but differ from gorilla and gibbon chains by 1 and 3 amino acids respectively. Delta hemoglobin in humans differs from chimps and gorillas by only one mutation, and differ from gibbons by only two mutations. Out of 153 amino acids which compose myoglobin in chimp, the human myoglobin differs in only 1. And so on. Colin Patterson observes that "the general conclusion from the molecules that have so far been investigated is that the genetic differences between a gorilla and a chimpanzee, minor to out eyes, are actually slightly greater than than those between a human and chimpanzee . . . In genetic terms, we are hardly more distinct from chimpanzees than are subspecies in other groups of animals" (p112).
To understand why these similarities are far greater than would be expected on the basis of either chance or design, it is necessary to discuss the relationship between truly identical proteins and functionally identical proteins. In reality, there are a diversity of functionally identical macromolecules which go under the name cytochrome c or apha, beta and delta hemoglobin, for example. Out of the 104 amino acids which compose cytochrome c in modern organisms, only 3 amino acids (Cys-17, His-18, and Met-80) are are found to be totally invariant! In all 101 other positions on the chain, at least some variation is allowable. These cytochrome molecules can be shown with x-ray crystallography to posses the same three-dimensional structure, and to carry out the same metabolic functions. What's more, through the use of recombinant genetic technology, the genes which code for these various cytochrome c variants (say, chimp) can be inserted into different organism (say, a sheep), again demonstrating the functional identity of these proteins.
Just as human cytochrome c differs from that of chimps by only one protein, the cytochrome c gene in humans differs from that of chimps by only a single codon. To calculate the probability that humans and chimps would share 103 out of 104 codons which code for their cytochrome c's, it is necessary to estimate the number of codon sequences which code for functionally identical forms of cytochrome c.
As we said above, almost all of the proteins in cytochrome c are variable, and only three proteins are totally invariant among all examined organisms. For the sake of an extremely conservative estimate, however, we'll assume that all cytochromes must be composed of the same amino acids, and that variability exists only with respect to the codons which code for those particular amino acids. If we assume that each of the 20 amino acids is coded for by an average of 3 different codons [64 codons, minus 3 STOP codons], this means that there are at least 3^104, or over 10^49 possible ways to produce identical cytochrome c molecule, that is, with the exact same sequence of amino acids. The number of possible cytochrome c gene sequences is in reality exponentially higher, since this calculation assumes that one and only one sequence of amino acids will work, which we have just shown to be false (some yeasts, for example, differ from humans in 51 out of 104 amino acids).
Why then do humans and chimps share 103 out of 104 codons in their cytochrome c genes? This degree of similarity is certainly not expected on the basis of chance convergence, since the probability that two such identical sequences would form independently is astronomically low. This degree of similarity is also not expected on the basis of design constraints or objectives, since their are no design-based reasons for choosing between functionally identical gene sequences. The theory of common descent is the one theory which not only explains this genetic convergence, but absolutely predicts it. Thus, according to common descent, humans share specific sequences with chimps and other primates for the same reason that siblings share such specific sequences with each other -- because both species have inherited their DNA from a common ancestor.
4. While the similarities in functional proteins and functional genes provides compelling evidence for the common descent of humans and other primates, even clearer evidence is provided by an examination of highly-variable noncoding DNA regions between species. Only about 5-10% of the genetic material in humans and other vertebrates actually codes for proteins. The vast majority of the vertebrate genome is composed not of protein-encoding genes or regulatory elements, but of "junk" DNA -- large numbers of duplicated pseudogenes, Alu sequences, retroposons, transposable elements. These non-coding regions are not translated into proteins, and thus do not contribute to the function of the organism which carries them.
The genetic information which is actually expressed as proteins is embedded in this much larger background of genetic gibberish left behind by random gene duplications, nucleotide additions and deletions, transposing elements, retroviral infection, and other genome-altering events. The high variability in these noncoding genetic elements allow hypotheses about geneaological relatedness between species to be tested with more precision than is possible with the less-variable protein-coding genes. Such analyses are used every day to establish genealogical relationships within species. Unfortunately for special creationism, the very same analyses can be used to demonstrate genealogical relationships between species. Let's look at some examples.
Pseudogenes originate in a number of ways. One form of pseudogene originates when a once-functional gene is rendered nonfunctional by a mutation. Though no longer functional, the "vestigial" gene is nonetheless copied along with the rest of the genome and passed on to future generations. If speciation occurs within the lineage, thee the descendent species will share the pseudogene as well.
One example of shared pseudogenes offered by Max (1998), that of the L-gulono-gamma-lactone oxidase, or LGGLO, gene is particularly interesting. LGGLO is an enzyme necessary for the synthesis of vitamin C. While most mammals possess functional LGGLO enzymes, and can synthesize their own vitamin C, guinea pigs, most primates and humans do not possess functional LGGLO. Each of these groups have diets rich enough in vitamin C that the loss of the gene to mutation is without consequence. Now, given the widespread prevalence of functional LGGLO in most other mammals, the most parsimonious assumption is that functional LGGLO was possessed by the ancestor of modern mammals, but was mutated independently in the guinea pig and primate lineages. Max notes:
"Molecular geneticists who examine DNA sequences from an evolutionary perspective know that large gene deletions are rare, so scientists expected that non-functional mutant LGGLO gene copies--known as 'pseudogenes'--might still be present in primates and guinea pigs as relics of the functional ancestral gene. In contrast, Creationists believe that humans and guinea pigs were each created independently of all other species and must have been 'designed' to function without LGGLO. If this is true, these two species would not be expected to carry a defective copy of the LGGLO gene."
As Max goes on to point out, LGGLO pseudogenes have indeed been found in both guinea pigs and humans (Nishikimi et al. J Biol Chem 267: 21967, 1992; Nishikimi et al. J BIol Chem 269:13685, 1994). Not only did both groups retain the LGGLO pseudogene as expected, but in each group the gene had been rendered nonfunctional by different mutations. Once again, evolution makes a prediction that passes the observational tests.
Another type of pseudogene originates when an existing functional gene or gene segment is duplicated. Transposable elements are often involved in such duplications. In both the human and chimpanzee genome, for example, the 21-Hydroxylase gene and part of a nearby gene called "compliment C4" has been duplicated. In both cases, the duplicate copy has been rendered nonfunctional by the same 8 base pair deletion (American Journal Human Genetics, 50:766-80, 1992). Humans, apes, and the Old World monkeys share a unitary pseudogene for the enzyme galactosyltransferase. Humans, Old and New World monkeys, Tarsiers and Lemurs all share a Eta Hemoglobin psuedogene (Patterson, p68).
A third type of pseudogene originates when a normal gene is transcribed into the form of mRNA, is edited by enzymes which remove the introns, splice the exons together, and attach a string of 100-200 adenosine bases to the end (the poly-A tail), and then is reinserted back into the genome at a random location via the action of reverse transcriptase. normal processing which all mRNA transcripts undergo before they leave the nucleus to be translated into proteins at ribosomes located in the cytoplasm. Occasionally, if RT happens to be present in the nucleus, one of these processed mRNA transcripts can be reinserted into a chromosome in DNA form. The resulting pseudogenes are called processed pseudogenes because they have all the sequence characteristics of genes which have been transcribed from the genome into mRNA form and processed by RNA-editing enzymes. These pseudogenes are typically not expressed as proteins, because the promoter region is not copied and inserted along with the processed gene. It is conceivable, however, that a pseudogene could on rare occasions be reinserted near a promoter region. A processed psuedogene for epsilon immunoglobulin is shared by humans, chimps, gorilla, orangutans, and the Old World monkeys (Max, 1998).
Reverse transciptase is the same enzyme which allows RNA-based retroviri such as HIV to copy their genetic information into the host cell DNA. Occasionally, a retrovirus will invade a germline cell, copy their own genome into the host genome, and are propagated to descendents of that cellular line. If that cell happens to be in the germ line of the organism, the offspring's genome will retain copies of the viral genome, in every cell. If that organism happens to be in a line which speciates, then the new species will inherit the retroviral sequence as well. Surprisingly, perhaps several percent or more of the human genome seems to consist of copies of these retroviral invaders. This seems a small amount, until we remember that as little as 5% of our genome actually codes for proteins.
Why, to take one example, do humans, chimps and gorillas share unique retroviral insertions (ERVs), again supporting the same, very specific historical scenario implied by the fossil evidence? For reference, Ed Max describes how ERVs are identified:
Infectious retroviruses were discovered as agents of human disease and have been intensively studied. They are the most complex of retroposing elements and may have evolved from simpler ones described above. All contain two identical non-coding Long Terminal Repeats (LTRs) at their ends as well as three genes known as gag, pol and env. These genes are encoded in the virus not by DNA but by RNA. The pol gene encodes reverse transcriptase, and may also encode additional enzymatic activities. The env gene encodes proteins that coat the outside surface (envelope) of the infectious virus. The gag gene encodes additional proteins necessary for processing the viral components. The structure common to all retroviruses is thus LTR-gag-pol-env-LTR. The "left" LTR includes regulatory sequences that can initiate RNA transcription towards the right, into the gag-pol-env-LTR; the "left" LTR is then recopied from the "right" LTR by a complex mechanism. Infectious retroviruses include HTLVI, which causes a kind of leukemia in humans, and HIV, which causes AIDS. These viruses typically infect specific kinds of white blood cells--lymphocytes--and insert reverse-transcribed copies of their RNA genes into the DNA of these cells. Soon after the discovery of infections retroviruses, scientists noticed that similar sequences were present in the DNA of many mammalian species, including humans; these copies are called endogenous retroviruses, and presumably represent the consequences of ancient retroviral infections of germline cells. In human DNA there are about 8 different classes of endogenous retroviruses with members of each class varying in number from one or two to more than 50 copies. Essentially all of these endogenous retroviruses contain mutations that would disrupt the function of their genes, as would be expected if they inserted millions of years ago with no selective pressure to maintain the function of the genes. In addition, the duplicated LTR sequences represent potential targets for "homologous recombination" events that delete the DNA between the corresponding region of the LTRs, leaving only a single composite LTR sequence; many more copies of these isolated LTR fragments exist in the DNA than complete retroviral copies.
Plagiarized Errors and Molecular Genetics
These ERVs can tell us quite a bit about phylogeny. Since evolution is a branching process, we should expect ERVs to be distributed in specific ways. For instance, ERVs inserted recently will retain more sequence similarity across species, while those inserted longer ago will be found in more species, but share less sequence similarity. And so on. Johnson and Coffin write:
"Endogenous retrovirus loci provide no less than three sources of phylogenetic signal, which can be used in complementary fashion to obtain much more information than simple distance estimates of homologous sequences. First, the distribution of provirus-containing loci among taxa dates the insertion. Given the size of vertebrate genomes (>1 x 10^9) bp) and the random nature of retroviral integration, multiple integrations (and subsequent fixation) of ERV loci at precisely the same location are highly unlikely. Therefore, an ERV locus shared by two or more species is descended from a single integration event and is proof that the species share a common ancestor into whose germ line the original integration took place." (Johnson and Coffin 1999, p. 10255)
AND:
"The genetic distance between the 5' and 3' LTRs of an ERV reflects mutations accumulated since the time of integration and should therefore be proportional to the age of the provirus. HERV-KC4, HERV-KHML6.17, and RTVL-1a are found in both OWMs and hominoids, which are estimated to have last shared a common ancestor over 31 million years ago. By contrast, HERV-K18, RTVL-Ha, and RTVL-Hb are found only in humans, chimpanzees, and gorillas, which are thought to have diverged around 5 million years ago." (Johnson and Coffin 1999, p. 10259)
Constructing primate phylogenies from ancient retrovirus sequences
Conclusion
The hypothesis that humans are related to other species by a process of descent with modification provides the best explanation for the patterns seen in the homonid fossil record and for the genetic similarities and differences which exist between humans, primates, and other organisms, including the existence of shared pseudogenes and shared endogenous retroviral sequences. The alternative hypothesis offered by young-earth special-creationism, YESC, is not only inconsistent with the temporal and morphological distribution of human fossils, but is also incapable of generating any predictions with regard to the genetic similarities and differences which exist between humans and other organisms.
Works Cited
Cullen, Bryan. (1999). Proceedings of the National Academy of Sciences, Nov. 11.
Falk, Dean. (1992). Braindance. Henry Holt, New York.
Welkin E. Johnson and Jon M. Coffin. (1999). "Constructing Primate Phylogenies from Ancient Retrovirus Sequences," Proc. Natl. Acad. Sci., 10254-10260, p. 10255
Max, Edward, M.D., Ph.D. (1999). Plagiarized Errors and Molecular Genetics. Originally published in Creation/Evolution, 1986 (XIX, p.34).
Patterson, Colin.. (1999). Evolution: Second Edition. Cornell University Press, New York.
Some abstracts:
Biomechanical sppraisal of some skeletal features associated with head balance and posture in the Hominoidea. Acta Anat (Basel) 1975;92(4):580-4. Adams LM, Moore WJ.
The condylar position index, condylar angle and the area of insertion of the nuchal musculature corrected for condylar position, direction of muscle pull and skull size were determined in Homo sapiens, Gorilla, Pan and the casts of two Neanderthal and two australopithecine crania. In all three attributes, the values of H. sapiens exceeded, by statistically significant amounts, those for the ape genera. The greater value for the condylar position index indicates a better balance of the head, that for the condylar angle reflects the more vertical orientation of the vertebral column while that for the corrected nuchal area suggests a less effective nuchal musculature in H. sapiens as compared to the apes. In the casts of the Neanderthal crania, the values all came within, or close to, the ranges for H. sapiens. In the australopithecine casts, the condylar angle cane within the ranges for H. sapiens while the condylar position index and the corrected nuchal area were intermediate in value between H. sapiens and the extant apes. These findings are consistent with observations from other skeletal regions that while Australopithecus had an upright posture, this creature was, in the complex of posturally significant morphological features, distinct from extant hominoids.
Hallucal tarsometatarsal joint in Australopithecus afarensis. Latimer B, Lovejoy CO. Am J Phys Anthropol 1990 Jun;82(2):125-33
Hallucal tarsometatarsal joints from African pongids, modern humans, and Australopithecus afarensis are compared to investigate the anatomical and mechanical changes that accompanied the transition to terrestrial bipedality. Features analyzed include the articular orientation of the medial cuneiform, curvature of the distal articular surface of the medial cuneiform, and the articular configuration of the hallucal metatarsal proximal joint surface. Morphological characteristics of the hallucal tarsometatarsal joint unequivocally segregate quadrupedal pongids and bipedal hominids.
Talocrural joint in African hominoids: implications for Australopithecus afarensis. Am J Phys Anthropol 1987 Oct;74(2):155-75. Latimer B, Ohman JC, Lovejoy CO.
Talocrural joints of the African apes, modern humans, and A.L.288-1 are compared in order to investigate ankle function in the Hadar hominids. Comparisons between the hominids and African pongids clearly illustrate the anatomical and mechanical changes that occurred in this joint as a consequence of the evolutionary transition to habitual bipedality. Features which are considered include the obliquity of the distal tibial articular surface, the shape of the talar trochlea, and the location and functional implications of the talocrural axis. In every functionally significant feature examined the A.L.288-1 talocrural joint is fully bipedal. Moreover, the Hadar ankle complex also shows the functional constraints which are necessarily imposed by the adaptation to habitual bipedalism.
The mechanical effectiveness of erect and "bent-hip, bent-knee" bipedal walking in Australopithecus afarensis.Crompton RH, Yu L, Weijie W, Gunther M, Savage R.J Hum Evol 1998 Jul;35(1):55-74.
It is universally accepted that the postcranial skeleton of the early hominid Australopithecus afarensis shows adaptations, or at least exaptations, towards bipedalism. However, there continues to be a debate concerning the likely form of terrestrial bipedality: whether gait was erect, like our own, or "bent-hip, bent-knee" like the bipedalism of living chimpanzees. In this study we use predictive dynamic modelling to assess the mechanical effectiveness of AL-288-1 under both hypotheses, on the basis of data on segment proportions from the literature. AL-288-1's proportions are incompatible with the kinematics of chimpanzee bipedalism, but compatible with the kinematics of either erect or "bent-hip, bent-knee" human gait. In the latter case, neither the ankle nor the knee joint would have contributed substantial mechanical work to propulsion of the body, and net energy absorption is predicted for these joints, which would have resulted in increased heat load. Such an ineffective gait is unlikely to have lead to selection for "bipedal" features in the postcranial skeleton.
Metatarsophalangeal joints of Australopithecus afarensis. Latimer B, Lovejoy CO. Am J Phys Anthropol 1990 Sep;83(1):13-23
Metatarsophalangeal joints from African pongids, modern humans, and Australopithecus afarensis are compared to investigate the anatomical and mechanical changes that accompanied the transition to terrestrial bipedality. Features analyzed include the shape and orientation of the metatarsal heads, excursion of the metatarsophalangeal joints, and orientation of the basal articular surface of the proximal phalanges. These features unequivocally segregate quadrupedal pongids and bipedal hominids and demonstrate a clear adaptation to terrestrial bipedality in the Hadar pedal skeleton.
Latimer B, Lovejoy, CO. Am J Phys Anthropol 1989 Mar;78(3):369-86 The calcaneus of Australopithecus afarensis and its implications for the evolution of bipedality.
Calcanei from African apes, modern humans, and Australopithecus afarensis are compared to investigate the anatomical and mechanical changes that occurred in this bone as a result of the transition to terrestrial bipedality. Features analyzed include the cross-sectional area and volume of the calcaneal tuber, the geometry and orientation of the articular surfaces, and the surface topography of the calcaneal corpus. Calcaneal morphology is unequivocal in its partitioning of quadrupedal pongids and bipedal hominids.
Berge C, Kazmierczak JB. Effects
of size and locomotor adaptations on the hominid pelvis:
evaluation of australopithecine bipedality with a new
multivariate method. Folia Primatol (Basel) 1986;46(4):185-204.
Three pelves and eight innominate bones belonging to the fossil
species, Australopithecus africanus, Australopithecus robustus,
Homo erectus, and Homo sapiens, have been studied biometrically
and compared with those of recent humans and apes. A new method
of logarithmic factorial analysis suppresses both the size
effects and the size reference on pelvic proportions. In
combination with principal component analysis it allows
specializations to be dissociated from allometrical variations.
Some morphological differences on the hominid pelvis prove to be
mainly allometric. However, the pelvic morphology of
australopithecines is clearly differentiated from that of the
genus Homo (including H. erectus, OH 28, KNMER 3227). A.
africanus (Sts 14, MLD 7, AL 288) is nearer the humans than is A.
robustus (SK 50, SK 3155), which appears to be more specialized
in the australopithecine lineage. The pelvic morphology of A.
africanus, as integrated with the articular pelvic-femoral link,
appears to be biometrically equivalent to that of humans.
Wood B. Human evolution. Bioessays
1996 Dec;18(12):945-54
The common ancestor of modern humans and the great apes is
estimated to have lived between 5 and 8 Myrs ago, but the
earliest evidence in the human, or hominid, fossil record is
Ardipithecus ramidus, from a 4.5 Myr Ethiopian site. This genus
was succeeded by Australopithecus, within which four species are
presently recognised. All combine a relatively primitive
postcranial skeleton, a dentition with expanded chewing teeth and
a small brain. The most primitive species in our own genus, Homo
habilis and Homo rudolfensis, are little advanced over the
australopithecines and with hindsight their inclusion in Homo may
not be appropriate. The first species to share a substantial
number of features with later Homo is Homo ergaster, or 'early
African Homo erectus', which appears in the fossil record around
2.0 Myr. Outside Africa, fossil hominids appear as Homo erectus-like
hominids, in mainland Asia and in Indonesia close to 2 Myr ago;
the earliest good evidence of 'archaic Homo' in Europe is dated
at between 600-700 Kyr before the present. Anatomically modern
human, or Homo sapiens, fossils are seen first in the fossil
record in Africa around 150 Kyr ago. Taken together with
molecular evidence on the extent of DNA variation, this suggests
that the transition from 'archaic' to 'modern' Homo may have
taken place in Africa.
Henry M. McHenry and Katherine Coffing. AUSTRALOPITHECUS TO HOMO: Transformations in Body and Mind. Annu. Rev. Anthropol. 2000. 29:125-146.
Significant changes occurred in human evolution between 2.5 and 1.8 million years ago. Stone tools first appeared, brains expanded, bodies enlarged, sexual dimorphism in body size decreased, limb proportions changed, cheek teeth reduced in size, and crania began to share more unique features with later Homo. Although the two earliest species of Homo, H. habilis and H. rudolfensis, retained many primitive features in common with australopithecine species, they both shared key unique features with later species of Homo. Two of the most conspicuous shared derived characters were the sizes of the brain and masticatory apparatus relative to body weight. Despite the shared derived characters of H. habilis and H. rudolfensis, one unexpected complication in the transition from australopithecine to Homo was that the postcranial anatomy of H. habilis retained many australopithecine characteristics. H. rudolfensis, however, seems to have had a more human-like body plan, similar to later species of Homo. H. rudolfensis may therefore represent a link between Australopithecus and Homo.
Ruvolo M. Molecular phylogeny of
the hominoids: inferences from multiple independent DNA sequence
data sets. Mol Biol Evol 1997 Mar;14(3):248-65
Consensus on the evolutionary relationships of humans,
chimpanzees, and gorillas has not been reached, despite the
existence of a number of DNA sequence data sets relating to the
phylogeny, partly because not all gene trees from these data sets
agree. However, given the well-known phenomenon of gene tree-species
tree mismatch, agreement among gene trees is not expected. A
majority of gene trees from available DNA sequence data support
one hypothesis, but is this evidence sufficient for statistical
confidence in the majority hypothesis? All available DNA sequence
data sets showing phylogenetic resolution among the hominoids are
grouped according to genetic linkage of their corresponding genes
to form independent data sets. Of the 14 independent data sets
defined in this way, 11 support a human-chimpanzee clade, 2
support a chimpanzee-gorilla clade, and one supports a human-gorilla
clade. The hypothesis of a trichotomous speciation event leading
to Homo; Pan, and Gorilla can be firmly rejected on the basis of
this data set distribution. The multiple-locus test (Wu 1991),
which evaluates hypotheses using gene tree-species tree mismatch
probabilities in a likelihood ratio test, favors the phylogeny
with a Homo-Pan clade and rejects the other alternatives with a P
value of 0.002. When the probabilities are modified to reflect
effective population size differences among different types of
genetic loci, the observed data set distribution is even more
likely under the Homo-Pan clade hypothesis. Maximum-likelihood
estimates for the time between successive hominoid divergences
are in the range of 300,000-2,800,000 years, based on a
reasonable range of estimates for long-term hominoid effective
population size and for generation time. The implication of the
multiple-locus test is that existing DNA sequence data sets
provide overwhelming and sufficient support for a human-chimpanzee
clade: no additional DNA data sets need to be generated for the
purpose of estimating hominoid phylogeny. Because DNA
hybridization evidence (Caccone and Powell 1989) also supports a
Homo-Pan clade, the problem of hominoid phylogeny can be
confidently considered solved.
McCollum MA. Subnasal
morphological variation in fossil hominids: a reassessment based
on new observations and recent developmental findings. Am J
Phys Anthropol 2000 Jun;112(2):275-83
Quantitative and qualitative assessments of subnasal morphology
in fossil hominids yield distinct patterns which have been used
both to sort robust from nonrobust australopithecine taxa and to
distinguish individual species. Recently, new developmental
models have been applied to hominoid subnasal morphological
variation. These studies require that certain features of the
fossil hominid subnasal region, in particular the topography of
the nasal cavity entrance and details of vomeral morphology, be
reevaluated. This study does so for the robust and nonrobust
australopithecines, early Homo (H. habilis/H. rudolfensis), and
African H. erectus. Results reaffirm an overall similarity of the
nonrobust Australopithecus subnasal morphological pattern with
that of the chimpanzee. They further indicate that a vomeral
insertion above the nasal surface of the premaxilla should be
added to the list of traits characteristic of the robust
australopithecine subnasal morphological pattern. Finally,
reassessment of subnasal morphology in the early Homo and H.
erectus samples from Africa suggest that these two taxa share a
similar subnasal morphological pattern. This pattern consists of
a smooth nasal cavity entrance, a horizontal nasal sill whose
anterior edge is demarcated by a strong nasal crest, and a well-developed
horizontal spine at the posterior edge of the nasal sill.
Although none of the African fossil Homo specimens preserve a
vomer, indirect evidence suggests that it would have inserted
above the nasal sill.
Wood B, Aiello LC. Taxonomic and
functional implications of mandibular scaling in early hominins. Am
J Phys Anthropol 1998 Apr;105(4):523-38
Body mass estimates for fossil hominin taxa can be obtained from
suitable postcranial and cranial variables. However, the nature
of the taphonomic processes that winnow the mammalian fossil
record are such that these data are usually only available for
the minority of the specimens that comprise the hypodigm of a
species. This study has investigated the link between species
mean body mass and the height and width of the mandibular corpus
in a core sample of 23 species of extant simians. The slopes of
the least-squares regressions for the whole sample and for the
hominoid subset are similar. However, the intercepts differ so
that for a given body mass, a hominoid will generally have a
smaller mandible than a generalized simian. The same mandibular
measurements were taken on 75 early hominin mandibles assigned to
eight species groups. When mandibular corpus height- and width-derived
estimates of body mass for the fossil taxa were compared with
available postcranial and cranial-derived body mass estimates,
the eight early hominin species sort into four groups. The first,
which includes A. afarensis and A. africanus, has mandibles which
follow a "generalized simian" scaling relationship. The
second group, which comprises the two "robust"
australopithecine species, P. boisei and P. robustus, has
mandibles which scale with body mass as if they are "super-simians,"
for they have substantially larger mandibles than a simian with
the same body mass. The two "early Homo" species, H.
habilis sensu stricto and H. rudolfensis, make up the third group.
It has mandibular scaling relationships that are intermediate
between that of the comparative simian sample and that of the
hominoid subsample. The last of the four groups comprises H.
ergaster and H. erectus; their mandibles scale with body mass as
if they were hominoids, so that of the four groups they have the
smallest mandibles per unit body mass. These results are related
to comparable information about relative tooth size. Their
relevance for attempts to interpret the dietary adaptations of
early hominins are explored.