Contents
1 Introduction
*1.1 The nitrogen cycle
*1.1.1 Symbiotic Nitrogen fixation
*1.2 Nitrogen in Agriculture
*1.2.1 Nitrate, ammonium and SON
*1.2.2 Legumes as a source of N for agriculture
*1.2.3 Nitrogen Transfer from legumes to non-legumes
*1.2.4 Synthetic N fertiliser
*1.2.5 Organic Agriculture
*1.2.6 Energy efficiency of fertiliser N and legume-based systems
*1.2.7 Economic benefits of legume-based farming
*1.2.8 Environmental benefits of legume-based farming
*1.3 White clover
*1.3.1 A description of the plant
*1.3.2 A history of White clover cultivation
*1.4 Methods of estimation of Nitrogen fixation
*1.4.1 Nitrogen benefit, yield and total nitrogen difference methods
*1.4.2 Acetylene reduction
*1.4.3 Natural Abundance
*1.4.4
15N Dilution *1.4.5 Xylem-solute techniques
*1.4.6
15N2 Incubation method *1.4.7 Summary
*1.5 Estimates of N fixation by white clover
*1.6 Effect of soil nitrogen on N fixation
*1.7 Effects of grazing and cutting on N fixation in grassland
*1.7.1 Effects of dung and urine deposition
*1.7.2 Effects of grassland management on N fixation
*1.7.3 Effect of companion species
*1.7.4 Temporal variation of N fixation
*1.7.5 Effect of temperature on N fixation
*1.8 Other factors affecting N fixation
*1.9 Summary
*2 Aims of study
*2.1 Specific objectives
*2.1.1 Cyclical Replacement
*3 N cycle in grass/white clover leys
*3.1 Experimental sites
*3.1.1 The Tulloch Organic Unit
*3.1.2 The Aldroughty Organic Unit
*4 N fixation and grass and clover dynamics
*4.1 Aims
*4.2 Hypotheses
*4.3 Methods
*4.3.1 N fixation using 15N dilution technique
*4.3.2 Flowering rate
*4.3.3 Tap-roots
*4.4 Calculations
*4.4.1 pNdfa
*4.4.2 N fixation
*4.4.3 N transfer
*4.4.4 Soil derived N
*4.5 Results
*4.5.1 Nitrogen fixation
*Tulloch
*Aldroughty
*4.5.2 Proportion of nitrogen derived from the atmosphere
*Tulloch
*4.5.3 Nitrogen concentration in clover herbage
*Tulloch
*Aldroughty
*4.5.4 Dry matter yield of clover
*Tulloch
*Aldroughty
*4.5.5 Soil derived N in grass and clover
*Tulloch
*Aldroughty
*4.5.6 Nitrogen concentration of grass
*Tulloch
*Aldroughty
*4.5.7 Dry matter yield of grass
*Aldroughty
*4.5.8 N transfer
*4.5.9 Plant physiology
*4.5.10 Tap-roots
*4.5.11 Nitrogen budgets
*4.6 Discussion
*4.6.1 Annual variation
*4.6.2 Within-season variation
*4.6.3 Age of ley related variation
*4.6.4 Effects of cutting and grazing on clover and grass
*4.6.5 Effects of rotation type
*4.6.6 Variation between sites
*4.6.7 Variation within sites
*4.6.8 N transfer
*5 Soil chemistry of grass-clover swards
*5.1 Aims
*5.2 Hypotheses
*5.3 Methods
*5.3.1 Soil sampling and analysis
*5.3.2 Calculations
*5.3.3 Sampling dates
*5.3.4 Statistical analysis
*5.4 Results
*5.4.1 Soil Nitrate-N
*5.4.2 Soil ammonium-N
*5.4.3 Soluble Organic Nitrogen-N
*5.4.4 Total soluble N
*5.4.5 P, K and Mg
*5.5 Discussion
*5.5.1 Hypothesis a)
*5.5.2 Hypothesis b)
*5.5.3 Hypothesis c)
*5.5.4 Hypothesis d)
*5.5.5 Total and Soluble N in organic soils
*5.5.6 Annual variation of soil nutrients
*5.5.7 Seasonal variation of soil nutrients
*5.5.8 Effects of management on soil nutrients
*5.6 Discussion of grass white clover leys
*5.6.1 Comparison of soil N and N fixation in 1999 and 2000
*5.6.2 Grass and Clover 1997-2000
*5.6.3 Comparison of soil N and plant behaviour in different ages of ley
*5.6.4 Seasonal variation of soil N and grass and clover dynamics
*5.6.5 Effects of rotation type on soil chemistry and plant behaviour
*5.6.6 Other soil properties and plant behaviour
*5.6.7 Final points
*6 Effects of clover and grass on soil nitrogen
*6.1 Introduction
*6.2 Hypotheses
*6.3 Methods
*6.3.1 Calculations
*6.4 Results
*6.4.1 Dry matter of grass and clover
*6.4.2 Soil N
*6.5 Discussion
*7 Colonisation of different soil microsites by clover and ryegrass (invasion experiment)
*7.1 Introduction
*7.2 Hypotheses
*7.3 Methods
*7.3.1 Treatments
*7.4 Results
*7.4.1 Invasions
*7.4.2 Yields of non-invading clover and grass
*7.5 Discussion
*7.5.1 Hypothesis a)
*7.5.2 Hypothesis b)
*7.5.3 Hypothesis c
*7.5.4 Hypothesis d)
*8 Discussion
*8.1 Hypothesis 1
*8.2 Hypothesis 2
*8.3 Hypothesis 3
*8.4 Hypothesis 4
*8.5 Hypothesis 5
*8.6 Hypothesis 6
*8.7 Hypothesis 7
*8.8 Hypothesis 8
*8.9 The cyclical replacement hypothesis
*8.10 Nitrogen fixation and fertiliser use in Scottish Agriculture
*8.10.1 Measuring N fixation
*8.11 Soil N at Tulloch
*8.12 Morphology of clover
*8.13 Conclusions
*
Figures
Figure 1.1.1 The Nitrogen Cycle
*Figure 1.6.1 Effect of N fertiliser applications on N fixation by white clover-grass leys.
*Figure 3.1.1 Layout of the trial rotation plots at Tulloch in 2000.
*Figure 3.1.2 Average monthly soil temperature and total monthly rainfall in 1997.
*Figure 3.1.3 Average monthly soil temperature and total monthly rainfall in 1998
*Figure 3.1.4 Average monthly soil temperature and total monthly rainfall in 1999
*Figure 3.1.5 Average monthly soil temperature and total monthly rainfall in 2000
*Figure 4.5.1 Nitrogen fixation plotted against clover dry matter yield in 1-year-old leys at Tulloch in 1997. Each data point represents data from one subplot.
*Figure 4.5.2 Nitrogen fixation plotted against clover dry matter yield in 1- and 2-year-old leys at Tulloch in 1998. Each data point represents data from one subplot.
*Figure 4.5.3 Nitrogen fixation plotted against clover dry matter yield in 1-, 2- and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 1999. Each data point represents data from one subplot.
*Figure 4.5.4 Nitrogen fixation plotted against clover dry matter yield in 1-, 2-, 3- and 4-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 2000. Each data point represents data from one subplot.
*Figure 4.5.5 Nitrogen fixation excluding N transfer at Tulloch in 1-year-old leys in 1997. Values represent the accumulated N fixed during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.6 Nitrogen fixation excluding N transfer at Tulloch in 1998, 1- and 2-year-old leys. Values represent the accumulated N fixed during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.7 Nitrogen fixation excluding N transfer in 1999, 1-, 2- and 3-year-old leys (Tulloch) and 1-year-old leys (Aldroughty). Values represent the accumulated N fixed during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.8 Nitrogen fixation excluding N transfer in 2000, 1-, 2-, 3- and 4-year-old leys (Tulloch) and 1-year-old leys (Aldroughty). Values represent the accumulated N fixed during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.9 N fixation excluding transfer in 3-year-old leys in the 66% ley rotation and the 50% ley rotation in 2000. Values represent the accumulated N fixed during the month of sampling (mean of 6 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.10 Proportion of nitrogen in clover herbage derived from atmosphere, excluding transfer (pNdfaET) in 1-year-old leys at Tulloch in 1997. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.11 Proportion of nitrogen in clover herbage derived from atmosphere, excluding transfer (pNdfaET) in 1- and 2-year-old leys at Tulloch in 1998. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.12 Proportion of nitrogen in clover herbage derived from atmosphere, excluding transfer (pNdfaET) in 1,2 and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 1999. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.13 Proportion of nitrogen in clover herbage derived from atmosphere, excluding transfer (pNdfaET) in 1-, 2-, 3- and 4-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 2000. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.14 Concentration of nitrogen in clover herbage expressed as a percentage of dry matter. 1-year-old leys at Tulloch, in 1997. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.15 Concentration of nitrogen in clover herbage expressed as a percentage of dry matter. 1- and 2-year-old leys at Tulloch, in 1998. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.16 Concentration of nitrogen in clover herbage expressed as a percentage of dry matter. 1-, 2- and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty, in 1999. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.17 Concentration of nitrogen in clover herbage expressed as a percentage of dry matter. 1-, 2-, 3- and 4-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 2000. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.18 Clover herbage dry matter yield at Tulloch in 1-year-old leys in 1997. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.19 Clover herbage dry matter yield at Tulloch in 1- and 2-year-old leys in 1998. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.20 Clover herbage dry matter yield in 1-, 2- and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 1999. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.21 Clover herbage dry matter yield in 1-, 2-, 3- and 4-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 2000. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.22 Percentage ground cover of clover in autumn in successive years since the establishment of the trial. Each point is a mean of four samples (SAC data)
*Figure 4.5.23 Soil derived N in grass and clover herbage in 1-year-old leys at Tulloch in 1997. Values represent the total accumulation of non-fixed N during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.24 Soil derived N in grass and clover herbage in 1- and 2-year-old leys at Tulloch in 1998. Values represent the total accumulation of non-fixed N during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.25 Soil derived N in grass and clover herbage in 1-, 2- and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 1999. Values represent the total accumulation of non-fixed N during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.26 Soil derived N in grass and clover herbage in 1-, 2-, 3- and 4-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 2000. Values represent the total accumulation of non-fixed N during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.27 Soil derived N in grass and clover herbage (kg N ha-1) in leys in the 66% ley rotation and the 50% ley rotation. Columns represent total soil derived N over the growing season (mean of 6 subplots). Bars represent standard errors.
*Figure 4.5.28 Concentration of nitrogen in grass herbage expressed as a percentage of dry matter. 1-year-old leys at Tulloch, in 1997. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.29 Concentration of nitrogen in grass herbage expressed as a percentage of dry matter. 1- and 2-year-old leys at Tulloch, in 1998. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.30 Concentration of nitrogen in grass herbage expressed as a percentage of dry matter. 1-, 2- and 3-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 1999. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.31 Concentration of nitrogen in grass herbage expressed as a percentage of dry matter. 1-, 2-, 3- and 4-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 2000. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.32 Grass herbage dry matter yield at Tulloch in 1-year-old leys in 1997. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.33 Grass herbage dry matter yield at Tulloch in 1- and 2-year-old leys in 1998. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.34 Grass herbage dry matter yield at Tulloch in 1-, 2- and 3-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 1999. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.35 Grass herbage dry matter yield at Tulloch in 1-, 2-, 3- and 4-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 2000. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
*Figure 4.5.36 Grass dry matter yields in the 66% and 50% ley rotations in 2000. Columns represent total grass dry matter yield for the sampling period (mean of 6 subplots). Bars represent standard errors.
*Figure 4.5.37 Number of flowers per g dried clover in samples taken for 15N analysis in 2001. Columns represent the mean of 12 subplots. Bars represent standard errors.
*Figure 4.5.38 Dry matter content of clover herbage in 2001, expressed as a percentage of fresh weight. Columns represent average dry matter content for the whole year (mean of 4 subplots).
*Figure 4.5.39 Number of tap-roots present in 8 soil cores from clover microsites in August 2001. Columns represent the mean of four samples. Bars represent standard errors.
*Figure 5.4.1 Nitrate-N in soil under grass and clover microsites in all ley plots at Tulloch, between February 1999 and August 2001. Points represent means of 14 samples. Bars represent standard errors.
*Figure 5.4.2 Nitrate-N in soil in 1-, 2-, 3- and 4-year-old leys at Tulloch between January 1999 and August 2001. Columns represent means of 8 samples (1-, 2- and 3-year-old leys) and 4 samples (4-year-old leys). Bars represent standard errors.
*Figure 5.4.3 Ammonium-N in soil in 1-, 2-, 3- and 4-year-old leys at Tulloch between January 1999 and August 2001. Columns represent means of 8 samples (1-, 2- and 3-year-old leys) and 4 samples (4-year-old leys). Bars represent standard errors.
*Figure 5.4.4 Ammonium-N in the 50% and 66% ley rotations at Tulloch (1–3-year-old leys only). Columns represent means of values for 12 plots. Bars represent standard errors.
*Figure 5.4.5 Soluble organic nitrogen under grass and clover microsites in all ley plots at Tulloch, between February 1999 and August 2001. Points represent means of 14 samples. Bars represent standard errors.
*Figure 5.4.6 Total Soluble N under grass and clover microsites in all ley plots at Tulloch, between February 1999 and August 2001. Points represent means of 14 samples. Bars represent standard errors.
*Figure 5.4.7 Total Soluble N in soil in 1-, 2-, 3- and 4-year-old leys at Tulloch between January 1999 and August 2001. Columns represent means of 8 samples (1-, 2- and 3-year-old leys) and 4 samples (4-year-old leys). Bars represent standard errors.
*Figure 5.4.8 Potassium levels in soil from under grass and clover microsites in 1-, 2-, 3- and 4-year-old leys in August 2001. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors.
*Figure 5.5.1 Percentage organic matter in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
*Figure 5.5.2 Phosphorus levels in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
*Figure 5.5.3 Potassium levels in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
*Figure 5.5.4 Magnesium levels in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
*Figure 5.5.5 pH levels in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
*Figure 5.6.1 Proportion of nitrogen derived from the atmosphere (average over whole year, mean of three subplots) and total soluble nitrogen (values for April) for 1999 and 2000. Each point represents one plot
*Figure 5.6.2 Correlation between proportion of clover N derived from the atmosphere (pNdfa) and grass N yield. Each point represents one subplot and sampling date.
*Figure 5.6.3 Correlation between N concentration of clover (%) and proportion of clover N derived from the atmosphere (pNdfa). Each point represents one subplot and sampling date.
*Figure 6.4.1 Dry matter yields of grass and clover in main root exudation experiment. Columns represent means of 8 replicates. Bars represent standard errors.
*Figure 6.4.2 Nitrate-N levels in soil from all treatments in the main root exudation experiment. Columns indicate means of 8 replicates. Bars indicate standard errors.
*Figure 6.4.3 Ammonium-N levels in soil from all treatments in the main root exudation experiment. Columns indicate means of 8 replicates. Bars indicate standard errors.
*Figure 6.4.4 TSN levels in soil from all treatments in the main root exudation experiment. Columns indicate means of 8 replicates. Bars indicate standard errors.
*Figure 6.4.5 SON levels in soil from all treatments in the main root exudation experiment. Columns indicate means of 8 replicates. Bars indicate standard errors.
*Figure 7.3.1 Treatments in the invasion experiments. Each circle represents one of the replicates of experiment A, B, C or D. Letters in bold are codes for experiments A, B, C and D.
*Figure 7.4.1 Yield of invading clover on all four sampling dates AG1 = experiment A, grass, unfertilised; AG2 = experiment A, grass + urine; AB1 = experiment A, bare soil, unfertilised; AB2=experiment A, bare soil + urine; BB1=experiment B, bare soil, unfertilised; BB2 = experiment B, bare soil + urine; BG1 = experiment B, grass, unfertilised; CGH1 = experiment C, grass, harvested, unfertilised; CGH3 = experiment C, grass, harvested + 5 g N m-2; CGU1 = experiment C, grass, unharvested, unfertilised; CGU3 = experiment C grass, unharvested + 5 g N m-2; DG1 = experiment D, grass, unharvested, unfertilised.
*Figure 7.4.2 Yield of invading grass on all four sampling dates. AC1 = experiment A, clover, unfertilised; AB1 = experiment A, bare soil, unfertilised; AB2 = experiment A, bare soil + urine; BC1 = experiment B, clover, unfertilised; BC2 = experiment B, clover + urine; BB1 = experiment B, bare soil, unfertilised; BB2 = experiment B, bare soil + urine; CC1 = experiment C, clover, unfertilised; DCH1 = experiment D, clover, harvested, unfertilised; DCH3 = experiment D, clover, harvested + 5 g N m-2; DCU1 = experiment D, clover, unharvested, unfertilised; DCU3 = experiment D clover, unharvested + 5 g N m-2.
*Figure 7.4.3 Yield of clover in microsites initially planted with clover on all four sampling dates AC1 = experiment A, clover, unfertilised; BC1 = experiment B, clover unfertilised; BC2 = experiment B, clover + urine; CC1 = experiment C, clover, unfertilised; DCH1 = experiment D, clover harvested, unfertilised; DCH3 = experiment D, clover, harvested + 5 g N m-2; DCU1 = experiment D, clover, unharvested, unfertilised; DCU3 = experiment D clover, unharvested + 5 g N m-2.
*Figure 7.4.4 Yield of grass in microsites initially planted with grass on all four sampling dates. AG1 = experiment A, grass, unfertilised; AG2 = experiment A, grass + urine; BG1 = experiment B, grass, unfertilised; CGH1 = experiment C, grass, harvested, unfertilised; CGH3 = experiment C, grass, harvested + 5 g N m-2; CGU1 = experiment C, grass, unharvested, unfertilised; CGU3 = experiment C grass, unharvested + 5 g N m-2; DG1 = experiment D, grass, unharvested, unfertilised.
*Figure 7.4.5 Number of flowerheads of invading clover on 13.8.01 AG1 = experiment A, grass, unfertilised; AG2 = experiment A, grass + urine; AB1 = experiment A, bare soil, unfertilised; AB2 = experiment A, bare soil + urine; BB1=experiment B, bare soil, unfertilised; BB2 = experiment B, bare soil + urine; BG1 = experiment B, grass, unfertilised; CGH1 = experiment C, grass, harvested, unfertilised; CGH3 = experiment C, grass, harvested + 5 g N m-2; CGU1 = experiment C, grass, unharvested, unfertilised; CGU3 = experiment C grass, unharvested + 5 g N m-2; DG1 = experiment D, grass, unharvested, unfertilised.
*Figure 7.4.6 Number of flowerheads of clover in microsites initially planted with clover on 13.8.01. AC1= experiment A, clover, unfertilised; BC1 = experiment B, clover, unfertilised; BC2 = experiment B, clover + urine; CC1 = experiment C, clover, unfertilised; DCH1 = experiment D, clover, harvested, unfertilised; DCH3 = experiment D, clover, harvested + 5 g N m-2; DCU1 = experiment D, clover, unharvested, unfertilised; DCU3 = experiment D, clover, unharvested + 5 g N m-2.
*Figure 7.4.7 Yields of grass and invading clover, experiment A. 13.8.01. Each point represents the grass and clover yields for one of the microsites initially planted with grass (treatments AG1 and AG2).
*Figure 8.12.1 Correlation between average annual pNdfa x (1/average annual clover N concentration) and total N fixation in the following year. Each point is the mean of three subplots in a plot.
*
Tables
Table 1.4.1 Methods of measuring N fixation
*Table 1.5.1 N fixation by white clover in different parts of the world and at different levels of N fixation
*Table 1.8.1 Factors reducing nitrogen fixation in white clover leys
*Table 3.1.1 Cropping sequences for the 66% ley rotation and the 50% ley rotation at Tulloch. u/s = undersown with grass and clover
*Table 3.1.2 Dates of silage cuts on leys at Tulloch, 1997-2000
*Table 3.1.3 Manure applications to cut leys at Tulloch 1997-2000
*Table 3.1.4 Cropping sequences for the 66% ley rotation and the 50% ley rotation at Aldroughty. u/s = undersown with grass and clover
*Table 4.3.1 Sampling dates for 15N dilution study of leys of different ages at Tulloch 1997-2000
*Table 4.3.2 Sampling dates for 15N dilution study of leys of different ages at Tulloch 1997-2000
*Table 4.5.1 N transfer at Tulloch (kg ha-1) between 1997 and 2000, calculated by isotope methods a and b, and the N difference method. Numbers in brackets represent standard errors.
*Table 4.5.2 Correlations between N transfer estimates at Tulloch, calculated by isotope methods a) and b) and the N difference method. Data from plots 13, 16, 17 and 18 in 2000.
*Table 4.5.3 N transfer calculated by isotope methods a) and b) and the N difference method at Aldroughty (kg ha-1). Numbers in brackets represent standard errors.
*Table 4.5.4 Number of clover flowers produced per m2 within the subplots used for the 15N dilution study. Numbers in brackets represent standard errors.
*Table 4.5.5 N budgets in all years and ages of ley, showing fixed and soil derived N present in all grass and clover mixtures and monocultures. Data from all plots.
*Table 4.5.6 N budget for plots 13 (4-year-old ley), 16 (1-year-old ley), 17 (2-year-old ley) and 18 (3-year-old ley) showing fixed and soil derived N present in all grass and clover mixtures and monocultures.
*Table 5.6.1Correlations between soil and plant factors at Tulloch 1997-2000 (ns= not significant, * = p<0.05, ** = p<0.01, *** = p<0.001)
*Table 6.3.1 Experimental treatments in the pot experiment to test effects of clover and grass on soil
*Table 7.4.1 Experiment A: % of artificial microsites invaded by species alien to that microsite on four sampling dates and five treatments. AG1 = grass, untreated; AG2 = grass + artificial urine; AC1 = Clover, untreated; AB1 = bare soil, untreated; AB2 = bare soil + artificial urine.
*Table 7.4.2 Experiment B: % of artificial microsites invaded by species alien to that microsite on four sampling dates and five treatments. BC1 = clover, untreated; BC2 = clover + artificial urine; BG1 = grass, untreated; BB1 = bare soil, untreated; BB2 = bare soil + artificial urine.
*Table 7.4.3 Experiment C: % of artificial microsites invaded by species alien to that microsite on the final two sampling dates (after removal of the plastic barriers). CGH1 = grass, harvested, unfertilised; CGH3 = grass, harvested + 5 g N m-2; CGU1 = grass, unharvested; CGU3 = grass, unharvested + 5 g N m-2; CC1 = clover, harvested, unfertilised.
*Table 7.4.4 Experiment D: % of artificial microsites invaded by species alien to that microsite on the final two sampling dates (after removal of the plastic barriers). DCH1 = clover, harvested, unfertilised; DCH3 = clover, harvested + 5 g N m-2; DCU1 = clover, unharvested; DCU3 = clover, unharvested + 5 g N m-2; DG1 = clover, harvested, unfertilised.
*
Factors affecting Symbiotic Nitrogen Fixation in Organic Farming Systems
"That the spent earth may gather heart again,
And bettered by cessation, bear the grain.
At least where vetches, pulse and tares, have stood,
And stalks of lupines grew (a stubborn wood),
The ensuing season, in return, may bear
The bearded product of the golden year"
-Virgil’s Georgics Book I
Introduction The nitrogen cycle
After water, nitrogen (N) is the nutrient that most commonly limits plant growth (Vitousek and Howarth, 1991). Every time crops, meat, milk, eggs or wool are exported from a farm, the nitrogen contained within them must be replaced, as this nitrogen has come out of the soil of the farm, and if it is not replaced, soil fertility will suffer. For this reason, the supply of nitrogen is vital for successful agriculture.
In natural ecosystems, nitrogen passes through a cycle that involves living organisms, the soil and the atmosphere. Nitrogen gas (N2) is the main component of the atmosphere. N2 is largely inert, but it can be converted into more reactive N compounds by a variety of processes known collectively as N fixation. Certain types of bacteria can convert N2 into ammonium (NH4+) compounds. These N-fixing bacteria can be either free living or in close associations with plants (symbiosis). Of these the symbiotic bacteria present in the roots of leguminous plants are the most important, both in terms of the amounts of N that they fix globally, and in their importance in agriculture (Schlesinger, 1997). In most environments, N fixation by free living N fixing bacteria is low, usually less than
10 kg ha-1yr-1 (Lockyer and Cowling, 1977). However, free living N fixing bacteria can be important in some environments, especially where there is an abundant supply of energy rich organic matter, but little nitrogen. Relatively high levels of non-symbiotic nitrogen fixation often occur in decaying logs (Roskoski, 1980). In the broadbalk continuous wheat experiment at Rothamsted, N fixation by free living bacteria has been estimated at 28 kg N ha-1 year-1 (Witty, 1979). Some N fixing bacteria live symbiotically within the hindguts of arthropods, particularly those that feed on decaying wood and other low N diets e.g termites. In some environments, N fixation in arthropod guts may be as high as 10-40 kg N ha-1 year-1 (Nardi et al. 2002). Symbiotic N fixation will be discussed more fully in section 1.2. N2 can also be fixed by lightning, and industrially by the Haber process, which is used to manufacture fertiliser. The Haber process was developed during the First World War, originally in order to manufacture explosives. N is artificially fixed in the form of ammonia, by heating N2 at high pressure in the presence of a catalyst. The Haber process can be summarised by the following equations:
![]()
![]() [.2]
[.2]
(Schlesinger, 1997). In 2000, the world used approximately 80 million tonnes of fertiliser N, in 1960 the figure was nearer 11 million tonnes
(International Fertilizer Industry Association, 2000). Agricultural legumes currently fix approximately 40 million tonnes of nitrogen. In pre-industrial times, terrestrial N fixation was approximately 90-130 million tonnes per annum. Human impacts have therefore approximately doubled the inputs of N into the terrestrial N cycle (Vitousek et al., 1997). Burning of fossil fuels also causes a release of fixed N. Vehicle exhausts can also be a significant input of nitrogen in industrialised countries (Lee and Dollard, 1994). By these processes N passes from the atmosphere into the soil, water and living organisms. N is returned to the atmosphere from the soil by the actions of denitrifying bacteria, completing the cycle.The majority of organisms have no ability to fix nitrogen, and so they are dependent on a N cycle within the larger scale N cycle of fixation and denitrification, in which fixed nitrogen is continually recycled. N fixation only accounts for an estimated 12% of the nitrogen used annually by plants. The rest is obtained by recycling of nitrogen already present in plants and the soil. It has been estimated that the mean turnover time of nitrogen in terrestrial ecosystems, from fixation to denitrification, is around 700 years (Schlesinger, 1997). Within the soil, microbes convert NH4+ into nitrate (NO3) and nitrite (NO2), a process known as nitrification. Plants take up
NO3- and ammonium NH4+ from the soil nutrient solution and convert this into protein. Some of the plant protein is returned to the soil through death and decay of part or all of the plant. Some plants can utilise amino acids and complex organic N compounds as N sources in addition to nitrate and/or ammonium (e.g. Persson and Nasholm, 2001, Lipson and Nasholm, 2001). In the soil, bacteria convert the dead organic material into microbial biomass and soil organic matter, which includes a complex mixture of nitrogen compounds. Over time, soil organic nitrogenprotein is mineralised by bacteria into amino compounds, then ammonium and finally to nitrate (Bolger et al, 2001, Killpack and Bucholz, 1993). The speed at which this occurs depends upon the ratio of carbon to nitrogen in the soil organic matter. From measurements of the turnover of soil organic matter and inputs, it has been estimated that on average N remains in soil organic matter for over 100 years (Schlesinger, 1997).
When animals feed on plants, some of the plant protein N is converted into animal protein, but most is returned to the soil as dung and urine. Henzel and Ross (1973) compared N intakes and excretion of N by farm animals on diets with a range of N contents and concluded that beef cattle excreted 90-96%, sheep excreted 87-95% and dairy cattle excreted 72-87% of the N in their diet. Ultimately, when animals die, the N contained in their bodies decays and eventually becomes available to plants and microbes. Nitrogen from dung and urine is incorporated into the soil organic matter, and is eventually released into solution as ammonium, nitrate and nitrite. These basic principles apply as much to animals within the soil that feed on roots and decaying plant material, as they do to cows and sheep.
Some of the N in soil is lost to the atmosphere, either by volatilisation of ammonia, or denitrification by bacteria, in which nitrate is converted to nitrite, and then into nitric oxide (NO), nitrous oxide (N2O) and N2. The latter three compounds can escape to the atmosphere (Erickson et al. 2001). Denitrification takes place under anaerobic conditions, and is commonest in waterlogged soil, although it can take place in anaerobic pockets within most soils (Schlesinger 1997). Volatilisation losses of N are greatest in fertilised soils and during the decomposition of urea excreted by animals (Terman, 1979,
Fillery, 2001). Nitrogen is also lost from the soil when it is leached out of reach of the plant root systems, by rainfall. In natural ecosystems, leaching and gaseous losses are usually small (e.g. Erickson et al. 2001). A simplified overview of these processes is shown in Figure 1.1.1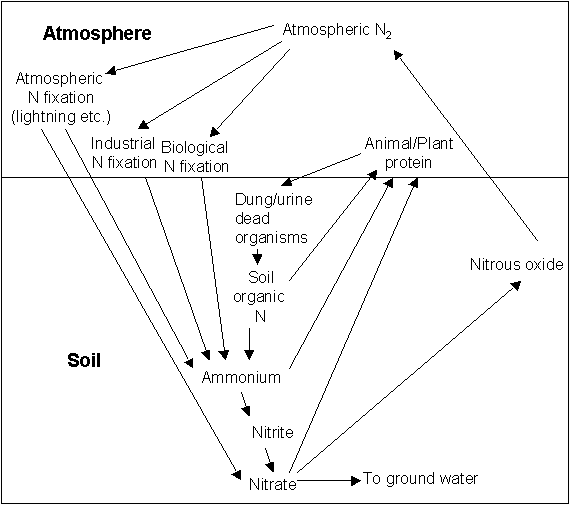
Figure .1 The Nitrogen Cycle
As we have seen, certain bacteria have the ability to fix atmospheric N. Some plants have developed associations or symbioses with these bacteria, allowing the plant to benefit from the fixed N. The bacteria generally benefit from the relationship because the plant provides an environment in which they can thrive. In some cases the association is a loose one, in which N fixing bacteria simply live around the roots of the plant, feeding off root exudates. Examples of this include the N fixing bacteria Azospirillum and Azotobacter, which are found associated with the root systems of tropical grasses. Sugar cane can be grown using these bacteria as an N source (AbdelMonem et al., 2001, Oliveira et al. 2002).
A closer association is between the aquatic fern Azolla and the nitrogen fixing cyanobacteria Anabaena. Here the two species have evolved together to such an extent that the fern has developed hollow leaves in which the cyanobacteria live. Azolla is an important source of N for many rice farming systems in the tropical and subtropical areas of the world (Ferentinos et al. 2002).
The main group of plants capable of forming symbiotic relationships with N-fixing bacteria, are the legumes. The majority of the Leguminosae are capable of fixing nitrogen by forming symbioses with bacteria of the genus rhizobium. Rhizobia infect the root hairs of legume plants, and root nodules form, which provide an anaerobic environment in which the rhizobium can live. The legume provides the bacteria with sugars as an energy source, the product of photosynthesis, and utilises the N fixed by the rhizobium. As with any mutualistic relationship between species, there are costs and benefits to both species. In particular, maintaining the symbiosis has energy costs for the legume, with the effect that it is more energetically expensive for the legume to fix nitrogen than to utilise soil N. This suggests that the ability to fix N will be more advantageous in situations where N is limiting. Certain trees, notably Alder form associations with the actinomycete Frankia, which are similar to the associations between legumes and Rhizobia.
It is often stated that "When taken up by plants, nitrate and other nutrients are identical in form whether they come from organic matter, soil reserves or applied fertiliser." (e.g. Fertiliser Association of Ireland, 2002).
This does not mean that clover has precisely the same effect on soil as N fertiliser. For instance, legume roots can aerate soil (Patriquin et al., 1995) and improve soil structure (Mytton et al., 1993, Miller and Jastrow, 1996). In practice, there are often major differences between legume based and fertiliser based farming systems. In Britain, legume based farming is likely to involve ley-arable rotation, and the use of animal manure (Philipps and Stopes, 1995), neither of which are necessary in fertiliser based farming. The effects of these factors on soil, plants and the environment are discussed in Section 1.2.8. Also, nitrate is not the only source of nitrogen in soil: ammonium and soluble organic nitrogen (SON) compounds can also be used by plants (Lipson and Nasholm, 2001). A popular dictum in organic farming is "feed the soil not the plants" (e.g. Flowerdew, 1998). Nitrate fertiliser is intended to feed crops in the short term, animal and green manures are intended to improve the soil, producing long term benefits to the farm ecosystem as a whole. The use of N fertiliser does not rule out the addition of organic matter to the soil, in the form of green manures or crop residues (e.g. Yadav et al., 2000), and it is possible that organic matter inputs could be increased in this way. However, in recent decades there has been a tendency for N fertiliser to replace other sources of N in agriculture in the developed world. (Uhlin, 1998).SON is difficult to characterise, but some of it may be amino acids, produced by the breakdown of plant protein. Concentrations of SON in Scottish soils are highest in the presence of plant roots, and removal of vegetation causes this SON to be mineralised into nitrate (Chapman et al. 2001), which strongly suggests that much of this SON is derived from plant material. Plants may be able to access SON after it has been converted to nitrate or ammonium by microbes, and also in some cases by direct uptake. The ability to utilise SON has been observed in plants found in natural ecosystems all over the world (Lipson and Nasholm, 2001), but it has been observed in crop plants as well. Owen and Jones (2001) fed wheat plants with three amino acids, and observed that the wheat plants were only able to utilise 6% of the amino acids, the remainder was absorbed by microbes. Whalen et al. (1999) used 15N labelled dead earthworms to investigate uptake of organic N by perennial ryegrass (Lolium perenne), and found that the organic N pool was depleted significantly more rapidly in the presence of perennial ryegrass. Jones and Darrah (1993) found that maize plants were capable of using amino acids as an N source, and Matsumoto et al. (2000) found evidence that carrot (Daucus carota) and chingensai (Brassica campestris L.) were capable of utilising soluble organic N compounds from the soil.
The behaviour and role of organic nitrogen compounds in the nitrogen cycle is poorly understood. In conventional farming, N is mostly supplied in the form of nitrate or ammonium compounds (or as urea), and the intention is to feed N to the crop plants directly. In organic and low input agriculture, much of the nitrogen is supplied as organic N compounds, in the form of manure, compost, crop residues etc. Organic farming aims to maintain the soil organic matter content so that the soil can supply crops with the N (and other nutrients) that they need. SON may be an important part of this process, and may also be involved in the transfer of nitrogen from legumes to non-legumes.
Organic agriculture is dependent on a cycle of nitrogen that flows through legumes, non-legumes, animals, microbes, air and soil. It is based on a "systematic connexion or co-ordination of parts in one whole." (Scofield, 1986). For this reason, an understanding of the processes of the nitrogen cycle within the farm is extremely important
For the reasons explained in section 1.2.1, comparing the use of legumes and fertiliser N as N sources is complicated because legumes may affect soil in other ways besides simply adding N, and this N may be in different forms to the N applied as fertiliser. Another complication is that the use of legumes in agriculture may require or encourage radically different methods of farming from those associated with fertiliser use.
A wide variety of methods can be adopted to utilise the N fixed by legumes. Legumes in agriculture perform two main functions:
1) Providing protein for human or animal consumption
and/or
2) Providing fixed nitrogen for the benefit of other crops.
When a grain legume such as soybean, is grown and harvested at full seed ripeness, the majority of the fixed N is present in the harvested crop, and soil N may even be depleted by the crop
(Toomsan et al., 1995, Haynes et al., 1993). This is not the case with all grain legumes, faba beans (Vicia faba) have been observed to fix 3 times as much N as is removed in harvested grain (Rochester et al., 1998). In these cases, the primary purpose of growing a grain legume is to provide protein (obtained as far as possible from biological N fixation) for human or animal consumption, but residues of the crop may provide fixed N for the successive crop. If the grain legume is fed to animals (or humans) and the animal (or human) faeces are returned to the soil, then much of the fixed N in the grain becomes available to other crops. Grain legumes can be grown in alternate rows with a non-N-fixing species (intercropping), or sown in a mixture with a non-N fixing grain (mixed cropping). This often has the effect of reducing the availability of soil N to the legume, and thus forcing it to fix a higher proportion of its N (see section 1.6). In addition, the non-fixing species may obtain extra N by N transfer (see section 1.2.3). Intercropping and mixed cropping may result in increased yields of one or both species, or a higher land equivalent ratio. The land equivalent ratio (LER) is the area of monoculture of two species required to produce yields obtained in the mixed crop (Helenius and Jokinen, 1994, Fujita, et al., 1992).Legumes can be grown as green manure crops, and ploughed into the soil in order to improve its nitrogen status prior to sowing of a non-fixing crop (Schmidt et al. 1999). Forage legumes such as clover can be undersown with a taller crop. This provides similar benefits to mixed cropping and intercropping, and can also benefit subsequent crops, smother weeds and provide a habitat for the benevolent, predatory carabid beetles (Armstrong and McKinlay, 1997, Brandsaeter et al., 1998). Perennial legumes, especially leguminous trees, can be cut for composting or mulching (Sanginga et al. 1995), although this is seldom practised in the UK. Cereals can be drilled into an established clover sward (Schmidt et al. 2001, White and Scott, 1991).
In a ley-arable crop rotation, grass and a forage legume such as clover are grown for several years, grazed by animals and/or cut for silage or hay, before being ploughed up so that arable crops can be sown. The last arable crop to be grown before the ley phase can often be undersown with clover. The grass in the ley phase obtains additional N from the presence of the clover; the grazing animals obtain a high protein diet. Manure from the grazing animals transfers fixed N to grass and can be spread on arable crops, which also benefit from the accumulated fixed N that is released when the ley is ploughed (Philipps et al., 1996; Høgh-Jensen, 1996). Ley-arable crop rotations were the main type of agriculture in Europe before the invention of the Haber-Bosch process, for example the Norfolk four course rotation (Lampkin, 1990). This thesis focuses on white clover in ley-arable rotations.
Atmospheric nitrogen is transferred from legumes to non-legumes either by the so-called "below ground pathway", through root exudation and decay of plant tissue (including foliage), or the "above ground pathway", through dung and urine produced by grazing animals. The below-ground pathway is poorly understood. Nitrogen may be released from the legumes by the senescence and decay of leaves, roots etc. Transport of nutrients between plants via mycorrhizal networks has been suggested (Haystead et al., 1988), although the general view is that transfer is a complicated pathway involving a number of organic compounds. Root herbivory by larvae of the Sitona weevil, and damage to clover root systems may enhance nitrogen transfer (Murray and Hatch, 1996, Hatch and Murray, 1994). Defoliation can accelerate root nodule turnover, and this may be one route by which nitrogen is transferred (Ryle et al. 1995).
Dubach and Russelle (1994) have shown that when roots and nodules of alfalfa (Medicago sativa) and Birdsfoot trefoil (Lotus corniculatus L.) are shed, they still contain significant quantities of nitrogen (most other plants efficiently remobilise nutrients from senescing roots and leaves). Estimates of rates of nodule turnover are currently unavailable.In the post war period in the developed world, nitrogen fertiliser has largely replaced nitrogen fixing plants as sources of N in agriculture (Lanyon, 1995). The main exception to this is the soybean, which is widely grown for animal feed, without the use of N fertiliser. Cereals are mostly grown with N fertiliser, high quality pasture is largely maintained by N fertiliser and even some grain legumes are grown using N fertiliser (Redden and Herridge, 1999). The transition to N fertiliser has had numerous environmental and economic effects:
Nitrogen fertiliser has enabled different regions to specialise in crop or animal production (Granstedt, 1991). The nitrogen exported from arable farms, as animal feed no longer has to be returned as manure. In some cases, this has created high concentrations of manure on livestock farms, which must be disposed of (Lanyon, 1995; Granstedt, 1991).
Organic agriculture has been defined a number of different ways. The UN Food and Agriculture Organisation (FAO) define organic agriculture as follows:
"Organic agriculture is a holistic production management system which promotes and enhances agro-ecosystem health, including biodiversity, biological cycles, and soil biological activity. It emphasises the use of management practices in preference to the use of off-farm inputs, taking into account that regional conditions require locally adapted systems. This is accomplished by using, where possible, agronomic, biological, and mechanical methods, as opposed to using synthetic materials, to fulfil any specific function within the system." (FAO/WHO Codex Alimentarius Commission, 1999). Organic agriculture does not permit the use of synthetic N fertiliser, instead relying on biological N fixation as its principal source of N. Organic agriculture also prohibits the use of synthetic pesticides and growth regulators. In Britain, most organic farms consist of ley-arable rotations (Philipps and Stopes, 1995). Biodynamic agriculture is a form of organic agriculture based on the ideas of Rudolph Steiner. Biodynamic agriculture requires adherence to a stricter set of guidelines than ordinary organic farming, and additional practices such as the use of special plant based preparations.
Energy efficiency can be defined simply as the amount of energy required to produce a unit of calorific energy in food. However, for practical reasons it is important to consider whether the energy inputs come from a renewable or non-renewable resource. Sunlight, although variable is not going to run out, and use of human labour creates jobs and brings other benefits, even though it may be financially expensive. The definition of energy efficiency should therefore take into account the amount of non-renewable energy required to produce a unit of food or other produce (Jones, 2002). For the purposes of this study, energy efficiency will be defined as the amount of non-renewable energy required per unit of produce.
In the past, the energy used in agriculture derived mostly from human or animal muscle power, which in turn was derived from the calorific energy in the human or animal food (Pimentel, 1979). All of this energy ultimately derived from solar energy, converted into calorific energy by photosynthesis. Modern agriculture in the developed world often replaces human and animal labour with machines such as tractors, which obtain their energy from fossil fuels (although in future, biofuels, such as biodiesel could become more important). Tilling one hectare of soil by human labour requires 400 man-hours and 8.4 ´ 108 joules ha-1 of energy. A pair of oxen reduces the man-hours to 65, but increase the total energy input to 1.2 ´ 109 joules ha-1 because of the energy that must be provided to the animals as fodder. A 50 hp tractor reduces the man-hours to 4 but increases the total energy input to 2.3 ´ 109 Joules ha-1, because of the energy that must be supplied in fuel and the manufacture of the machinery (Pimentel, 1979).
Manufacture of fertiliser consumes fossil fuels. Natural gas is used as the source of hydrogen, and fossil fuels provide much of the energy required for the process. In 1975, corn production in the USA required, on average, 128 kg of N fertiliser per hectare, the manufacture and transport of which consumed 7.9 ´ 109 joules ha-1. Total energy inputs per ha (labour, machinery, fuel, N, phosphorus (P), potassium (K), lime, seeds, irrigation, pesticides, transport, electricity and processing), consumed a total of 2.7 ´ 1010 J ha-1, of which labour comprised 2.3 ´ 107 J ha-1. The energy efficiency measured as energy output in produce/energy input was 2.93. In 1945, the equivalent figure was 3.7, primarily because increases in energy input have not been matched by increases in productivity (Pimentel, 1979). Fertiliser use accounts for approximately 1/3 of the energy use in US agriculture (Pimentel, 1983). Wittwer (1978) commented that "The current and projected natural gas dependency of chemically fixed nitrogen fertiliser remains as one of the most flagrant violations of good economics, use of a non-renewable resource. It is inconceivable for us to continue to go this route". However, production of ammonia based fertilisers has become more energy efficient over recent years, owing to improvements in the catalysts used. In 1967 4.94 ´ 107 J were required to produce a kg of N fertiliser in the form of calcium ammonium nitrate. In 1998 the equivalent figure was 3.53 ´ 107 J per kg of N fertiliser (Hulsbergen et al. 2002).
These high inputs of fossil fuel derived energy can potentially be reduced by the use of biologically fixed N. Nitrogen fixing plants obtain all of the energy required for N fixation from sunlight. The process does not consume any non-renewable energy sources (Ryle et al., 1979). Laboratory studies have shown that the nitrogenase enzyme (used by rhizobium to fix nitrogen) requires only 1.2 ´ 107 J per kg of N fixed, about 35% of the energy required by the Haber-Bosch process (Postgate, 1979). A survey by Pimentel et al. (1983) estimated energy inputs and outputs (calorific energy in food) in conventional and organically grown corn, spring wheat, potatoes and apples, receiving equal amounts of N as fertilisers, FYM, sewage sludge or green manure. The energy inputs in the form of machinery, fuel, electricity, fertilisers, seeds, pesticides etc. for corn were 2.6 ´ 1010 J ha-1 in the conventional system. For conventional wheat inputs were 1.1 ´ 1010 J ha-1, for conventional potatoes 6.6 ´ 1010 J ha-1 and for conventional apples 1.1 ´ 1011 J ha-1. The inputs for maize in the organic systems ranged from 1.5 ´ 1010 J ha-1 to 1.8 ´ 1010 J ha-1; organic wheat 7.2 - 7.8 ´ 109 J ha-1; organic potatoes 3.5 - 3.9 ´ 1010 J ha-1 and for organic apples 8.4 - 8.8 ´ 1010 J ha-1. The organic systems therefore used less fossil fuel energy than the conventional systems. The ratio of energy output to energy input in the conventional corn system was 4.47; in the conventional wheat 2.38; conventional potatoes 1.28 and conventional apples 0.89. In the organic systems the equivalent figures were 5.75-7.6, for organic corn, 3.22-3.49 for organic wheat, 1.07-1.2 for organic potatoes and 0.06 for organic apples. The energy efficiency of organic cereal production was therefore greater than that of conventional cereals. The reduced energy efficiency of organic potato production was due to the need for mechanical weed control and the reduced yields. The reasons for the extremely low energy efficiency in organic apple production will be discussed later. The organic systems required between 4 and 64% more human labour. Yields of conventional corn were estimated at 8 t ha-1, conventional wheat, 1.9 t ha-1, conventional potatoes 33 t ha-1 and conventional apples 41.5 t ha-1. Yields of organic corn were estimated at 7.9 t ha-1, organic wheat, 1.8 t ha-1, organic potatoes 16.5 t ha-1 and organic apples 2.1 t ha-1. The exceptionally low estimate for organic apple yield is based on the assumption that without pesticides 95% of the apples would be unsuitable for sale. These high losses due to cosmetic damage are also responsible for the low energy efficiency of the organic system (Pimentel, 1983). In contrast Reganold et al. (2001) found that the energy output/input ratio for conventionally grown golden delicious apples was 1.11 and in an organic system 1.18. This latter study ignored the effect of russeting (roughening of the skin) on marketability of apples, on the grounds that, while organically grown apples had a higher level of russeting, this did not make them unacceptable to consumers. In some varieties of apples and pears, russetting is a desired trait (Reganold et al. 2001).
The studies described above only considered the yields of individual crops. In an organic system, arable crops will usually be grown as a rotation, and the efficiency of the rotation as a whole must be considered. Mader et al. (2002) found in 21-year trial of conventional organic and biodynamic ley-arable rotations in Switzerland, that yields were reduced by 20% in the organic systems. However, total energy inputs were 2.1-2.4 ´ 1010 J ha-1 year-1 in the conventional systems but only 1.3 ´ 1010 J ha1 year-1 in the organic systems. It is important to note that this was a comparison between organic and conventional ley arable rotations, not between fertiliser and ley arable systems. In all systems there was a need for energy to be expended ploughing leys. Improvements in soil structure caused by clover may reduce this energy requirement (Mytton et al. 1993).
In the above cases, the reason for the generally higher energy efficiency in organic systems was the energy used in the manufacture and distribution of fossil fuels, but the organic systems required additional energy to be used in mechanical weed control etc. This, combined with the lower yields reduced the energy efficiency of the organic systems. It is reasonable to examine the potential for making conventional agriculture more energy efficient, without adopting all of the rules of organic farming. Hulsbergen et al. (2002) examined the energy efficiency of various rates of N fertiliser on a rotation of 5 arable crops: potatoes, winter wheat, winter barley, sugar beet and spring barley in central Germany between 1968 and 2000. The study considered inputs of fossil fuel energy, but ignored human labour. In order to achieve the optimum energy output/input ratio for potatoes, the optimum N rate was 54-101 kg ha-1, which achieved an output/input ratio of 4.7-5.2. For winter wheat, the optimum N rate was 46-54 kg N ha-1 producing an output/input ratio of 10.5-13. For winter barley, the optimum N rate was 85-102 kg N ha-1, producing an output/input ratio of 8.7-10.5. For sugar beet, the maximum energy output/input ratio was achieved with 0 kg N ha-1, producing an output/input ratio of 15-15.2. For spring barley the optimum N rate was 60 kg N ha-1, producing an output/input ratio of 7-7.2 kg N ha-1. When by-products such as straw are included, the output/input ratios are even higher (Hulsbergen et al. 2002). The energy efficiency values of cereals and potatoes obtained in the aforementioned study are noticeably higher than the values obtained by Pimentel et al. (1983) for both organic and conventional systems. This may be partly due to the increased efficiency of N fertiliser production (estimated at 50 MJ kg-1 by Pimentel et al. 1983, and 35.3 MJ kg-1 in the quoted data from Hulsbergen et al. 2002). The lower energy efficiency in the study by Pimentel et al. (1983) may also mean that maximum energy efficiency may not be the same thing as maximum profitability (see section 1.2.7). Currently the average rates of N application to the principal arable crops in the UK are 189 kg N ha-1 for winter wheat, 114 kg N ha-1 for spring barley, 149 kg N ha-1 for winter barley, 175 kg N ha-1 for maincrop potatoes, 196 kg N ha-1 for oil seed rape and 106 kg N ha-1 for sugar beet (Defra, 2001). The yields of these crops per hectare in 2000 were 7.26 tonnes ha-1 for wheat, 5.18 tonnes ha-1 for spring barley, 6.87 tonnes ha-1 for winter barley, 54 tonnes ha-1 for sugar beet and for potatoes 41.11 tonnes ha-1 (DARD, 2001; National Statistics online, 2003). The equations for N fertiliser response calculated by Hulsbergen et al. (2002), underestimated British potato yields by 30.9%, wheat yields by 31%, sugar beet by 2.6% and spring barley by 18%. Winter barley yields were overestimated by 9.6% (equations [1.3.1] – [1.3.5], below). Assuming that energy expenditure on machinery use etc. is the same on British and German farms, the energy output/input ratios for the British arable crops are as follows: potatoes 6.7, winter wheat 7.9, winter barley 8.4, sugar beet 13.2 and spring barley 7. This assumes 20% dry matter in potatoes (Dobozi and Lehoczky, 2002) and 25% dry matter in sugar beet (British Sugar, 2003). These figures suggest that for some arable crops, such as winter wheat, British farmers are currently using higher levels of N fertiliser than the optimum for maximum energy efficiency.
Would the energy efficiency of the arable crops studied by Hulsbergen et al. (2002) be improved if N was supplied by a grass-legume leys rather than fertiliser? To calculate this, it is necessary to know how much N is supplied to subsequent crops by a ploughed ley, and the amount of energy expended in ploughing the ley. Nevens and Reheul (2002) found that a ploughed 3-year-old ley in North West Europe can add 231 kg N ha-1 to successive crops over 3 years (150, 52 and 29 kg N ha-1 in the first, second and third years respectively after ploughing of the ley). Adams and Jan (1999) observed that a subsequent crop of ryegrass was able to utilise 30-100 kg N ha-1 released by the ploughing of a grass-clover ley. The ley phase of a ley-arable rotation produces, meat, milk, wool etc. as well as adding N to the soil, and the energy inputs and production should be calculated separately from the inputs and outputs of the arable phase. The energy required to plough a ley, and prepare a seed-bed from it, prior to planting of an arable crop can be assumed to be equivalent to the "fertiliser energy input", and will be referred to as "ley energy input". Mouldboard ploughing to a depth of 300 mm requires 27.6 L of diesel ha-1, equivalent to 1.1 ´ 109 J ha-1 (Sijtsma et al. 1998, Hulsbergen, et al. 2002). Ploughing to 200 mm as favoured by some organic farmers requires 20 L of diesel ha-1 or 7.9 ´ 108 J ha-1, because shallow ploughing requires less traction than deep ploughing (Kouwenhoeven et al. 2002, Hulsbergen, et al. 2002). Subsequent seedbed preparation requires between 1.6 ´ 106 J ha-1 and 4.9 ´ 105 J ha-1, depending on the machinery used (Chamen et al. 1998), giving a total of between 1.3 ´ 109 J ha-1 and 2.8 ´ 109 J ha-1 depending on tillage methods. Assuming that ploughed leys provide between 29 and 150 kg N ha-1, we can then use the N input/energy yield curves of Hulsbergen et al. (2002), to calculate a figure for energy output/ energy input. This will give an estimate of the energy efficiency if leys rather than fertiliser had been used. The N input/energy yield curves (averaging data from 1989-93 and 1994-98 rotations) are:
Potatoes:
![]() [.3]
[.3]
Winter wheat:
![]() [.4]
[.4]
Winter barley:
![]() [.5]
[.5]
Sugar beet:
![]() [.6]
[.6]
Spring barley:
![]() [.7]
[.7]
Where E is the energy output in the crop and N is the N input either from fertiliser or from ploughed leys
(Hulsbergen et al., 2002, average of 1989-93 and 1994-98 rotations)
The figures for N input/ley energy input for crops grown after ploughing of a ley are then 4.1-5.9 for potatoes, 9.8-12.9 for winter wheat, 5.7-12.4 for winter barley, 12.9-15.1 for sugar beet and 4.4-7.7 for spring barley, similar to the maximum energy efficiencies calculated by Hulsbergen et al. (2002). This suggests that the production of arable crops in a ley arable rotation could potentially be as energy efficient or in some cases more energy efficient than production from all-arable systems.
Energy efficiency of arable crops in a ley-arable rotation depends partly on the amount of N transferred from the ploughed ley to the subsequent arable crop, and the amount of energy expended in ploughing the ley. Rather than looking at the arable stage of the rotation in isolation, it is important to also consider the energy efficiency of the ley phase of the rotation. Mader et al. (2002) calculated that a three year biodynamic clover ley requires 15.2 ´ 109 J of energy input over its lifetime. An organic three year ley requires 18.4 ´ 109 J, and a conventional ley requires 22 ´ 109 J, or 39 ´ 109 J, if it receives only mineral N (no manure). Most of the additional energy in the conventional leys was due to fertiliser use (FIBL, 2002). In Britain in 2001, the average rate of N application to grasslands was 130 kg ha-1 for grazed land, 165 kg ha-1 yr-1 for silage plots and 85 kg ha-1 yr-1 for hay plots (Defra, 2001). This amounts to 4.5 ´ 109 J ha-1 yr-1, 5.8 ´ 109 J ha-1 yr-1 and 3 ´ 109 J ha-1 yr-1 of fossil fuel derived energy for grazed, silage and hay producing land respectively. Is this input of non-renewable energy justified by the yields?
Attempts have been made to calculate the benefit to grass from clover, in terms of the amount of nitrogen fertiliser required producing the same yield of grass in the absence of clover. This is known as the fertiliser N equivalent. Estimates of fertiliser N equivalent range from 124-275 kg N ha-1 (Royal Society, 1983). Another study found that grass/legume swards are as effective as grass monocultures receiving 150-200 kg N ha-1 year-1, in terms of lamb production, and superior to the monocultures in the post-weaning period. The grass/legume sward could support fewer animals, but the improved performance of the individual animals compensated for this (Frame et al., 1998). The results of these studies strongly suggest that biologically fixed N could replace much of the fertiliser N currently used on grassland in the UK, reducing the fossil fuel energy inputs significantly without affecting production.
Ledgard et al. (1999) compared N inputs and outputs from dairy farmlets receiving 400, 200 and 0 kg N ha-1 in New Zealand over 3 years. N fixation in the 0N farmlet was 99-231 kg N ha-1, whereas in the 200 N farmlet it ranged from 75-149 kg N ha-1, and in the 400 N farmlet N fixation ranged from 15-44 kg N ha-1. It can be seen from this that inputs of biologically fixed N are reduced in proportion to inputs of mineral N. This subject will be covered in more detail in section 1.6. Dairy farms in the Netherlands typically use 331 kg N ha-1 of fertiliser, 181 kg N ha-1 in purchased feed and virtually no biologically fixed N (Ledgard et al., 1999). The ratio N output/N input is 0.14. Dairy farms in New Zealand typically use 40 kg N ha-1 of fertiliser, 4 kg N ha-1 in purchased feed, and 140 kg N ha-1 of biologically fixed N. The ratio of N output/N input is around 0.3. Milk production per hectare in the New Zealand system is about 70% of that in the Dutch system (Ledgard et al., 1999).
The examples given above show that simple comparisons of herbage yield between conventional and organic/legume-based grassland are not sufficient for a comparison of energy efficiency or economic efficiency. Grassland containing clover may offer greater nutritional benefits to grazing animals than a pure grass sward, even if the total herbage yields are lower. Clover also has nutritional benefits for grazing animals, having more protein than grass alone, is quicker to digest (Beever and Thorp, 1996), and remains digestible for longer than L. perenne with increasing maturity (Thomson, 1984). Cows and ewes fed on clover produce more milk, and their milk contains more total protein, more casein and has greater coagulum strength. This makes the milk more suitable for cheesemaking, and is probably the reason for the rapid growth of lambs on clover pastures (Thomson, 1984, Newton et al., 1983).
As we have seen from the examples in section 1.2.6, legumes can be an energy efficient way to add fixed nitrogen to agricultural systems, especially in the case of grasslands. However, in Britain and in much of the developed world, the use of N fertiliser is more popular as a source of N for agriculture. This section will discuss the reasons for this.
Nitrogen fertiliser is a simple and convenient way of adding nitrogen to the soil. Yields obtained with nitrogen fertiliser-based systems are generally larger than those obtained from other systems (Loomis and Connor, 1992). The amount of nitrogen that can be added to soil this way is only limited by the cost of the fertiliser, and by environmental regulations. Potential maximum crop production can be calculated from experiments in which the roots of the plant in question are provided with optimal growing conditions as regards, water, minerals and oxygen, so that yield becomes dependent on incoming light energy. Monocultures of grass treated with nitrogen fertiliser have a theoretical maximum yield of 30 t DM ha-1 year-1 in the UK (Alberda, 1971; Cooper and Breese, 1971). Some fertilised grasslands in Northern Ireland are currently producing 25 t ha-1 (DARD, 2001). In practice, yields are typically in the region of 13-15 t DM ha-1 year-1 in temperate climates (Robson, 1982, Loomis and Connor, 1992, Cooper and Breese, 1971). As long as fertiliser remains relatively cheap, relative to the prices of agricultural products, fuel and human labour, as well as the levels of agricultural subsidies, it makes good economic sense for farmers in developed countries to use it. However, hidden costs are associated with pollution resulting from fertiliser use, which must be paid for by society at large. In the UK in 1996, removing nitrate from drinking water cost an estimated £16m (Pretty et al. 2000). Water companies, such as the German Stadtwerke München, now pay farmers to convert to organic farming to reduce pollution of rivers, although the even higher cost of removing pesticides from drinking water (£120m per annum in the UK, Pretty et al. 2000), probably influences these decisions as well.
The use of grass-white clover pastures in New Zealand has shown their economic viability. The production from these pastures is among the most economical in the world (Frame et al., 1998). Other factors also contribute to the profitability of New Zealand grasslands: New Zealand has a very favourable climate that allows white clover to grow almost all year round, large farms. Production from grassland in New Zealand is dependent upon phosphate fertiliser (MAF, 1974). It has been estimated that a total switch to legume-based systems would benefit UK livestock producers by approximately £300m a year (Doyle and Bevan, 1996). The main reasons why legume-based systems have not been widely adopted are technical, rather than economic (Doyle and Bevan, 1996).
One disadvantage of white clover is that its growth is slow, during the spring, so yield benefits are not apparent until the summer months (Sprent and Mannetje, 1996). However, observations at Haughley Research Station showed that in most years organic pastures began to grow earlier than conventional ones (Balfour, 1976). Poor spring growth is probably due to stolon death over winter, and resulting patchiness of clover distribution (Rhodes and Ortega, 1996), as well as the lower temperatures (see section 1.7.5). Addition of nitrogen fertiliser can inhibit the growth and nitrogen fixation of clover, and cause it to suffer from competition with grass (e.g. Høgh-Jensen and Schjoerring, 1994). Another disadvantage is that clover seeds are small, making them hard to establish. Clover seed must be sown no deeper than 13 mm in a fine seed bed, with soil pH no less than 5.5 (Frame and Newbould, 1986) and they are not persistent. Growth can fluctuate dramatically from year to year, and the nitrogen yield is lower than that obtained with nitrogen fertiliser. The reduced N yield makes clover less attractive from an economic point of view (Sprent and Mannetje, 1996; Stewart and Haycock, 1984). Clover can cause bloat (tympanites) in livestock (D’Mello and MacDonald, 1996), and red clover can also cause fertility problems in livestock (Wong, 1973). Plant breeding programs are currently attempting to rectify these problems (Rhodes and Ortega, 1996). In low input farming a strong economic case can be made for the use of legumes. In organic farming they are essential.
While inputs of nitrogen have increased since the introduction of nitrogen fertiliser, efficiency of nitrogen use has declined, with consequent pollution of groundwater and the atmosphere. In the post war period, annual artificial N use in Sweden increased from <15 kg ha-1 to 79 kg ha-1, but output did not increase accordingly, in 1980, four times as much N was applied to Swedish farms as was recovered in produce (Granstedt, 1991). In the USA, the separation of animal and crop production, and the increase in animal production that occurred as a result of the use of N fertiliser, caused the efficiency of N use to decline. A high percentage of the applied N was lost to groundwater and the atmosphere (Lanyon, 1995). Nitrogen is lost to the atmosphere as NH3 and N2O (a greenhouse gas), and losses of N by this rate can be greater in fertiliser based systems than in legume based systems (Jensen and Hauggaard-Nielsen, 2003). When N is also lost to groundwater, it can cause eutrophication of natural habitats (Howarth, 1998) and pollution of drinking water. The high concentration of dung and urine N on animal production farms increases N losses (Granstedt, 1991).
Organic farming is widely believed to cause less N pollution of groundwater than conventional agriculture, but this belief is not always supported by the evidence. A number of studies have shown that legume-based pastures lose less nitrogen to groundwater than nitrogen fertilised pastures producing similar yields (Tyson et al. 1996; Drinkwater et al., 1998; Ruz-Jerez et al., 1995). Goulding (2000) reviewed data on leaching from conventional and organic leys and found that conventional grass-only first year leys leached around 1.5 times as much N as first year organic grass-clover leys, and second year conventional leys leached around 3 times as much N as second year organic leys. However, leaching from first year organic arable crops was around twice as high as leaching from first year conventional crops, owing to the greater N losses resulting from ploughing of grass-legume leys. N losses from second year conventional arable crops were around 1.5 times higher than N losses from second year organic arable crops. In the third year of arable crops leaching losses were around 1.5 times higher in the conventional system. Over the whole rotation, leaching losses of N were only slightly lower from the organic system.
Mixed farming systems also use nitrogen more efficiently than solely arable farms or pastoral farms (Granstedt, 1991). There is however, a risk of nitrate leaching, following the ploughing of pastures (Scholefield and Smith, 1996). Philipps and Stopes (1995) note that while leaching from this stage of the rotation is high, the lower losses during the rest of a typical organic rotation compensate for this, so the average nitrogen loss is 10.3-20.8 kg N ha yr-1. Kristensen et al. (1995b) recommend that following the ploughing of a grass clover ley, the best crops to grow are those with a high N demand and/or a long growing season, such as sugar beet, brassicas or cereals. Leaching losses can be minimised if ploughing is done in spring, or late in autumn. An alternative is to plough early in the autumn and plant a catch crop (Kristensen et al., 1995b). Eltun (1995) compared N leaching and surface run off from experimental organic and conventional ley-arable rotations in Norway. Conventional arable fields lost 41.32 kg N ha-1 as opposed to 13.6 kg N ha-1 from organic arable land. Leaching and run off of N from conventional leys totalled 22.82 kg N ha-1 and from organic leys 14.78 kg N ha-1. The fact that in this comparison, organic leys were ploughed in spring and conventional leys in autumn may account for some of the difference observed (Eltun, 1995). A study by Stopes et al. (2002) compared organic farms to conventional farms with similar crop rotations and climate, and found that N leaching was similar or slightly lower from the organic farms when crop yields and inputs were taken into account. Kristensen et al. (1995b) observed no significant differences in the levels of leachable N in soil from conventional and organic farms, but noted that other factors such as crop type; soil type and precipitation made the comparison difficult. A review by Kirchmann and Bergstrom (2001) found that organic farms had lower nitrogen leaching losses, but they also had lower N inputs than conventional farms, so they were not any more efficient in their use of N inputs than conventional farms. The review was unable to compare crop yields and N leaching on organic and conventional farms, which is unfortunate, because crop yields are more relevant to this discussion than N inputs. Webster et al. (2003) showed that under a low input integrated conventional system, N leaching losses were 67 kg N ha-1 from beans and peas, 46 kg N ha-1 under winter wheat and barley, and 24 kg N ha-1 from grass clover leys. Applied N fertiliser rates were 0 kg N ha-1 for beans and peas, 141 kg N ha-1 for winter wheat, 137 kg N ha-1 for winter barley and 68 kg N ha-1 for grass clover (Webster et al. 2003). This makes the efficiency of fertiliser use in the integrated system for winter wheat (N output/N fertiliser input) 0.49 and for barley 0.62.
Milk yields from grass-clover pasture with no applied N in New Zealand were found to be 83% of those from pastures receiving 400 kg N ha-1, but the efficiency of N use was much greater when no N fertiliser was applied. At the high rate of N application, only 26% of the fertiliser N and biologically fixed N was recovered in milk and other produce. When the pasture was reliant on biologically fixed N, 52% of the N input was recovered in the farm produce. Applying 400 kg N ha-1 as fertiliser increased losses of N to the atmosphere and to groundwater by a factor of approximately 3.7-3.9 (Ledgard et al. 1999).
It is difficult to make comparisons between conventional and organic farms because so many conventional farms are not self-contained or self-sufficient units, as organic farms ideally should be, but rather they are often part of a broader national and international food chain. In particular, the separation of arable production and animal production that has resulted from the use of N fertiliser has also affected the efficiency of N use.
Granstedt (1995) examined the N use efficiency of individual conventional and organic farms in Sweden, and the communities that depended on them. Over the whole of Sweden, 86 kg N ha-1 was lost from farms to denitrification and leaching. In addition, 19 kg N ha-1 was lost in the wider community as food waste, slaughterhouse waste, domestic waste, and sewage (Granstedt, 1995).
In one area that specialised in intensive animal production, with an average of 0.8 livestock units (LU) per hectare (1 LU is equivalent to a large dairy cow), 101 kg N ha-1 were imported as fertiliser, 34 kg N ha-1 were imported as animal feed and 9 kg N ha-1 were fixed biologically. From this system, 26 kg N ha-1 were exported as animal products and 30 kg N ha-1 as crops. Total N losses were 101 kg N ha-1 from farms and 14 kg N ha-1 from the wider community. Total N output/N input (N in produce/N inputs) was 0.34 (Granstedt, 1995).
An area with a moderate intensity of animal production (0.5 LU ha-1) imported 82 kg N ha-1 as fertiliser, 3 kg N ha-1 as animal feed and 13 kg N ha-1 was fixed biologically. From this system, 15 kg N ha-1 were exported as animal products
and 6 kg N ha-1 as crops. Losses from this system were 97 kg N ha from farms and 96 kg N ha from the community. Total N output/N input was 0.178 (Granstedt, 1995).An area with a lower intensity of animal production (0.2 LU ha
-1) imported 76 kg N ha-1 as fertiliser, 6 kg N ha-1 as animal feed and 5 kg N ha-1 were fixed biologically. From this system, 6 kg N ha-1 were exported as animal products and 39 kg N ha-1 as crops. Losses from this system were 42 kg N ha-1 from agriculture and 17 kg N ha-1 from the community. N output/N input was 0.44. In general, N losses were greatest in areas with intensive animal production. (Granstedt, 1995).In the same study, a mostly arable farm, a dairy farm and a pig production unit were compared to a biodynamic farm at the farm level. The arable system imported 97 kg N ha-1 as fertiliser, 2 kg N ha-1 as feed, 7 kg N ha-1 were fixed biologically, 102 kg N ha-1 was sold as produce and 19 kg N ha-1 was lost to leaching and denitrification. The dairy farm imported 91 kg N ha-1 as fertiliser, 15 kg N ha-1 as feed, 17 kg N ha-1 were fixed biologically and 87 kg N ha-1 was lost to atmosphere and groundwater. The pig unit imported 103 kg N ha-1 as fertiliser, 135 kg N ha-1 as feed, 4 kg N ha-1 were fixed biologically, 93 kg N ha-1 were sold as animal products and 166 kg N ha-1 was lost to the atmosphere and groundwater. The biodynamic mixed farm imported no fertiliser, 3 kg N ha-1 as feed and 50 kg N ha-1 were fixed biologically, 7 kg N ha-1 were exported as crops and 14 kg N ha-1 as animal products. Total losses of N to leaching and denitrification were 45 kg N ha-1. The biodynamic farm had a yield similar to the Swedish average. Losses of N to leaching were about 25% of the national average, and gaseous losses were 50% of the national average. The biodynamic farm was self-sufficient in nutrients, the other farms were connected in a larger system, and this system had considerable losses of N, because of an imbalance between crop and animal production at both national and local levels. (Granstedt, 1995).
Where cereals are grown for many successive years without a break in the rotation, there is a risk that soil organic matter will be lost (Uhlen, 1991), releasing carbon dioxide into the atmosphere (Pulleman et al. 2000). Soil organic matter supplies essential nutrients and protects them from leaching (Davies, 2000), retains water, raises soil temperature and absorbs cations. It is also needed to maintain soil structure and aeration. Low soil organic matter also makes soils vulnerable to wind erosion. Approximately 2.2 million tonnes of topsoil are lost every year in England and Wales to erosion (Harrod and Fraser, 1999). In the USA, on average 16 tonnes of soil are lost per hectare per year (USDA, 1991). At most 1 tonne of new soil is formed per hectare per year (Pimentel, 1993), which means that losses of N in soil erosion (an estimated 64 kg N ha-1 year-1 in the USA) exceed fertiliser inputs (average 48 kg N ha-1 year-1, Pimentel, 1996). Forage legumes are particularly good for improving soil structure, because they have a high rate of root turnover, providing substrates for bacteria that produce polysaccharides, important structural components of soil. Decomposing legume roots and mycorrhizal hyphae bind soil aggregates together, increasing their stability (Miller and Jastrow, 1996). A study by Drinkwater et al. (1998), found that legume-based cropping systems retain more soil carbon than those based on nitrate fertiliser. This was attributed to the low C:N ratio of the legume residues, and the greater temporal diversity of the cropping sequences in the organic rotations examined (Drinkwater et al., 1998). There is some disagreement about the effect of mineral N fertiliser on soil organic matter. Thomsen et al. (2003) examined the long term (20-28 years) effects of different N fertiliser applications on spring and autumn sown cereals grown on sandy loam, and concluded that soil organic matter was not increased by high rates of N application. In a 30-year experiment comparing the effects of leys, manure, crop residues and mineral fertilisers, soil carbon content in ley- arable rotations with or without manure remained constant or showed small increases. All cereal rotations with no return of straw showed a 5% decline in soil carbon content (Uhlen, 1991). Kapkiyai et al. (1999) compared different farm management techniques in East African smallholdings. Losses of soil organic matter were greater from N fertiliser-only treatments, than from manure treatments. In contrast, Kidanu et al. (2000) observed that soil organic matter in the Ethiopian highlands was increased by applications of N fertiliser.
Organic farming and other legume-based farming systems are widely believed to pose less of a risk of nitrate leaching than farming systems based on fertiliser. However, the environmental benefits in this regard are less than is commonly imagined. In addition, it should be stressed that the terms "organic" and "conventional" both cover a wide variety of farming systems, so it is difficult to make general statements. The ratio of pasture to arable land, stocking density, and the use of autumn sown crops have considerable impacts on the amount of N leached, and these factors may be more significant than whether legumes, manure or N fertiliser are used. There is some evidence that in the long term, organic farming systems may benefit soil and prevent erosion. Replacing N fertiliser with biologically fixed N could improve the energy efficiency of agriculture, especially in grassland, although organic farming systems have additional energy costs of their own, resulting from mechanical weed control etc., as well as generally lower yields, which can reduce their energy efficiency.
White clover (Trifolium repens L.) is a leguminous plant native to the UK. It is a creeping perennial, although its growth and morphology are highly variable:
Seedlings consist of a taproot and stem, from which leaves, petioles, flowers and stolons grow. The stolons branch and form adventitious roots, as well as more leaves, petioles and flowers. After 2 or 3 years of growth, the central taproot senesces, and eventually the stolons become independent plants. Stolons may also become detached from the parent plant by treading or grazing (Caradus, 1990, Frame and Newbould, 1986). Consequently, white clover can spread rapidly, and completely change its pattern of distribution in the space of two years (Thorhallsdottir 1990a,b). White clover produces a variable proportion of "hard" seeds that are slow to germinate, and can remain viable for several years in the soil (Frame and Newbould, 1986). There is some evidence that the degree of hardness in clover seed may be related to the availability of water at the time of seed formation, so it is possible that hard seeds are an adaptation to drought conditions (Thomas, 1987). Clover seeds can pass through the guts of ruminants and subsequently germinate. This may be a significant factor in their dispersal (Young, 2002). White clover morphology varies considerably between varieties. Small leafed varieties, such as Kent wild white, are low growing, have long, branching stolons and are well suited to grazing. Large leafed varieties, such as the Ladino varieties are taller growing, have shorter, less branched stolons and are well suited to cutting for silage (Frame and Newbould, 1986).
The Romans were aware of the beneficial effects of legumes on soil fertility, and white clover seems to have been held in high regard for many centuries. Homer describes it as "The lotus" and mentions it as fine fodder for horses. In Welsh mythology it is said to have sprung up wherever the Princess Olwen trod, and is celebrated "with praise out of all proportion to its beauty" (Graves, 1948). The deliberate cultivation of white clover probably began in the 13th century in Moorish Andalucia. Over subsequent centuries it spread first into Lombardy and the Netherlands (both areas under Spanish control), and in 1620, clover seed was exported from the Netherlands to England (for this reason, white clover is sometimes known as Dutch clover). The introduction of white and red clover had wide-ranging consequences: forage and grain production increased dramatically. The potato ceased to be a luxury vegetable and became part of the European staple diet as a result of the improved soil fertility. In Denmark, cattle numbers increased by 1/3 and grain production doubled within 35 years when clover was introduced. The cultivation of clover was a major driving force for the agricultural revolution, and made productive low input agriculture possible (Kjaergaard, 1995).
Today white clover is the most widely grown forage legume in temperate zones, and is especially popular in Europe and in New Zealand. It is also used as a green manure and for undersowing. White clover is an extremely valuable, renewable source of N and animal feed for agriculture. However, during the twentieth century, there has been a tendency for synthetic N fertiliser to replace white clover and other legumes as the source of N for grasslands and agriculture as a whole. The reasons for this have been discussed in section 1.2.7.
The simplest method of estimating nitrogen fixation is to compare yields from legume-based systems receiving no N and legume-free systems receiving varying amounts of fertiliser e.g. organic grass-clover leys and conventional grass-only leys. The fertiliser N equivalent is the amount of N required by the legume free system to match the yield of the legume-based system. One possible source of error in this method is that soil might be enriched under legumes simply because legumes might be less efficient at depleting soil N than non-legumes. This is known as the N sparing effect (Senaratne and Hardarson, 1988). There is also the possibility that N released from the roots of the legume could stimulate the release of additional N from the soil, e.g. by stimulating the mineralisation of soil organic N (Laidlaw et al., 1996).
A more sophisticated version of this approach is to compare the total N content of legume and non-legume biomass:
A non-legume monoculture compared to a mixture of the same non-legume and a legume, grown on the same soil, under identical conditions. Total yield of nitrogen is measured for both plots and species. Assuming that both mixture and monoculture are equally capable of taking up nitrogen from the soil, any additional nitrogen accumulated by the legume mixture will be due to nitrogen fixation (Elgersma et al. 2000).
The legume and non-legume monoculture may not have identical patterns of nitrogen uptake throughout the growing season, and they may not have access to the same sources of soil nitrogen (Bremer et al., 1993). Turkington and Burdon (1983) observed that T. repens and L. perenne (the usual reference crop in 15N dilution studies of T. repens) have asynchronous growth cycles, and that this explains their ability to cohabit. Grasses such as L. perenne are also known to have a much more extensive and finely branched root system than clover, and longer root hairs. Only 68 % of white clover roots have root hairs, as opposed to 95% in L. perenne. The root hair cylinder (the volume of soil contained within the span of the root hairs around the main root) of L. perenne is far larger than the root cylinder of T. repens. The root hair cylinder of L. perenne has a volume of 411 mm3 per mg of root dry matter. The equivalent figure for T.repens is 68 mm3 mg-1 (Evans, 1977), and this may allow L. Perenne to exploit soil N more effectively than white clover. Non-nodulating mutant varieties of some legumes exist, and these are the ideal reference plants. Unfortunately non-nodulating white clover varieties are not currently available (Warembourg, 1993). In laboratory studies, legumes can be grown as reference plants in sterile conditions, in the absence of rhizobium. The non-legume monocultures used for estimates of nitrogen transfer should ideally be as close to the mixed plots as possible, because of soil spatial variation (Reichardt et al., 1987).
An even simpler method of estimating N fixation is to assume that nitrogen fixation is proportional to dry matter yield, and simply weigh the legume plants. This method is useful for quick comparisons of large numbers of varieties of a single species of legume, or of rhizobium strains (Hardarson and Danso, 1993). The method assumes that there are no significant variations in pNdfa or N concentration in plant tissues. Correlations have also been found between nodule weight and number, and nitrogen fixation calculated by other methods (Hardarson and Danso, 1993). Carranca et al. (1999) observed significant differences between estimates of N fixation by subterranean clover using this method, and estimates obtained by isotope dilution and natural abundance.
Nitrogenase, the enzyme which rhizobium use to convert N2 into NH3, also converts acetylene into ethylene (Warembourg, 1993). By incubating legume root nodules or whole root systems in an atmosphere containing acetylene, and measuring the rate at which the acetylene is converted into ethylene, nitrogenase activity can be measured, although the ratio of nitrogenase activity to ethylene production varies between species (Warembourg, 1993). If the total number and mass of root nodules can be measured or estimated, then this method can be used to quantify the total amount of N2 fixed. Errors may occur because microorganisms other than rhizobium may produce ethylene and the treatment may inhibit the rhizobium activity (Wood, 1995). The technique can only provide short-term measurements of nitrogen fixation. Nitrogen fixation may vary throughout the course of a day. Attempts to extrapolate results obtained this way over the course of a growing season are likely to be inaccurate (Caldwell and Virginia, 1989). The acetylene reduction assay is known to produce significantly different estimates of nitrogen fixation from other methods (Martensson and Ljunggren, 1984).
Soil N and atmospheric N may naturally contain slightly different concentrations of the isotope 15N, and this can be utilised to estimate the uptakes of soil and atmospheric N by a legume (a natural abundance technique). The 15N abundance of available soil N is hard to determine, so usually a comparison with a non-fixing reference plant is made. The 15N abundance of fixed N is sometimes slightly different from the natural abundance of atmospheric N, because nitrogen fixation preferentially uses certain isotopes. This difference (known as an isotopic fraction effect) can be taken into account when estimates of nitrogen fixation are made (Caldwell and Virginia, 1989). Natural abundance techniques require extremely accurate measurements to be made, because the differences in natural abundance are extremely small, relative to background variation (Handley, 1996). Typically the d 15N values of clover may vary by less than 1% (e.g. Hanson et al., 2002). Care must be taken to avoid contamination of the samples. Often it is necessary for a mass spectrometer to be used solely for natural abundance, if accurate results are to be obtained (Shearer and Kohl, 1993). Estimates of N-fixation by this method have been known to differ from those obtained by 15N dilution (Høgh-Jensen and Schjoerring, 1994), although Carranca et al. (1999) observed no significant differences between 15N dilution and natural abundance methods. This method suffers from the same problems with reference plants as the N difference method.
Natural differences between soil and atmospheric nitrogen 15N abundance values are often small, variable or non existent, so the soil nitrogen pool is often artificially labelled with 15N. 15N-labelled fertiliser is added to the soil in which nitrogen fixing and non-fixing reference plants are growing. The uptake of the label is compared in nitrogen fixing and non-fixing species (e.g. Laidlaw et al. 1996). The percentage of N derived from the atmosphere (pNdfa) is calculated from:
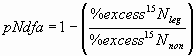 [.1]
[.1]
(Wood, 1996), where %excess15Nleg is the percentage enrichment of 15N in the legume as compared to the background (unlabelled) 15N level in the legume, and %excess15Nnon is the enrichment of the non-legume.
Nitrogen fixation is calculated as:
![]() [.2]
[.2]
Where %Nleg is the percentage N concentration of legume dry matter and DM is the dry matter yield of the legume. A comparison of label uptake in the non-fixing species, grown alone and in mixture with legumes is used to provide estimates of nitrogen transferred from the legume to the grass (McNeill and Wood, 1990). Total fixation is often assumed to be equal to the amount of atmospheric derived nitrogen in the legume, plus atmospheric nitrogen (N transfer) in the non-legume (Goodman, 1988). Fixed N transferred from the legume to the soil is not measured in this or any other method described here.
This method suffers from much the same problems as the total nitrogen difference and natural abundance methods, with respect to reference plants.
It assumes that both the legume and the non legume must take up soil N in proportion to the amount of soil N available, and that the ratio of fertiliser N:soil mineral N available to both species must be the same. The soil enrichment is likely to decline over time, especially if 15N is applied as a single application. If enrichment declines with time, then comparisons of legume varieties with different life cycles may not be valid (Danso et al. 1993). Jorgensen et al. (1999) suggested that the method could be improved by immobilising the 15N using sucrose and straw, as this caused the 15N enrichment to decline more slowly over the growing season. Errors associated with the reference plant are greatest at low levels of fixation (Danso, et al. 1993). For accurate results from this method, the legume and reference crop should also absorb similar ratios of fertiliser N and soil N during each phase of their life cycle. It is also necessary to assume that the legume and non-legume have identical patterns of nitrogen uptake throughout the growing season, and that both have access to the same sources of soil nitrogen (Bremer et al., 1993). The non-legume monocultures used for estimates of nitrogen transfer should ideally be as close to the mixed plots as possible, because of soil spatial variation (Reichardt et al., 1987).
Usually only herbage above stubble height is sampled (e.g. Høgh-Jensen and Schjoerring, 1994). Sometimes a correction is made for nitrogen present in stolon and root material (Jorgensen and Ledgard, 1997), and this is assumed to be constant, although there seems to be disagreement about this:
Jorgensen and Ledgard (1997) and McNeill and Wood (1990) found that estimates of the proportion of nitrogen derived from the atmosphere (pNdfa) were the same whether roots and stolons were included in the calculations or not. However, estimates of nitrogen transfer are changed if changes in root and stubble N are taken into account (Laidlaw et al., 1996).
There is some phenotypic variation in clover root:shoot dry matter ratios (Burdon, 1980). Fertiliser nitrogen also affects root:shoot ratios (Jorgensen and Ledgard, 1997; Ryle et al. 1981a), as does defoliation (Davidson et al. 1990, Pedersen, 1989); defoliation can also cause an increase in the proportion of soil derived nitrogen in clover roots (Marriott and Haystead, 1992) and a decrease in root and stolon mass (Chapman and Robson, 1992).
Legumes are also known to store nitrogen in their root systems over winter (e.g. Li et al., 1996), so root nitrogen may not be constant throughout the growing season. Hay (1985) observed that the percentage of stolon below ground, at the surface and above ground, varied with season: In midsummer, surface and above ground stolon material constituted around 50% of the total stolon weight, but at midwinter, around 90% of the stolon material was below ground. All of these factors suggest that root stolon and stubble sampling could be necessary to obtain accurate measurements of nitrogen fixation.
These techniques work by measuring the levels of N-containing compounds transported from the roots to the shoots in xylem sap. When this differs between fixing and non-fixing varieties of the same species of legume, it can be used as a measure of nitrogen fixation (Hardarson and Danso, 1993). Sap can be extracted from cut stems using a vacuum pump, or from cut roots, by root pressure. Extracts can also be taken from stems and petioles using hot water (Herridge and Peoples, 1990). This technique is most useful for studying tropical legumes, such as soybean and cowpea, which transport N as ureides, allantoin and allantoic acid. Other legumes transport fixed N as amides, asparagine and glutamine, and show less response to changes in N source in this respect (Hardarson and Danso, 1993). So far this technique has not been used with T. repens.
Measurements of the level of ureide in the sap of soybean plants has been shown to be highly correlated with nitrogen fixation estimates obtained by 15N labelling, with different strains of soybean and rhizobium (Herridge and Peoples, 1990).
If a legume is grown in an atmosphere enriched with 15N2, direct measurements of pNdfa can be made. This requires an enclosed, airtight system, which makes this method impractical for use in the field. The volume of the system must be small enough to allow sensitive measurements to be made, and to keep the quantity of 15N2 (and therefore the cost) to a minimum (Warembourg, 1993). Problems could arise in a small system if oxygen became depleted, or if CO2 accumulated at higher levels than would occur in nature. Either of these things could disrupt the normal pattern of plant growth. In long term experiments of this sort, the atmosphere inside the system must be continually circulated and regulated by the addition of oxygen or nitrogen. CO2 can be removed if necessary by a soda trap. Atmospheric pressure must also be regulated (Warembourg, 1993). Results from this method are similar to those from the 15N-dilution method and the total N difference method (McNeill et al., 1994).
Advantages and disadvantages of the various methods of measuring N fixation are summarised in Table 1.4.1. Judging by the published evidence, the N difference, 15N dilution and natural abundance methods are most suitable for measurement of N fixation in the field over long time periods. These three methods can all provide estimates of N transfer from legumes to non-legumes, but the N difference method cannot be used to estimate pNdfa.
|
Method |
Time-scale |
Sources of error |
Can plants be grown in the field? |
Ever inconsistent with other methods? |
Cost |
|
N difference/ fertiliser equivalent |
No time limit |
Nitrogen sparing effect 8 |
yes |
Acetylene reduction 6 , 15N dilution, Natural abundance 2 |
Cheap |
|
Acetylene reduction |
hours |
Ethylene producing bacteria, Inhibition of rhizobium 10 |
yes |
15 N dilution and N difference methods 6 |
Cheap |
|
Natural abundance |
No time limit |
Background variation, contaminated samples, reference crop 4 |
yes |
15 N dilution 5, N difference 2 |
expensive equipment |
|
15 N dilution |
No time limit |
reference crop, soil variation variable root mass 1, 3, 7, |
yes |
Acetylene reduction 6, N difference 2 |
expensive |
|
Ureide assay |
short-term |
- |
yes |
- |
cheap |
|
15 N2 Incubation |
<year |
atmospheric composition, leaks 9 |
no |
no |
expensive |
Table .1 Methods of measuring N fixation
References: 1Bremer et al. (1993) 2Carranca et al. (1999) 3Danso et al. (1993) 4Handley (1996) 5Høgh-Jensen and Schjoerring (1994) 6Martensson and Ljunggren (1984) 7Reichardt et al. (1987) 8Senaratne and Hardarson (1988) 9Warembourg (1993) 10Wood (1995)
Estimates of N fixation by white clover have been summarised from available data (Table 1.5.1). Generally, N fixation by white clover is highest when no nitrogen is applied, and declines in proportion to the amount of N fertiliser applied (Figure 1.6.1). Average estimates of N fixation with low N (less than 5 kg N fertiliser ha-1 yr-1) are 161.6 kg N ha-1 in New Zealand, 145.1 kg N ha-1 yr-1 in the British Isles, and 130.7 kg N ha-1 yr-1 in Denmark. In Switzerland and the Netherlands, the average N fixation values are 215.25 kg N ha-1 yr-1 and 359.6 kg N ha-1 yr-1 respectively, but these values should be taken with caution as the data for these countries came from a single source each. When only data from 15N dilution studies is included, average N fixation by white clover is 158.3, 165.7 and 120.6 kg N ha-1 yr-1 for New Zealand, Britain and Denmark respectively. There was no clear evidence from this that different methods of estimating N fixation produced different results, but other factors such as N application, and the differences between sites made comparisons different.
|
N fixed (kg ha-1) |
Measurement technique |
fertilizer(kg ha-1) |
Location |
Ref. |
|
83.5-171 |
15 N dilution |
0 |
UK |
20 |
|
82-291 |
15 N dilution |
0 |
NZ |
17 |
|
82-213 |
15 N dilution |
0 |
NZ |
18 |
|
40-160 |
15 N dilution |
390 |
NZ |
18 |
|
27-122 |
15 N dilution |
0 |
UK |
7 |
|
-89-178 |
N difference |
0 |
UK |
7 |
|
158-195 |
15 N dilution |
0 |
Denmark |
13 |
|
42-200 |
15 N dilution |
0 |
Uruguay |
19 |
|
0-20 |
15 N dilution |
0 |
U.S.A |
10 |
|
83-283 |
15 N dilution |
0 |
Switzerland |
1 |
|
48-173 |
15 N dilution |
120 |
Switzerland |
1 |
|
165-211 |
15 N dilution |
150 |
Switzerland |
1 |
|
71.82-109 |
15 N dilution |
0 |
NZ |
16 |
|
184-232 |
Acetylene reduction |
0 |
NZ |
12 |
|
105 |
15 N dilution |
0 |
NZ |
26 |
|
71-114 |
15 N dilution |
3 |
Denmark |
11 |
|
54-90 |
15 N dilution |
24 |
Denmark |
11 |
|
52-85 |
15 N dilution |
48 |
Denmark |
11 |
|
47-78 |
15 N dilution |
72 |
Denmark |
11 |
|
150-545 |
N difference |
0 |
Netherlands |
6 |
|
114.9-233 |
15 N dilution |
1 |
UK |
15 |
|
146-167 |
acetylene reduction |
62.5 |
UK |
22 |
|
30-50 |
acetylene reduction |
0 |
UK |
2 |
|
49 |
acetylene reduction |
0 |
UK |
24 |
|
66-81 |
acetylene reduction |
0 |
Canada |
25 |
|
83-296 |
Acetylene reduction |
0 |
Eire |
21 |
|
76-105 |
Acetylene reduction |
0 |
NZ |
4 |
|
211-242 |
Acetylene reduction |
0 |
NZ |
3 |
|
45-142 |
15 N dilution |
3.6 |
NZ |
5 |
|
268 |
acetylene reduction |
0 |
UK |
9 |
|
191 |
N difference |
0 |
Denmark |
14 |
|
152 |
N difference |
78 |
Denmark |
14 |
|
115 |
N difference |
155 |
Denmark |
14 |
|
69 |
N difference |
310 |
Denmark |
14 |
|
38 |
N difference |
465 |
Denmark |
14 |
|
208 |
N difference |
125 |
Denmark |
23 |
|
143 |
N difference |
250 |
Denmark |
23 |
|
90 |
N difference |
375 |
Denmark |
23 |
|
74 |
N difference |
500 |
Denmark |
23 |
Table .1 N fixation by white clover in different parts of the world and at different levels of N fixation
References: 1Boller & Nosberger, 1987 2Bradshaw et al. 1975 3Clark et al., 1979 4Crush et al. 1983 5Edmeades & Goh, 1978 6Elgersma & Hassink 1997 7Goodman, 1988 9Halliday and Pate, 1976 10Heichel & Henjum, 1991 11Høgh-Jensen & Schjoerring 1997 12Hoglund & Brock, 1978 13Jorgensen et al. 1999 14Koefoed & Klausen, 1969 15Laidlaw et al., 1996 16Ledgard et al. 1987 17Ledgard et al., 1990 18Ledgard et al., 1996 19Mallarino et al. 1990 20McNeill & Wood, 1990 21Masterson & Murphy, 1976 22Palmer & Iverson, 1983 23Pedersen & Moller, 1976 24Skeffington & Bradshaw, 1980 25Vessey & Patriquin, 1984 26Wheeler et al. 1997)
As we have seen from section 1.5, N fertilisers significantly reduce N fixation by white clover and other legumes. Figure 1.6.1 gives an indication of the scale of this effect. When N is applied to a legume growing as a monoculture, the legume usually increases its uptake of soil N, and consequently the pNdfa (see section 1.4.4) is reduced. Application of fertiliser N causes a reduction in the number and mass of root nodules (Cowling, 1961). This is probably because uptake of soil N requires less photosynthetic energy than N fixation, and so under conditions of abundant N, the relationship between the rhizobium and its host legume will become increasingly parasitic, rather than mutualistic. The rhizobium is benefiting from its association with the legume, but the legume is not. Similar shifts between mutualism and parasitism have been observed in mycorrhizas (symbiotic plant root fungi, Johnson et al., 1997). Parsons et al. (1993) suggest that the effect of soil nitrogen upon pNdfa, may be caused by a feedback mechanism, in which nodule growth and activity are inhibited by high levels of phloem nitrogen compounds, probably amino acids.
Bergersen et al. (1989) observed that cereal crops grown after ploughing of grassland and before planting of soybeans, reduced available N in the soil. This had the effect of increasing N fixation by the soybeans and also improved yields and protein content of the soybean crop. Davidson and Robson (1986) showed that continuous exposure to low levels of nitrogen reduced pNdfa more than short-term exposure to high concentrations of nitrate. Growing legumes under conditions of high N is not making good use of natural resources. pNdfa will be reduced by high soil N, and there is likely to be a high risk of N leaching especially if the legumes are grown as monocultures (Webster et al. 2003). In organic systems there are also likely to be increased weed problems because of the high availability of N.
When N is applied to mixtures of legumes and non-legumes (such as grass-legume leys), the growth of the non-legume is commonly increased at the expense of the legume (Frame and Newbould, 1986, Danso et al. 1988, Waterer et al., 1994, Stewart and Chestnutt, 1974, Rys and Mytton, 1985, Høgh-Jensen and Schjoerring, 1994 & 1997). This seems to indicate that under conditions of high nitrogen, legumes suffer in competition for light, water, and soil nutrients other than N. Laidlaw and Withers (1998) suggest that in grass-clover swards, applications of N cause grass to shade out clover. Davidson and Robson (1986) observed that under laboratory conditions, high levels of soil N increased grass dry matter and reduced clover dry matter in mixed swards. The N concentration of the clover (N concentration in clover dry matter) was unaffected by N fertiliser, and pNdfa was reduced by N fertiliser inputs indicating that under these conditions, clover was able to compete with grass for soil N. Herrmann et al. (2001) also observed reduced clover yield and pNdfa when N fertiliser was applied at a rate of 80-160 kg N ha-1. Applications of fertiliser N did not affect the N concentration of the clover herbage (Herrmann et al., 2001). Similar effects have been observed in many other studies e.g. Ledgard et al. (2001), Høgh-Jensen and Schjoerring (1994). In all these examples, N fertiliser caused a reduction in the ratio of clover to grass and a decline in pNdfa. Clover N concentration was in all cases higher than grass N concentration and was unaffected by fertiliser N. This indicates that clover is capable of competing with grass for soil N, although under conditions of high soil N, other competition factors become important.
The effect of N fertiliser on N fixation in ryegrass-white clover swards is shown in Figure 1.6.1. Data was compiled from a number of experiments performed in Europe and New Zealand. There is considerable variation between sites, as would be expected, but overall, N fixation is reduced by approximately 25 kg N ha-1 for every 100 kg N ha-1 of fertiliser N applied.
Although N fertiliser generally benefits grass at the expense of clover, it has been suggested that small amounts of nitrogen improve establishment of clover, by delaying nodulation (e.g. Young, 2002). A review of the evidence by Peoples et al. (1995) found that in most cases, even small amounts of N fertiliser suppressed N fixation.
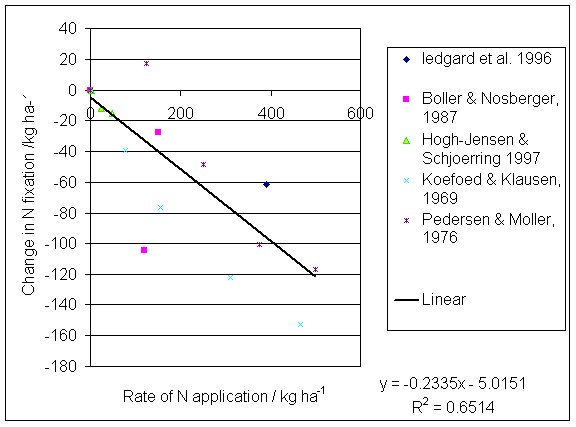
Figure .1 Effect of N fertiliser applications on N fixation by white clover-grass leys.
Data from 1Ledgard et al. 1996, 2Boller and Nosberger, 1987 3Høgh-Jensen & Schjoerring 1997, 4Koefoed & Klausen, 1969, 5Pedersen & Moller, 1976
Grazing removes N from large areas and transfers it to small, localised patches (Schwinning and Parsons, 1996a). Afzal and Adams (1992) found dramatic variations in nitrate and ammonium on cattle grazed pastures, over distances of only 100 mm. This small-scale variation was attributed to individual dung and urine patches. The patchy distribution of nitrogen in grassland caused by grazing could theoretically increase clover growth, by creating low N microsites, in which clover could thrive. Hoglund and Brock (1978) found that grazing depresses nitrogen fixation, probably because of the effects of dung and urine patches (Marriott et al., 1987a) and defoliation of clover (Farnham and George, 1994). Baars and Brands (1996) showed that composted farmyard manure inhibited clover stolon growth less than slurry, but that this effect varied with clover varieties. Jorgensen and Jensen (1996) found no effect of dung upon white clover growth or fixation in the first four weeks after application. This was attributed to the slow release of nitrogen from dung. Vinther (1998) estimated that dung and urine patches reduce N fixation in their immediate vicinity by around 15%. Weeda et al. (1967) observed that clover was more prevalent on cattle dung patches than other species, especially in winter. Clover was quick to colonise dung patches, and remained growing on the patches for approximately 1.5 years after deposition (Weeda et al., 1967). In contrast, Lieth (1960) observed that sites where dung had been deposited, became colonised by species such as Poa trivialis (rough meadow grass), Agrostis alba (wood meadowgrass) and Ranunculus repens (creeping buttercup) at the expense of T. repens, although the study appears to have been much less extensive than that of Weeda et al. (1967). Urine + faeces and urine alone decreased the clover content of swards, but faeces alone might actually benefit the clover (Weeda, 1967). Dung and urine generally contain virtually all of the nutrients necessary for plant growth, in varying quantities. It is probably misleading to think of dung and urine simply as sources of N. The effect of dung and urine on clover growth and N fixation could well depend upon the local soil conditions and the availability of key nutrients.
Wilde et al. (1983) compared rotational grazing to continuous stocking. After one growing season there was over 50% clover on the rotationally grazed plots, while set stocked plots had 5-7% clover. The study concluded that rotational grazing was better than continuous stocking, because it relieves clover from selective defoliation and allows the clover petioles to elongate, placing leaves in the top of the canopy.
Schils et al. (1999) found that white clover ground cover was greater in a system with cutting but no grazing, than in grazed systems. A rotational system of cutting and grazing reduced clover cover by 12%. The negative effect of grazing was most marked from July and August onwards. Where two silage cuts were taken rather than one, clover cover was on average 8% higher. However, dry matter and N yields of rotationally grazed plots were equal to or higher than those which were cut only, suggesting that return of N in dung and urine compensated for the reduced percentage of clover. Rhodes (1984) states that clover yields from rotationally grazed swards are lower than yields from cut swards. Frame and Newbould (1986) observed that grazing depressed clover content of swards and total production, and that clover performance was greatest in cut swards when the interval between cuttings was increased.
Laidlaw and Stewart (1987) found that maximum clover content of a pasture rotationally grazed by cattle, declined from 55% in the third year, to 24% in the sixth year. Hard grazing by sheep over the next three winters increased the clover content to 34 or 45%, depending upon whether or not nitrogen was added in spring. The corresponding figures for a control plot were 7 and 11%.
Sheep generally preferentially graze clover in grass-clover swards, and this may reduce N fixation. Goats will graze weeds and grass in preference to clover, and can increase the clover content of a sward (Grant et al., 1984).
Trampling and soil compaction caused by grazing animals may also reduce N fixation and clover content of pastures (Curll and Wilkins, 1983).
Acuna and Wilman (1993) noted that clover content in a sward was affected by cutting height: Cutting close to the ground encouraged clover, whereas cutting at 100 mm for several years almost completely eradicated clover. The authors explained the suppression of clover with increased cutting height as a shading effect (clover is a low growing plant compared to many grasses). Another possibility is that close cutting removes more nutrients from the soil than lax cutting, and this could also affect the competition between grass and clover. Experiments comparing regular and infrequent cutting of grass-clover swards, with and without applied N, show that less regular cutting of a grass-clover sward reduces clover yield only when N is applied (Wilman and Fisher, 1996), so it is probably true to conclude that grass often out-competes clover for light, when N is applied, or when the sward is allowed to grow tall. There seems to be a general agreement from all of these studies that cut swards favour clover more than grazed swards, and that clover yields better in rotationally grazed swards than in continuously grazed swards.
White clover in leys is seldom grown as a monoculture. More usually it is grown with one or more non-leguminous species, or companion species. Ryegrass is generally considered to be a good companion species for white clover, whereas cocksfoot (Dactylis glomerata) is less compatible (Chestnutt and Lowe, 1970). Bent grasses (Agrostis spp.) and Yorkshire fog (Holcus lanatus) are also considered poor companion species for clover (Frame, 1990). In contrast, Edmond (1964) found that the yield of clover grown with Holcus lanatus and Agrostis tenuis was actually slightly higher than yield of clover grown with Lolium perenne. Clover grown with D. glomerata yielded about half that of clover grown with L. perenne (Edmond, 1964)
Williams et al. (2000) compared yields of two medium leaf sized clover varieties (AberDai and AberVantage), grown individually and as mixtures. The clover was grown with four ryegrass varieties. Two of these (Augusta and AberOscar) were tetraploid hybrids between perennial ryegrass and Italian ryegrass (Lolium multiflorium L.) The other two varieties were a diploid Italian ryegrass (AberComo) and a tetraploid perennial ryegrass (Merlinda). The management involved silage cuts and grazing by sheep and cattle each year for four years. Overall there was no benefit from mixing the two clover varieties, but in most years, clover yields were highest with Merlinda (the lowest yielding grass) and lowest with AberComo (the highest yielding grass). This experiment only looked at dry matter yields, and not N yields or N fixation.
Mattner and Parbery (2001) showed that when perennial ryegrass became infected with crown rust, it suppressed the growth of clover more than healthy ryegrass. Soil previously growing rusted ryegrass and leachate from soil growing rusted ryegrass also suppressed clover growth. This was explained as an allelopathic effect.
Competition between clover and grass seems to be generally poorly understood. Competition between clover and other pasture species could depend on light, temperature, water or availability of nutrients, and the ability of the other species in the sward to compete for these resources. A grass variety or species which out-competes clover in the short term is likely to reduce the productivity of the sward in the long term. The species in a sward must effectively utilise soil nutrients to prevent leaching losses. On the other hand, if they compete too aggressively with clover, the productivity of the sward will be reduced in the long term.
Hoglund and Brock (1978) noted large differences in nitrogen fixation by white clover with season, and also between years. White clover populations in particular are claimed to "crash" every few years (Fothergill et al., 1996).
Schwinning and Parsons (1996a) suggest that clover and grasses replace each other cyclically at the level of individual patches. Grass has a competitive advantage when soil N is high, and clover is at an advantage when nitrogen levels are low, but clover gradually elevates the level of soil nitrogen locally. They modelled this by representing a pasture as 90,000 interlocking hexagonal cells. These cells could be in one of 4 states: legume dominant, grass dominant (with legume present), pure grass at high soil N and pure grass at low soil N. Cells moved between these states in response to urine deposition, N enrichment by clover, N depletion by grass, invasion by clover and extinction of clover. The output of the model suggested that this would lead to cyclical variation in clover content with a period of 3-4 years (Schwinning and Parsons 1996a). Evidence from field data provided some support for this (Fothergill et al., 1996, Schwinning and Parsons, 1996b). If clover is uniformly distributed, then clover in all parts of the pasture will oscillate in phase. However, clover dies back in winter and must re-invade the areas where it previously occurred, during the rest of the year. This raises the possibility that some patches of clover will disappear altogether, especially when other factors such as grazing, trampling, pests, dung and urine are considered. These processes will increase patchiness. As the pasture becomes more patchy, different regions will no longer be in phase with one another, so oscillations at the field scale will be damped down. Grass yield may be more closely correlated with previous years clover yield than with current clover growth, so that clover cycles and total biomass cycles are not in phase. The amount of dieback is related to climate, and to management (field scale effects) where there is very little dieback, the pasture will take many years to reach equilibrium, a lot of dieback and the pasture reaches equilibrium quickly. (Schwinning and Parsons, 1996a).
Turkington and Harper (1979) suggested the following cyclical succession sequence: T.repens would invade and be joined by L. perenne, the two species would coexist, because of the high nitrogen requirements of L. perenne and the asynchronous growth cycles of the two species. As the soil nitrogen level rose owing to the nitrogen fixation by T.repens, T.repens would go into decline, because of its poor ability to compete for soil nitrogen, and be replaced by Alopecurus pratensis (meadow foxtail) and/or Dactylis glomerata. L. perenne would also decline at this point. The nitrogen level in the soil would then decline and A. pratensis would be replaced by slower growing species with a low nitrogen demand, such as Anthoxanthum odoratum (sweet vernalgrass) and Agrostis capillaris. They also suggest that nitrogen inputs from dung and urine would complicate the picture, by effectively omitting clover from the sequence locally.
Experiments with simulated swards containing a number of grassland species, and observations of old pasture, by Thorhallsdottir (1990a,b) did not support the idea of simple cyclical species replacement, although some patterns of replacement related to T. repens were observed. T.repens was more likely to replace certain species, and be replaced by others, than could be expected from chance. T.repens had a tendency to colonise gaps, and to be replaced by gaps. It also moved rapidly through the pasture, never occupying the same space in successive years more often than would be expected by chance (Thorhallsdottir, 1990a). Cain et al. (1995) observed changes in clover density in a lawn between years, and a general pattern of moving clover patches in a sea of grass. Cyclical replacement of grass species by clover was observed (different grass species were not distinguished). The larger patches of clover persisted between one and three years. Some small patches might have persisted for longer than the 4 years of the study. Lieth (1960) also noted cyclical species replacement in grassland, but failed to describe it in any detail.
Climatic factors such as sunlight, temperature and rainfall also contribute to yearly variation in clover growth (Frame and Newbould, 1984). Hay et al. (1990) note that clover populations consist of a few large individuals, and a large number of small individuals (a result of senescence of stolons). These small individuals could easily die off under harsh environmental conditions, pests, disease etc. Frame and Newbould (1984) also suggest that inappropriate herbicide use might be partly responsible for some clover crashes.
Edmeades and Goh (1978), looked at pastures 2, 6, 15 and >20 years old, and found that nitrogen fixation generally decreased with age of pastures, although this study had no replicates. In contrast, Heichel and Henjum (1991) found that nitrogen fixation in forage legumes increased with the age of the pasture. Some of these differences could have been due to variations from year to year. Høgh-Jensen and Schjoerring (1997) showed that at low seeding densities, clover could take a full growing season to achieve the same yield output as an initially well-seeded sward. Kristensen et al. (1995a) estimated that a pasture containing 30% clover would average 190 kg N ha-1 yr-1 in the first two years, and 128 kg N ha-1 yr-1 in subsequent years.
At low temperatures, clover is less able to fix nitrogen, and suffers through competition for soil nitrogen with grass (Nesheim and Boller, 1990; Prevost and Bromfield, 1991). Macduff and Dhanoa (1990) also found that temperatures below 13oC suppress nitrogen fixation. Frame and Newbould (1986) suggest that N fixation by white clover requires a temperature of about 9o C. At low temperatures, plant growth as a whole is reduced, and so the reduced fixation could simply be a response to low nitrogen demand. Ollerenshaw and Baker (1981) observed that clover roots remained active at temperatures as low as 5oC. Nitrogen fixation is therefore likely to be affected by seasonal and annual temperature variations, as well as seasonal and annual variations in water availability and soil factors.
Different legume species (Heichel and Henjum, 1991) and cultivars (Ledgard et al., 1990, 1996) fix different amounts of N, and this may be related to their tolerance of soil nitrogen levels (Ledgard et al. 1996).
In the laboratory, water stress reduces nitrogen fixation (Engin and Sprent, 1973), but in the field, drying of the surface soil layers causes nitrogen fixation by T.repens to take place at greater depths, reducing this effect (Hoglund and Brock, 1978).
Topography also seems to be important: sloping sites tend to have lower N fixation, probably because of differences in microclimate and soil fertility (Ledgard et al., 1987), and lowland sites can have four times as much N fixation as upland sites, probably because of the longer growing season (Goodman, 1988). Jacot et al. (2000) observed that at altitudes over 2100m above sea level, T.repens was not present, although Trifolium pratense (red clover) did grow at this altitude and Lotus corniculatus (birdsfoot trefoil) and Trifolium alpinum (alpen klee) could grow at altitudes of 2300m and 2600m respectively. High altitude did not reduce
pNdfa values of any of the species in the study, even at the limits of their range, despite low temperatures and acid soils.A survey of dairy and beef farms by Forbes et al. (1980), showed that clover content of swards was significantly affected by soil drainage, fertiliser N use, whether or not clover was a preferred species, the potential for transpiration from the crop surface and the number of days of drought per year. Soil pH and available soil P and K had no significant effect on clover growth. In contrast, Snaydon (1961) observed that clover distribution in hill pastures was related to levels of Ca and P.
Giller and Cadisch (1995) looked at ways of increasing biological nitrogen inputs to world agriculture. They concluded that in the short term improvements could be made by improving soil conditions, such as acidity, water stress, nutrient deficiencies and high soil nitrogen, using liming, fertiliser, green manure and crop rotation. Simply encouraging the wider use of legumes would also be effective. They commented that: "Immediate dramatic enhancements in input from N2 fixation are possible simply by implementation of existing technical knowledge". Inoculation of legumes with appropriate rhizobium strains would have immediate benefits in many parts of the world. Breeding of improved legume varieties and rhizobium, and genetic engineering, will only show benefits in the longer term, if at all (Giller and Cadisch, 1995).
Some of these factors have been quantified experimentally, and these are shown in Table 1.8.1. It is clear from this study that much of the research on clover has focussed on the effects of N in fertiliser and in dung and urine, and this seems to cause consistently large reductions in N fixation. Some of the factors, such as clover variety, irrigation and management may not be universally applicable: different clover varieties may perform differently at different sites, and certain management techniques may be more appropriate in some areas than others. Table 1.8.1 only records available data from studies on N fixation. It does not include studies that have recorded factors affecting clover yield only, even though this is likely to have highly significant effects on N fixation (e.g. Acuna and Wilman, 1993). Some of the factors observed to inhibit nitrogen fixation might simply be reducing plant growth as a whole (Hartwig and Nosberger, 1996)
. Some factors, such as grazing management, are extremely difficult to examine in N fixation studies, because much of the fixed N in clover herbage in a grazed sward is likely to be consumed by the grazing animals.
|
Factor |
Scale of effect (% change) |
Reference |
|
Soil nitrogen (16 mg N/plant) establishing sward |
94.9% |
10 |
|
Soil nitrogen (8 mg N/plant) established sward |
58.33% |
10 |
|
Soil nitrogen (465kg ha-1 yr-1) |
80.1% |
5 |
|
Soil nitrogen (400 kg ha-1 yr-1) |
57.86% |
3 |
|
Soil nitrogen (390 kg ha-1 yr-1) |
57.6% |
6 |
|
Soil nitrogen (310 kg ha-1 yr-1) |
63.9% |
5 |
|
Soil nitrogen (155 kg ha-1 yr-1) |
39.79% |
5 |
|
Soil nitrogen (78 kg ha-1 yr-1) |
20.42% |
5 |
|
Soil nitrogen (72 kg ha-1 yr-1) |
27.7% |
4 |
|
Soil nitrogen (48 kg ha-1 yr-1) |
18.1% |
4 |
|
Soil nitrogen (24 kg ha-1 yr-1) |
14.5% |
4 |
|
4 cuts (compared to 3) |
-41% |
8 |
|
5 cuts (compared to 3) |
-46% |
8 |
|
Sandy soil (in comparison with clay soil) |
7.7% |
2 |
|
3rd/4th/5th years pasture (compared to 1st & 2nd years) |
34.6% |
2 |
|
Irrigation |
-15.4% |
2 |
|
Clover variety (Kopu vs Sabeda) |
60.8% |
6 |
|
Pattern of nitrogen supply |
18.79% |
1 |
|
Waterlogging |
97% |
9 |
|
Temperature (5-15oC) |
98.35% |
7 |
|
Dung and urine patches |
10-15% |
11 |
Table .1 Factors reducing nitrogen fixation in white clover leys
References: 1Davidson and Robson (1986) 2Gregersen (1980) 3Høgh-Jensen and Schjoerring (1994) 4Høgh-Jensen & Schjoerring (1997) 5Koefoed & Klausen (1969) 6Ledgard et al. (1996) 7Nesheim and Boller (1990) 8Pedersen and Moller (1976) 9Pugh et al. (1995) 10Rys & Mytton (1985) 11Vinther (1998)
Like any plant species, white clover has a niche, a set of environmental conditions, in which it can grow and compete with other plants for soil nutrients, water and sunlight. The fact that white clover is found growing in so many different parts of the world, and habitats, suggests that it has a fairly broad niche, and some varieties may be adapted to local conditions. Although white clover has been cultivated and bred, the cultivated forms still resemble those that are found in the wild, and the cultivated varieties can themselves grow in the wild. This is probably because many of the grasslands, in which clover is grown could be described as semi-natural habitats. Clover has the ability to fix nitrogen and spreads rapidly, and this suggests that it may sometimes behave in the wild as a pioneer species, rapidly colonising disturbed and bare ground. Pioneer species are generally transient, and replaced by other species in a successional cycle. This may explain why clover yields can fluctuate from year to year. The fact that clover elevates soil N while at the same time being vulnerable to high soil N, may also affect its growth and N fixation over time.
In order for clover to yield well, the conditions on the farm must resemble to some degree the conditions for which clover has evolved. In addition, for clover to fix nitrogen effectively, it requires a good supply of all nutrients besides nitrogen. A surplus of soil nitrogen reduces the ability of clover to compete with grass. Competition between clover and grass under these circumstances must depend on factors other than N. In Britain, there is some evidence that one of the most important factors affecting the balance of competition between clover and grass is light. Grass species such as
L. perenne grow taller than clover, and under high N conditions are able to grow rapidly and shade out clover. Close grazing or cutting of grass clover swards could reduce the shading of clover by grass, and thus mitigate the effects of high soil N. Temperature and management may also interact with soil N. Clover grows less well than grass at low temperatures and so a high level of soil N in spring and autumn could severely suppress the growth of clover at a time when it is vulnerable to competition from grass.
The overall aims of this study were to quantify the extent and scale of variation of nitrogen fixation in organic white clover/ryegrass leys, and to identify the causes of this variation. The study aims to measure and compare the effects of season, crop rotation, soil N and competition on N fixation and growth of white clover. In particular, the study aims to test a number of hypotheses about the relationship between availability of soil nitrogen and grass- clover competition, which will be described in section 2.1.
Specific objectivesSeveral authors (Schwinning and Parsons, 1996a; Fothergill et al. 1996; Turkington and Harper, 1979, Lieth, 1960) have suggested that white clover and grass may be responding to natural variations in soil N, and also contributing to this variation. These authors are broadly agreed on a set of ideas that will be referred to in this study as cyclical replacement. Cyclical replacement makes the following assumptions:
Some of the assumptions underlying cyclical replacement are well established from laboratory and field studies, but a number of questions must be answered before it can be shown that cyclical replacement is actually happening in the field:
One of the purposes of a ley in organic farming systems is to build soil fertility for the benefit of successive crops (section 1.2.2). If fixed nitrogen is accumulating in the soil over the course of the ley phase, this would be expected to affect the growth of grass and clover. If cyclical succession is taking place, changes in soil N under clover and grass microsites should be observable in the field. Invasion of microsites by clover and grass should also be influenced by the availability of soil N. It was therefore hypothesised that:
This part of the study consisted of two main field experiments: a study of N fixation and N transfer by white clover and N uptake by grass (Section 4 N fixation and grass-clover dynamics) and a study of soil fertility (Section 5 Soil chemistry of grass-clover swards). The field experiments were carried out on two experimental organic farms managed by the Scottish Agricultural College. The two farms contained experimental ley-arable rotations, which had been established several years previously in order to compare the effects of rotations with different ratios of grass-clover ley to arable crops. Although the two farms in this study are in the same broad region (North-Eastern Scotland), they have distinctly different climate and soil fertility.
The Tulloch Organic UnitThe Tulloch organic farm is located at the SAC’s Craibstone Estate, Aberdeen (Latitude N 57o 10’ Longitude W2 o14’, National Grid reference NJ843094). The farm covers 65.8 ha of exposed, marginal land, 160 m above sea level, of which 21.9 ha are arable land, 37.7 ha are permanent grazing and 6.2 ha are trees and buildings. The soil type is a sandy loam of the Countesswells series (leptic podzol in FAO classification). In 1997 at the start of sampling, soil pH averaged 5.8 and the soil contained 9.5% organic matter. Soil nutrient levels were moderate to high. P, K and Mg levels in extracts were 15, 97 and 88 mg l-1, respectively (M.Coutts pers. comm.). In 1992, the Soil Association certified the farm organic.
A rotational trial was established on the farm in 1991. The trial comprises two rotations each replicated twice. Plots are 26 ´ 30m (0.078 ha and the layout is shown in Figure 3.1.1.
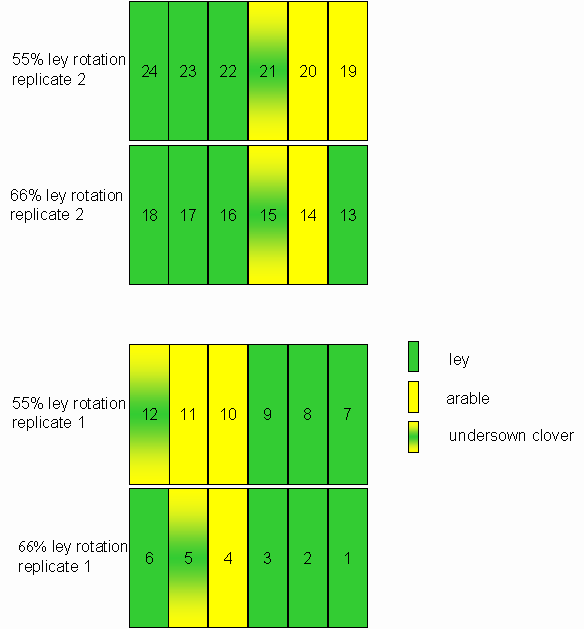
Figure .1 Layout of the trial rotation plots at Tulloch in 2000.
The rotations are referred to as the 66% ley rotation and the 50% ley rotation. The cropping sequences are shown in Table 3.1.1
|
Cropping sequence |
||||||
|
66% ley |
Ley (grazed) |
Ley (cut) |
Ley (grazed) |
Ley (cut) |
Oats |
Oats (u/s) |
|
50% ley |
Ley (grazed) |
Ley (cut) |
Ley (cut) |
Oats |
Swedes |
Oats (u/s) |
Table .1 Cropping sequences for the 66% ley rotation and the 50% ley rotation at Tulloch. u/s = undersown with grass and clover
The final year of oats in each rotation is undersown with perennial ryegrass (var. Condessa), Timothy (var. Scots) and White clover (var. Avoca). Ley plots are grazed rotationally by sheep, stocked at a rate of 1.7 livestock units per forage hectare. Farmyard manure (FYM) from organic livestock on the Tulloch unit is applied to the trial. Manure applications are based on the area of forage in the rotation, an assumed 7.2 tonnes manure available per livestock unit. FYM is only applied to cut leys. FYM is applied in early spring and immediately after the first silage cut when a second silage cut is to be taken. Dates of silage cuts are shown in Table 3.1.2. Application rates for FYM are shown in Table 3.1.3.
|
Age of ley |
1997 |
1998 |
1999 |
2000 |
|
2 (1st cut) |
23.6.97 |
30.6.98 |
30.6.99 |
19.6.00 |
|
2 (2nd cut) |
9.9.97 |
21.9.98 |
5.11.99 |
29.8.00 |
|
3 |
23.6.97 |
30.6.98 |
30.6.99 |
19.6.00 |
|
4 |
23.6.97 |
30.6.98 |
30.6.99 |
19.6.00 |
Table .2 Dates of silage cuts on leys at Tulloch, 1997-2000
|
Age of ley |
rotation |
1997 |
1998 |
1999 |
2000 |
|
2 (1st cut) |
66% |
20 |
20 |
15 |
15 |
|
50% |
15 |
15 |
10 |
15 |
|
|
2 (2nd cut) |
66% |
10 |
10 |
8 |
8 |
|
50% |
8 |
8 |
6 |
8 |
|
|
3 |
50% |
15 |
15 |
10 |
10 |
|
4 |
66% |
20 |
20 |
15 |
15 |
Table .3 Manure applications to cut leys at Tulloch 1997-2000
Management is in accordance with organic standards (Soil Association, 2000a). The trial rotations are in effect a "farm within a farm", and provide a good opportunity to measure N flows under controlled organic conditions. At the time of the start of the study, the trial rotations had undergone one complete cycle, which means that all of the plots in each rotation had had the same number of seasons of ley and arable treatments.
Weather data for Craibstone is shown in Figure 3.1.2, Figure 3.1.3, Figure 3.1.4 and Figure 3.1.5.
The growing season in 2000 was relatively cool compared to other years (Figure 3.1.5). 1999 was dry compared to other years (Figure 3.1.4). Generally the peak of temperature was in August, although in 1999 the warmest month was July, and in 1998 it was September. Rainfall showed no clear seasonal pattern.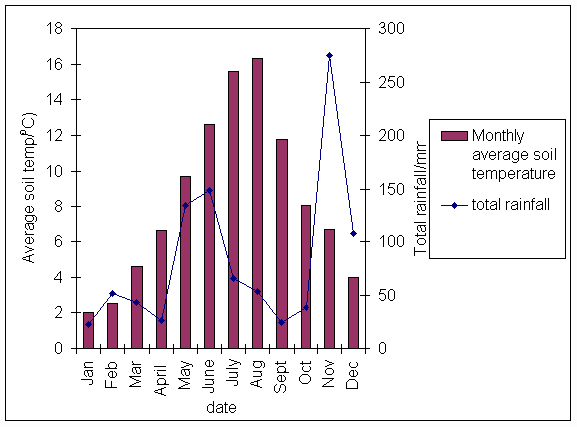
Figure .2 Average monthly soil temperature and total monthly rainfall in 1997.
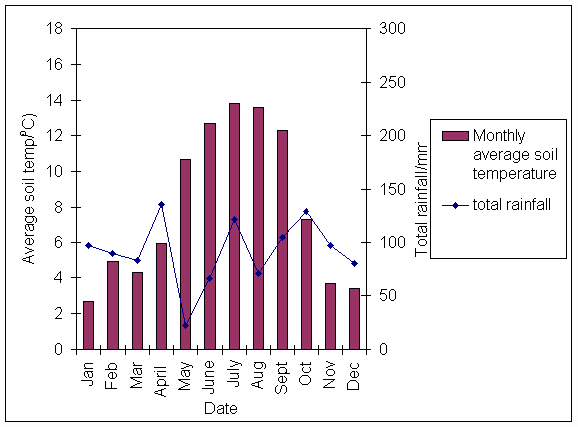
Figure .3 Average monthly soil temperature and total monthly rainfall in 1998
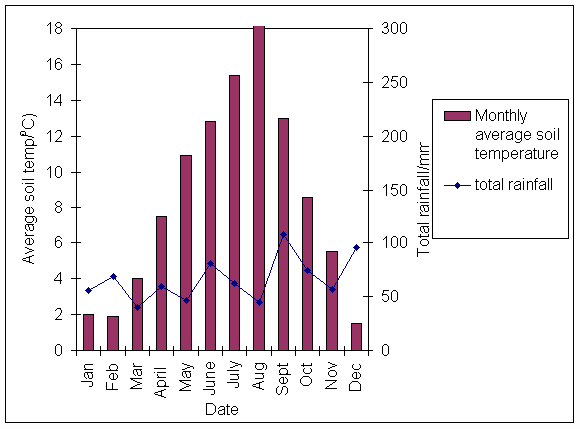
Figure .4 Average monthly soil temperature and total monthly rainfall in 1999
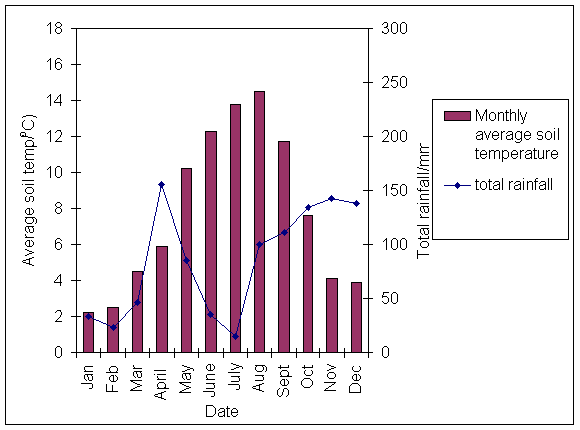
Figure .5 Average monthly soil temperature and total monthly rainfall in 2000
The Aldroughty organic farm is located near Elgin (Latitude N57:38 Longitude W3:23, map reference NJ167625) and covers 56.7 ha of which 43.1 ha are arable land and 13.6 ha are permanent grazing. The site is on sheltered land 25 m above sea level. The soil type is loamy sand/sandy loam. The farm was certified organic by the soil association in 1992.
The farm includes two organic trial rotations, one of which is a replicate of the 50% ley rotation at Tulloch. The plots are 26 ´ 30m (0.078 ha) as in the Tulloch trials, and the leys are managed in a similar way to those at Tulloch. The rotations will be referred to as the 38% ley rotation and the 50% ley rotation. The cropping sequences are shown in Table 3.1.4. Precise weather data for Aldroughty was not available for the period of the experiment.
|
Cropping sequence |
||||||||
|
38% |
Ley |
Ley |
Oats |
Roots |
Oats u/s |
Red clover |
Roots |
Oats u/s |
|
50% |
ley |
ley |
ley |
oats |
Roots |
Oats u/s |
||
Table .4 Cropping sequences for the 66% ley rotation and the 50% ley rotation at Aldroughty. u/s = undersown with grass and clover
This part of the study is concerned with the growth, chemical composition, morphology and yield of clover and grass in different years, ages of ley and rotation. Observations of the relative yields of clover and grass (and their changes over time) give a general indication of the balance of competition between clover and grass. Low total yields of all species, may indicate that some external factor e.g. the weather, is affecting the growth of both species. The isotope dilution experiment allows the quantification of the amount of fixed N present in clover herbage (pNdfa), and in the process provides figures for N concentration of clover herbage and grass herbage. This can give an indication of the importance of N in the grass- clover dynamics. For example, if pNdfa was low, clover N yield was high relative to grass N yield, and grass N concentration was high, this would indicate that clover was successfully competing for soil N. A high pNdfa, high clover N yield, and low grass N concentration would indicate that soil N is limiting grass growth. A low pNdfa, low clover N yield and high grass N concentration would indicate that N is abundant and not limiting the growth of grass, with the result that grass is suppressing the growth of clover due to competition for some other factor. Isotope dilution can also indicate how much N is being transferred from clover to grass over the course of the experiment.
From existing work at Tulloch it was known that clover content of the sward in autumn generally varied between approximately 15 and 45% ground cover (Figure 4.5.22). It is not known to what extent this reflects variation in N fixation, as measurements of pNdfa and N concentration have not been made. There have been few studies looking at N fixation over successive years of a ley arable rotation. Data on the flowering rates, and tap-root disappearance in white clover are lacking, but there is some evidence that flowering is influenced by the availability of light and nutrients (Zaleski, 1970). Flowering rate may be inversely related to persistence in white clover varieties (Williams, 1987). Early disappearance of the tap-root can be prevented by lax grazing and applications of P and K (Westbrook and Tesar, 1955).
AimsThe overall aim of this part of the study was to obtain information about N fixation by white clover in leys, and its relationship with age of ley, competition with grass, crop rotation, soil factors (especially N) and climate.
It was expected that the past history of a plot would influence the level of N fixation. Older leys were expected to have lower N fixation levels than younger leys, because of the accumulation of fixed soil N in the soil over the previous years of ley. Likewise, it was expected that the 66% ley rotation would have lower N fixation than the 50% ley rotation, owing to the greater amount of accumulated soil N resulting from the extra year of ley, and the shorter N depleting arable phase of the 66% ley rotation.
In order to do this, the following factors were examined: clover and grass yield, N concentration in herbage of clover and grass, pNdfa, N fixation and clover flowering rate.
The following hypotheses were tested in this section:
N fixation using 15N dilution technique
1997
In 1997, only the 1-year-old leys at Tulloch were studied (plots 3,10,13 and 19). In each plot, three subplots were chosen, by randomly placing quadrats on the ground. Subplots were covered with exclusion cages (converted lobster pots), to prevent grazing. Three more subplots were chosen in clover-free areas as controls. Subplots were 0.5 m x 0.5 m (0.25 m2). The subplots were cut down to ground level using shears, at the start of the experiment (initial sample) and the herbage was separated into grass and clover. Herbage was separated into grass + weeds and clover, and any soil removed. In a few instances, sheep managed to damage the cages, and partially graze the subplots. When this happened, it was noted and the dry matter yields of these subplots on these dates were not included in the final analysis. The fresh samples were placed in uniform paper bags and weighed, zeroing the balance with an empty paper bag. Samples were then dried overnight at 80 oC, along with an empty paper bag, before weighing again, this time zeroing the scales with the dried paper bag. The dried samples were then ball milled and analysed for their N and 15N concentration on a mass spectrometer (Europa Scientific Tracermass stable isotope analyser).
The standard used to calibrate the mass spectrometer was 2.3584 g of (NH4)2(SO4) (21.21%N, atom% 0.36600), dissolved in 25 ml of deionised water, equivalent to 4.7168 m g in 5m l. Immediately after removal of the herbage, 15N labelled fertiliser was applied to each subplot. The fertiliser was prepared as follows:
For each of the 15 subplots, 589.29 mg of (15NH4)2(SO4) were carefully weighed and mixed with 2 l deionised water in a plastic screw top bottle, and another plastic bottle was prepared with 2 l of deionised water. The (15NH4)2(SO4) had an isotopic enrichment of 10%. In the field, each subplot was watered with 2 l of the 15N mixture from a watering can and subsequently 2 l of deionised water from a separate and clearly marked watering can. This gave an application rate of 5 kg N ha-1 and 0.5 kg 15N
ha-1. A low application rate of highly labelled 15N was used because the sward would not normally receive any artificial N fertiliser. The cutting and labelling procedure was repeated every 28 days, until the end of the growing season. Owing to delays in the delivery of the (15NH4)2(SO4), the initial sample was not taken until 3.7.1997, and only four subsequent cuts were taken. The final labelling was made after the herbage was cut on 23.9.1997, and the final herbage sample was cut on 21.10.1997. Sampling dates for Tulloch are shown in Table 4.3.1.
1998
In 1998, 1 -year-old leys (plots 2, 9, 18 and 24) and 2-year-old leys (3, 10, 13 and 19) were studied (Figure 3.1.1). Three new subplots within each plot were chosen at random. The initial sample of the 1-year-old leys was taken on 29.05.1998, and the initial sample of the 2-year-old leys was taken on 26.05.1997. 15N label was applied to each subplot and herbage was separated into grass and clover, dried and analysed as described previously. Samples were taken every 28 days from these dates, until 28.10.1997, giving a total of 5 sampling dates for each plot. The final 15N applications were performed on 29.09.1997 and 22.9.1997 for the 1-year-old and 2-year-old leys, respectively (Table 4.3.1). Three new control plots for the 1-year-old leys and three new control plots for the 2-year-old leys were chosen in locations that were naturally free of clover. The 2-year-old leys were being cut twice for silage during the course of the experiment. When this happened, the cages were removed from the 2-year-old leys, and replaced with marker canes. During silage cutting, the subplots were covered with plastic sheeting, firmly staked down at the corners. This ensured that the grass and clover in the subplots was not damaged, and also prevented grass clippings from elsewhere in the ley, or manure (spread immediately after silage cutting) from contaminating the subplots.
1999
In 1998, 1,2 and 3-year-old leys were studied as well as 1-year-old leys at Aldroughty. As before, three new subplots were selected within each plot. Three control subplots were chosen for the 1-year-old leys, 2-year-old leys, and 3-year-old leys at Tulloch, and the 1-year-old leys at Aldroughty, making 12 clover-free control subplots in all. The controls were placed in nearby grass-clover leys of the appropriate age. Unlike previous years, the controls were not placed in clover-free zones. Instead, the clover present in the subplots at the start of the experiment was removed by hand. Labelling and analysis followed the same procedure as the previous year, with five applications of label and six sampling dates. Sampling dates are shown in Table 4.3.1 and Table 4.3.2. During the silage cutting, subplots in cut leys were covered with plastic as in the previous year.
2000
In 2000, 1-, 2-, 3- and 4-year-old leys were sampled at Tulloch and 1-year-old leys at Aldroughty. At Tulloch, three new control subplots were chosen within each of plots 16, 18, and 13 for 1, 3 and 4-year old leys, respectively and close to plot 17 for 2-year-old leys. The clover in these plots was removed by hand and lawn edging was used to prevent clover from re-invading. The control plots for Aldroughty were created in a nearby ley. Labelling, sampling and analysis followed the same procedures as the previous two years. As before, subplots in cut leys were covered with plastic during silage cutting. Sampling dates are shown in Table 4.3.1 and Table 4.3.2.
|
Age of ley |
Sample |
Sampling dates |
|||
|
1997 |
1998 |
1999 |
2000 |
||
|
1 |
Initial |
7th July |
9th June |
31st May |
22nd May |
|
1 |
- |
7th July |
28th June |
19th June |
|
|
2 |
29th July |
4th Aug |
26th July |
17th July |
|
|
3 |
26th Aug |
1st Sept |
23rd Aug |
14th Aug |
|
|
4 |
23rd Sept |
29th Sept |
20th Sept |
11th Sept |
|
|
5 |
21st Oct |
27th Oct |
18th Oct |
10th Oct |
|
|
2 |
Initial |
2nd June |
3rd June |
25th May |
|
|
1 |
30th June |
1st July |
22nd June |
||
|
2 |
28th July |
29th July |
20th July |
||
|
3 |
25th Aug |
26th Aug |
17th Aug |
||
|
4 |
22nd Sept |
23rd Sept |
14th Sept |
||
|
5 |
20th Oct |
21st Oct |
12th Oct |
||
|
3 |
Initial |
7th June |
29th May |
||
|
1 |
5th July |
26th June |
|||
|
2 |
2nd Aug |
24th July |
|||
|
3 |
30th Aug |
21st Aug |
|||
|
4 |
28th Sept |
18th Sept |
|||
|
5 |
25th Oct |
16th Oct |
|||
|
4 |
Initial |
1st June |
|||
|
1 |
29th June |
||||
|
2 |
27th July |
||||
|
3 |
24th Aug |
||||
|
4 |
21st Sept |
||||
|
5 |
19th Oct |
||||
Table .1 Sampling dates for 15N dilution study of leys of different ages at Tulloch 1997-2000
|
Age of ley |
sample |
Sampling dates |
|
|
1999 |
2000 |
||
|
1 |
initial |
June 11th |
May 29th |
|
1 |
July 6th |
June 27th |
|
|
2 |
Aug 5th |
July 24th |
|
|
3 |
Sept 2nd |
Aug 21st |
|
|
4 |
Sept 30th |
Sept 15th |
|
|
5 |
Oct 28th |
Oct 12th |
|
Table .2 Sampling dates for 15N dilution study of leys of different ages at Tulloch 1997-2000
In 2001, numbers of white clover flower-heads were counted in the herbage samples that had been collected for the 15N dilution study. The reason for this was that unexplained variations in clover N concentration had been observed in previous years (See section 4.5.3), and it was hypothesised that these may have been due to differences in the proportions of clover dry matter allocated to leaves, petioles, stolons and flowers. Flower buds were counted if they had emerged from the stipule, as were ripe seed heads. Flower stalks were not counted if they had lost the seed head, and detached seed heads were also not counted. Flowerheads were counted to try to explain variations in the N concentration of clover herbage that had been observed previously, and also to test hypotheses about changes in morphology of clover plants in response to soil N.
Flowering rate of white clover was calculated by the formula:
Flowering rate of clover = number of flowerheads per sample/dry matter of clover in sample. [.1]
Flowering rate was analysed by ANOVA on GENSTAT version 6. Model terms were: age of ley (1-, 2-, 3- or 4 years old), date (calculated as number of days after May 1st) and rotation (50% ley or 66% ley).
In August 2001, soil samples collected from clover microsites for chemical analysis (see section on soil chemistry for sampling methods), were examined for clover tap-roots. The number of tap-roots in the bulk sample of soil from clover microsites was recorded for each plot. This gave a figure for the number of tap-roots in 8 soil cores from clover microsites.
pNdfa
Proportion of N derived from the atmosphere, including transfer (pNdfaIT) was calculated from:
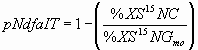 [.1]
[.1]
where %XS15NC is the 15N enrichment of the clover sample, calculated as the difference between the percentage 15N content of the sample and the percentage 15N content of the initial clover sample (background level) from that subplot. Likewise, %XS15NGmo is the 15N enrichment of the grass monoculture, calculated as the difference between the percentage 15N content of the grass monoculture sample and the percentage 15N content of the initial sample from the grass monoculture, averaged for all the monoculture plots of a particular age.
Proportion of N derived from the atmosphere, excluding transfer (pNdfaET), was calculated from:
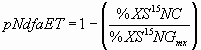 [.2]
[.2]
Where %XS15NGmx is the 15N enrichment of the grass in the grass-clover mixture, calculated as the difference between the 15N content of the grass in the mixed sample and the 15N content of the grass in the initial sample from that subplot.
N fixation including transfer (NfixIT) was calculated from:
![]() [.3]
[.3]
Where DMC is the dry matter yield of clover (kg ha-1) and %NC is the percentage N (by weight) in clover dry matter.
N fixation excluding transfer (NfixET) was calculated from:
![]() [.4]
[.4]
To account for fixed nitrogen present in roots and stolons, Jorgensen and Ledgard, (1997) suggest multiplying NfixET by 1.7. Data for NfixET has been presented without this multiplication factor, except where stated.
N fixation was also calculated by the N difference method:
![]() [.5]
[.5]
Where %NGmx and %NGmo are the %N in the grass mixture and the grass monoculture, respectively and DMGmx and DMGmo are the dry matter yields of the grass mixture and the grass monoculture, respectively.
N transfered from clover to grass was calculated by two isotope methods:
a)![]() [.6]
[.6]
and
b)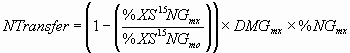 [.7]
[.7]
(Farnham and George, 1994)
This method takes account of the possibility that N transfer from clover to grass might increase uptake of soil N by grass, and increase the yield and nitrogen concentration of the grass.
N transfer was also calculated by a comparison of nitrogen yields in grass mixtures and monocultures (referred to as the N difference method):
![]() [.8]
[.8]
The calculation treats non-fixed N in clover as if it were part of the grass fraction (Ledgard, 1991).
Soil derived N in herbage/kg ha-1 = (N yield of grass/kg ha-1 + N yield of clover/kg ha-1 – NfixET/kg ha-1) [.9] (modified from Danso et al. 1988).
Soil derived N in herbage (SDN) can be used as an estimate of plant available soil N. This assumes that clover, grass and weeds have the same capacity to utilise soil N. A similar approach was used by Doyle et al. (1986) to estimate available soil N. SDN has two components: Grass N yield and soil derived N in clover. The relative proportions of these two fractions give an indication of the ability of clover to compete with grass for soil N.
Data were analysed by Residual maximum likelihood (REML) and ANOVA on Genstat version 6. Model terms were: age of ley (1-, 2-, 3- or 4 years old), sampling period (1, 2, 3, 4 or 5), year (1997, 1998, 1999 or 2000), and rotation (50% ley or 66% ley). For some of the analysis it was necessary to combine age of ley and year into a single variable (1997(1), 1998(1), 1998(2), 1999(1), 1999(2), 1999(3), 2000(1), 2000(2), 2000(3)), because of the unbalanced nature of the data. 4-year-old leys were examined separately in an analysis of the 66% ley rotation in 2000, because the 50% ley rotation did not have a fourth year of ley. Response variates were: Proportion of N derived from the atmosphere, excluding N transfer (pNdfaET), N fixation excluding N transfer (NfixET), %N concentration in clover dry matter, %N concentration in grass dry matter, dry matter of clover, dry matter of grass and soil derived (non-fixed) N in grass and clover. Because of non-normal residuals in the REML analysis, and a few values slightly higher than 1 (probably due to error in the measurements), pNdfaET was transformed by the following formula:
![]() [.10]
[.10]
N fixation values also had non-normal residuals and were transformed by the formula:
![]() [.11]
[.11]
and clover dry matter was also transformed to:
![]() [.12]
[.12]
Dry matter of grass and soil derived N were both transformed by logging. %NC and %NG were not transformed.
It was not possible in this experiment to examine the effects of cutting or grazing on N fixation, because the grass and clover in the subplots was all being cut at monthly intervals, regardless of the management of the surrounding leys. However, grazing generally began earlier in the spring than the 15N dilution experiments, so early spring grazing may have affected the subplots in grazed plots. The level of statistical significance in the REML analysis was set at p=0.05.
Nitrogen fixation was largely a function of clover dry matter, although this relationship varied slightly between years and ages of ley, owing to variations in the pNdfa and N concentration of the clover (sections 4.5.2 and 4.5.3). The lowest N fixation per unit dry matter was observed in 1999 (Figure 4.5.1, Figure 4.5.2, Figure 4.5.3 and Figure 4.5.4).
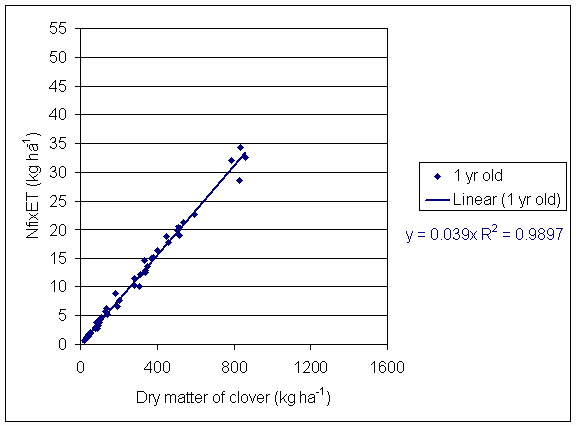
Figure .1 Nitrogen fixation plotted against clover dry matter yield in 1-year-old leys at Tulloch in 1997. Each data point represents data from one subplot.
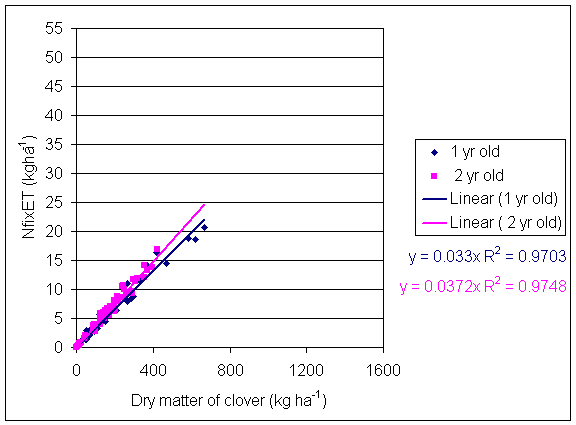
Figure .2 Nitrogen fixation plotted against clover dry matter yield in 1- and 2-year-old leys at Tulloch in 1998. Each data point represents data from one subplot.
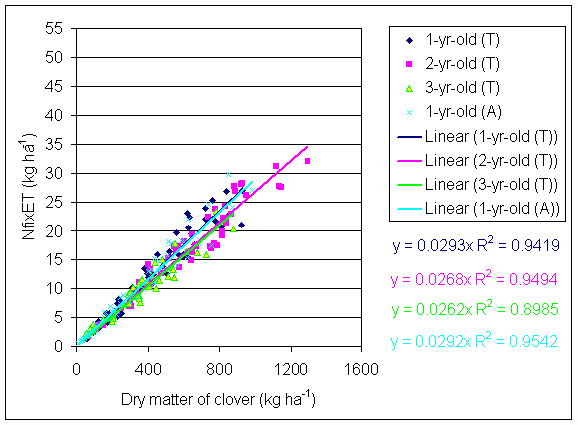
Figure .3 Nitrogen fixation plotted against clover dry matter yield in 1-, 2- and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 1999. Each data point represents data from one subplot.
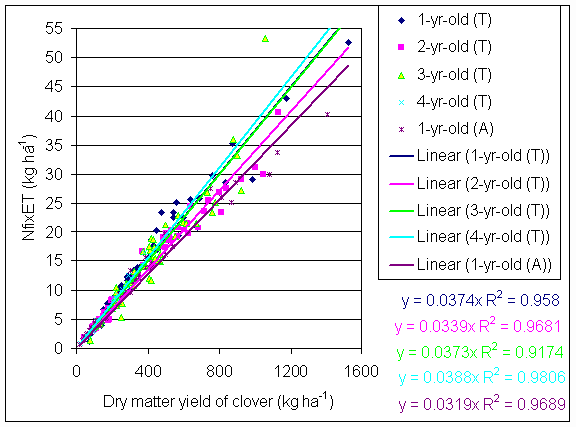
Figure .4 Nitrogen fixation plotted against clover dry matter yield in 1-, 2-, 3- and 4-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 2000. Each data point represents data from one subplot.
Data from all years and ages of ley was included in a single REML analysis. N fixation varied significantly between different years and/or ages of ley (REML p<0.001). N fixation followed a general seasonal trend, peaking roughly in July and August, and declining in September and October (REML p<0.001), and this pattern varied between different years and/or ages of ley (REML p<0.001). Overall, there was no effect of rotation on N fixation, but in some years and/or ages of ley the two rotations had significantly different N fixation values (REML p<0.001). In order to understand this variation better, the analysis was repeated on each year and age of ley individually.
In 1997 NfixET was highest on the first sampling date (7th-29th July) and subsequently declined (Figure 4.5.5, REML p<0.001). In 1997 1-year-old leys, NfixET was similar in both rotations, and there were no differences between the seasonal patterns of N fixation in the two rotations.
Total N fixation was 41.84 kg N ha-1. Taking into account fixed N present in roots, stolons and stubble, the total N fixed between July and October can be estimated at 71.55 kg N ha-1.
N fixation was not measured in June of 1997, but it is possible to estimate the value for June based on data for other years and ages of ley. There is also likely to have been some N fixation in May, although this was not measured directly in any year, because the 15N label was not applied until the end of June.
In other years and ages of ley the N fixation in June was 22–23% of the total N fixation for the sampling period. Assuming a similar seasonal pattern in 1997, the N fixation in June 1997 can be estimated at 16.1 kg N ha-1.
The N yield of clover herbage harvested from ungrazed leys prior to labelling (the initial sample) in 1998 2-year-old leys was 8.76 kg ha-1 (27% of the total for the growing season). In 1999 it was 26.12 kg ha-1 for 2-year-old leys (33% of the total) and 22.79 kg ha-1 for 3-year-old silage plots (23% of the total). In 2000 it was 30.19 kg ha-1 for 2-year-old leys (39% of the total), 23.78 kg ha-1 for 3-year-old plots (29% of the total) and 21.49 kg ha-1 for 4-year-old leys (58% of the total).
Assuming that pNdfa of clover was similar in May and June, and that root:shoot ratios were also similar in both months, the total N fixation figures should be multiplied by a factor of c.1.3 to provide a more realistic estimate of annual N fixation, including N fixed outside the sampling period. This means that, in 1997, N fixation in 1-year-old leys may have been as high as 114 kg N ha-1. N transfer also needs to be taken into account (see section 4.5.11).
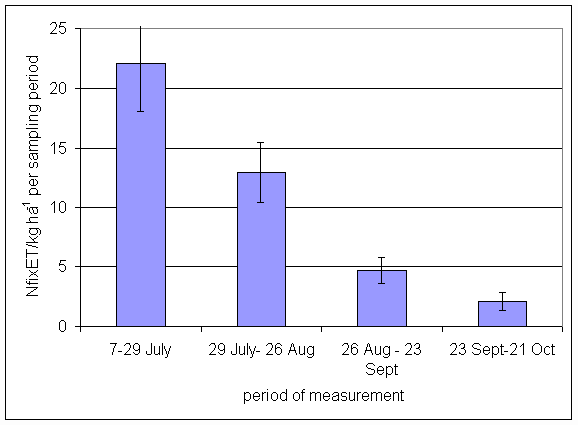
Figure .5 Nitrogen fixation excluding N transfer at Tulloch in 1-year-old leys in 1997. Values represent the accumulated N fixed during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1998 NfixET showed a clear seasonal pattern, declining towards the end of the growing season (REML p<0.001). The seasonal pattern varied with age of ley, showing a clear peak in yield between late July and early September in the 2-year-old leys but not in the 1-year-old leys (REML p<0.001, Figure 4.5.6). N fixation in 1998 was significantly lower than in other years. Total NfixET above ground during the period of sampling was 26.2 and 30.31 kg N ha-1 for 1- and 2-year-old leys, respectively. When N fixation below ground and early in the growing season is taken into account, the estimates became 57.9 and 66.96 kg N ha-1 yr-1 for 1- and 2-year-old leys, respectively. N fixation was not significantly different in the two ages of ley. The two rotations also had similar N fixation, and there were no indirect effects of the rotations on N fixation.
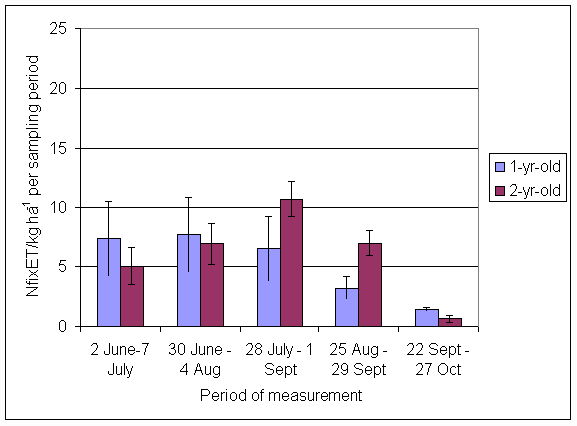
Figure .6 Nitrogen fixation excluding N transfer at Tulloch in 1998, 1- and 2-year-old leys. Values represent the accumulated N fixed during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1999, N fixation peaked between July and September (REML p<0.001), but the seasonal pattern was different in different ages of ley, with 3-year-old leys peaking in July and 1-year-old leys fixing most N in late August/early September (REML p<0.001, Figure 4.5.7). Overall, N fixation was highest in 2-year-old leys (REML p<0.001). N fixation over the sampling period was 59.18, 75.64 and 47.63 kg N ha-1 in 1-, 2- and 3-year-old leys, respectively. Estimated total annual N fixation, above and below ground, was 131.56, 168.15 and 105.88 kg N ha-1 yr-1 for 1-, 2- and 3-year-old leys, respectively. N fixation in 1999 was not affected significantly by the type of rotation.
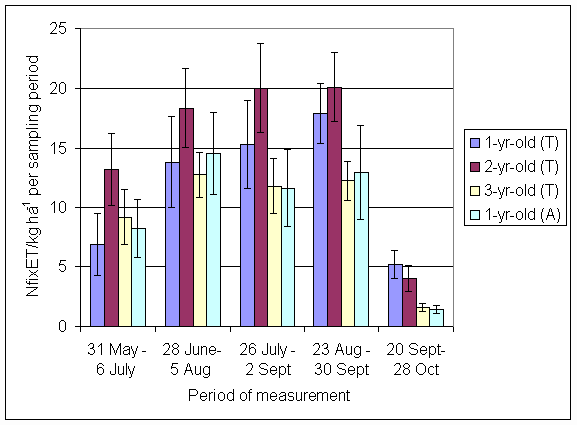
Figure .7 Nitrogen fixation excluding N transfer in 1999, 1-, 2- and 3-year-old leys (Tulloch) and 1-year-old leys (Aldroughty). Values represent the accumulated N fixed during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 2000 N fixation once again showed a strong seasonal trend, declining sharply in autumn (REML p<0.001, Figure 4.5.8). When 1–3-year-old leys from both rotations were considered, the rotation had no significant effect; however, 1 and 3-year-old leys in the 66% ley rotation had significantly higher N fixation than 3-year-old leys in the 50% ley rotation (REML p<0.001, Figure 4.5.9). N fixation was similar in 1,2 and 3-year-old leys (73.7, 70.4, 69.3 kg N ha-1 respectively) but significantly lower in 4-year-old leys (33.5 kg N ha-1) REML p<0.001, Figure 4.5.8). Allowing for N fixed below stubble height and early in the growing season, these figures become 163.8, 156.5, 154.1 and 74.5 kg N ha-1 for 1-2-3- and 4-year-old leys, respectively
When 1–4-year-old leys in the 66% ley rotation only were compared, 3 year-old leys in the 66% ley rotation were seen to have a significantly different seasonal pattern from 1,2 and 4-year-old leys in the 66% ley rotation. N fixation in 3-year-old leys in the 66% ley rotation was highest in the first sample (late May–June), whereas the 1,2 and 4-year-old leys in this rotation showed a peak of N fixation in the 2nd-4th samples (late June-Mid September, REML p<0.001, Figure 4.5.9).
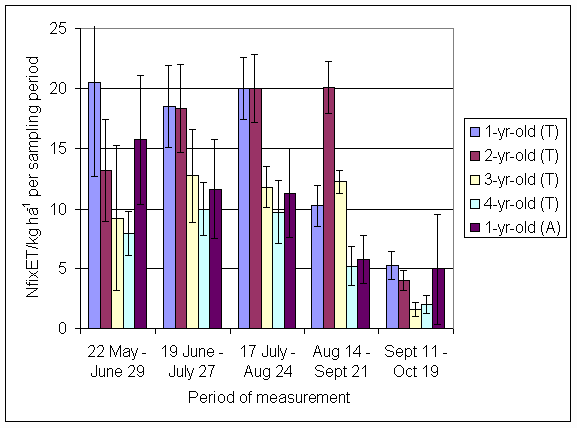
Figure .8 Nitrogen fixation excluding N transfer in 2000, 1-, 2-, 3- and 4-year-old leys (Tulloch) and 1-year-old leys (Aldroughty). Values represent the accumulated N fixed during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In order to examine differences in N fixation between years, 1-year-old leys in 1997, 1998, 1999 and 2000 were compared. N fixation varied significantly with year, lowest in 1998 and highest in 2000 (REML p<0.001). In all years, N fixation declined in autumn (REML p<0.001, Figure 4.5.5-Figure 4.5.8) in 1-year-old leys, but the seasonal pattern varied significantly from year to year (REML p<0.001). In 1999, N fixation was relatively low early in the growing season, but in 2000, N fixation was highest at the start of the season in 1-year-old leys. There were no effects of rotation on 1-year-old leys overall, but there was a significant year ´ month ´ rotation interaction (REML p<0.005) because N fixation was higher in the 66% ley rotation in early and mid-growing season in 1998, 1999 and 2000.
The same procedure was repeated for 2-year-old leys in 1998, 1999 and 2000; and 3-year-old leys in 1999 and 2000. In 2-year-old leys, there were significant differences between years (REML p<0.001) probably due to the low N fixation in 1998, and also significant seasonal variation similar to that seen in the 1-year-old leys (REML p<0.001). The seasonal patterns also varied between years (REML p<0.001), in a similar manner to the 1-year-old leys. There was no effect of rotation on 2-year-old leys when all years were considered.
N fixation was not significantly different in 3-year-old leys in 1999 and in 2000. The seasonal trends in N fixation were significantly different in 1999 and 2000 (REML p<0.001, Figure 4.5.7, Figure 4.5.8). There was also a significant effect of rotation on the seasonal trends. The 3-year-old leys in the 66% ley rotation had higher N fixation than the 3-year-old leys in the 50% ley rotation (REML p<0.001, Figure 4.5.9). This effect was more marked in 2000 than 1999 (REML p<0.001).
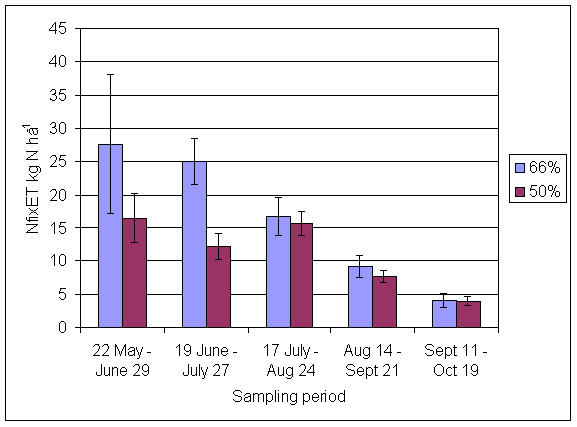
Figure .9 N fixation excluding transfer in 3-year-old leys in the 66% ley rotation and the 50% ley rotation in 2000. Values represent the accumulated N fixed during the month of sampling (mean of 6 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
NfixET at Aldroughty was closely related to the dry matter yield of clover (Figure 4.5.3, Figure 4.5.4). In both years the rate of N fixation per unit of clover dry matter was low, because of low %NC and pNdfa, although in 1999 these were low at Tulloch also. Total N fixation in 1-year-old leys at Aldroughty over the sampling period was 48.8 kg N ha-1 in 1999 and 49.34 kg N ha-1 in 2000. N fixation was not significantly different in the two years. The estimated total annual N fixation, including N fixation below cutting height and outside the growing season (see above) was 107.148 kg N ha-1 yr-1 in 1999 and 109.04 kg N ha-1 yr-1 in 2000.
There were no significant differences between N fixation in 1-year-old leys at Aldroughty and Tulloch (Figure 4.5.7, Figure 4.5.8), and the seasonal patterns were similar (Figure 4.5.7, Figure 4.5.8). There was no significant effect of crop rotation on N fixation at Aldroughty.
When data from all years and ages were compared, pNdfa varied with year and/or age (REML p<0.001). pNdfa also showed a seasonal pattern (REML p<0.001), which varied with year and/or age (REML p<0.001).There were also significant differences between the two rotations in some years and/or ages of ley (REML p<0.005). To understand these patterns better, data from each year and age of ley were examined separately:
In 1997 pNdfaET showed a distinct seasonal pattern, declining in autumn (Figure 4.5.10, REML p<0.001). There was no overall effect of rotation on pNdfaET, but in August and September pNdfa in the 66% ley rotation was lower than pNdfa in the 50% ley rotation (REML p<0.01). Over the whole sampling period, pNdfa was on average 0.79 (calculated as total N fixation/total clover N yield). This is likely to be a low estimate because sampling started late in 1997.
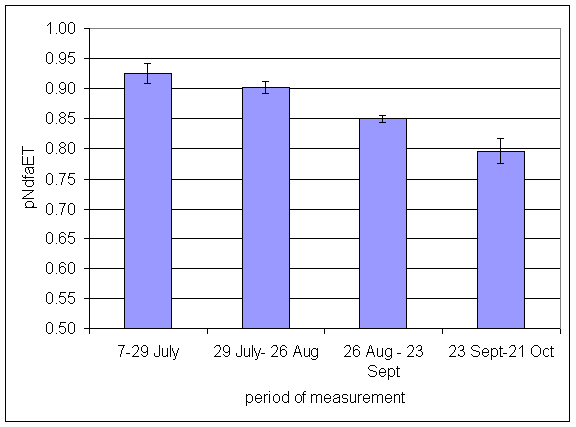
Figure .10 Proportion of nitrogen in clover herbage derived from atmosphere, excluding transfer (pNdfaET) in 1-year-old leys at Tulloch in 1997. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1998 pNdfa followed a similar seasonal pattern to 1997, declining significantly in September and October (REML p<0.001, Figure 4.5.11). 1- and 2-year-old leys did not have significantly different values of pNdfa and there were no significant differences between the two rotations. Over the whole growing season, average pNdfa was 0.94 in 1-year-old leys and 0.95 in 2-year-old leys.
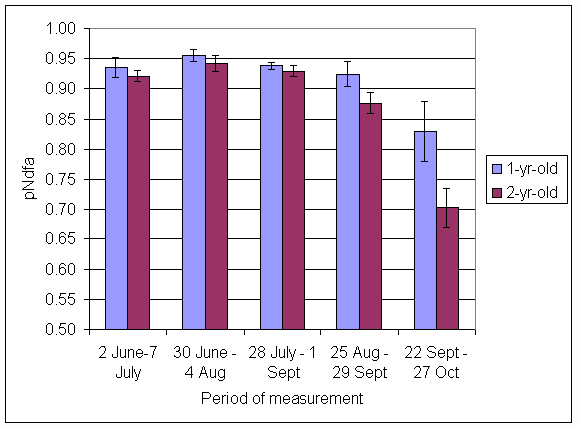
Figure .11 Proportion of nitrogen in clover herbage derived from atmosphere, excluding transfer (pNdfaET) in 1- and 2-year-old leys at Tulloch in 1998. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1999 pNdfaET followed a similar seasonal trend to previous years, declining in October (REML p<0.001, Figure 4.5.12). pNdfaET was highest in 1-year-old leys and lowest in 3-year-old leys (REML p<0.001). pNdfaET was higher in the 50% ley rotation, in the 1 and 3-year-old leys (REML p<0.01). Over the whole growing season, pNdfaET was 0.95, 0.95 and 0.92 in 1-, 2- and 3-year-old leys, respectively.
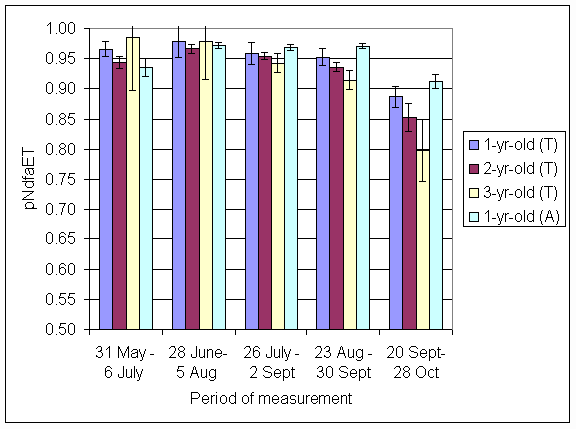
Figure .12 Proportion of nitrogen in clover herbage derived from atmosphere, excluding transfer (pNdfaET) in 1,2 and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 1999. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 2000, pNdfaET followed a similar seasonal trend to previous years, declining significantly in autumn (REML p<0.001, Figure 4.5.13). When pNdfaET was compared in 1-, 2-, 3- and 4-year-old leys in the 66% ley rotation were compared, there was no significant difference between leys of different ages. Over the whole growing season, pNdfa was on average 0.91, 0.92, 0.85 and 0.9 in 1-, 2-, 3- and 4-year-old leys, respectively.
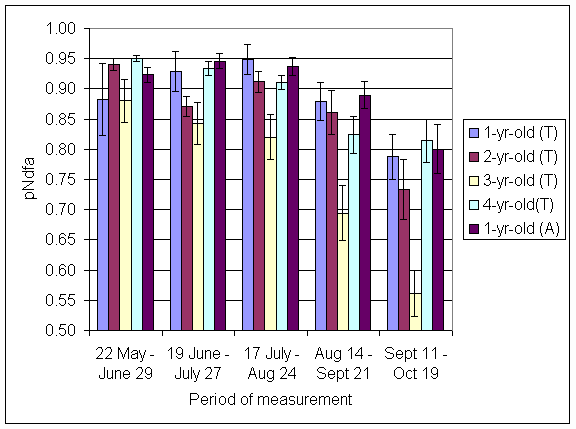
Figure .13 Proportion of nitrogen in clover herbage derived from atmosphere, excluding transfer (pNdfaET) in 1-, 2-, 3- and 4-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 2000. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
pNdfaET values in 1-year-old leys were compared in all years: 1997 and 2000 had significantly lower pNdfaET than other ages of ley (REML p<0.001, Figure 4.5.10, Figure 4.5.11, Figure 4.5.12 and Figure 4.5.13). There was significant seasonal variation (REML p<0.001), and this varied from year to year (REML p<0.001). In 1999, pNdfaET remained relatively high into October (Figure 4.5.12), unlike other years. There was also a significant year ´ month ´ rotation interaction (REML p=0.001), because of low pNdfa in the 50% ley rotation in September 1997.
When 2-year-old leys in 1998, 1999 and 2000 were compared, there was significant seasonal variation (REML p<0.001), which varied significantly between years (REML p<0.001), in 2000, pNdfaET declined unexpectedly in late June-early July (Figure 4.5.13). There was also a significant interaction between year, month and rotation (REML p<0.005), caused by slightly lower pNdfa values in the 66% ley rotation, in 1-year-old leys late in 2000, when compared to the 50% ley rotation.
pNdfaET in 3-year-old leys was lower in 2000 than in 1999 (REML p<0.001). The seasonal pattern also differed between the two years: in 2000 pNdfa declined steadily through the growing season, whereas in 1999 it remained high until October (REML p<0.001, Figure 4.5.12 and Figure 4.5.13). There was a slight difference between the seasonal patterns in the two rotations in 2000, pNdfa in the 66% ley rotation was lower than pNdfa in the 50% ley rotation late in the year (REML p<0.005).
Aldroughty
pNdfaET at Aldroughty was not significantly different from pNdfa at Tulloch in the years and ages of leys studied (Figure 4.5.12. and Figure 4.5.13). The seasonal patterns of pNdfa at both sites were also similar. pNdfaET was significantly higher in 1999 (average 0.96) than in 2000 (average 0.91, ANOVA p<0.01).
When data from all years and ages of ley was considered, %NC varied significantly with year and/or age (REML p<0.001). %NC followed a seasonal pattern (REML p<0.001), which varied with year and/or age of ley (REML p<0.001). There were also significant differences between the two rotations in some year or ages of ley (REML p=0.01). Data from each year and age of ley were re-analysed separately:
In 1997, percentage nitrogen in clover (%NC) varied significantly over the growing season, peaking in September and at a minimum in July (REML p<0.001). There was no effect of rotation on %NC in 1997. %NC was on average 4.35% (calculated as: total clover N yield / total clover dry matter). This was likely to be an overestimate of the average annual nitrogen concentration, because sampling began late in 1997.
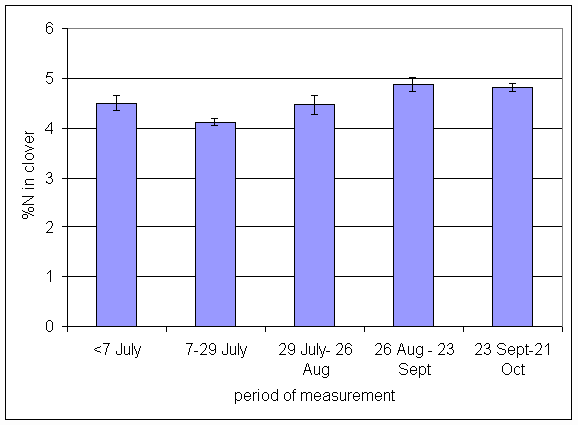
Figure .14 Concentration of nitrogen in clover herbage expressed as a percentage of dry matter. 1-year-old leys at Tulloch, in 1997. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1998, %NC varied with season in a similar way to the previous year (REML p<0.001, Figure 4.5.15). There was no significant difference between 1- and 2-year-old leys overall, but 2-year-old leys had significantly higher %NC in June than 1-year-old leys (REML p<0.001, Figure 4.5.15). The average %NC, in 1998 at Tulloch was 3.7 in 1-year-old leys and 3.85 in 2-year-old leys.
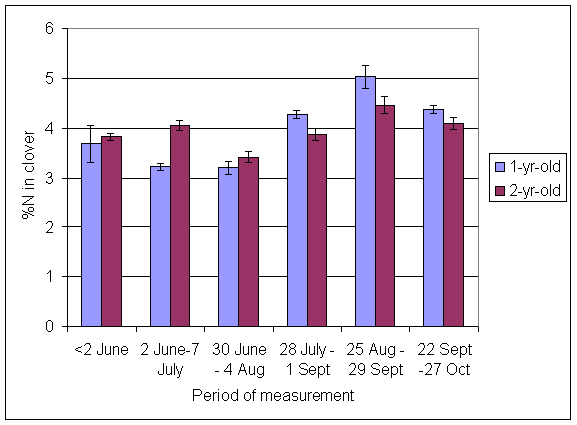
Figure .15 Concentration of nitrogen in clover herbage expressed as a percentage of dry matter. 1- and 2-year-old leys at Tulloch, in 1998. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1999, %NC was significantly lower in 2-year-old leys than other ages of ley (REML p<0.001, Figure 4.5.16). There was significant seasonal variation (REML p<0.001) similar to that seen in 1997. The seasonal pattern in 3-year-old leys was slightly different from that of the other ages of ley, because %NC was relatively low at the start of the growing season and relatively high at the end of the growing season in 3-year-old leys (REML p<0.001, Figure 4.5.16). There was no effect of rotation on %NC in 1999. The average %NC values in 1999 at Tulloch were 3.1, 2.87 and 3.87 in 1-, 2- and 3-year-old leys, respectively.
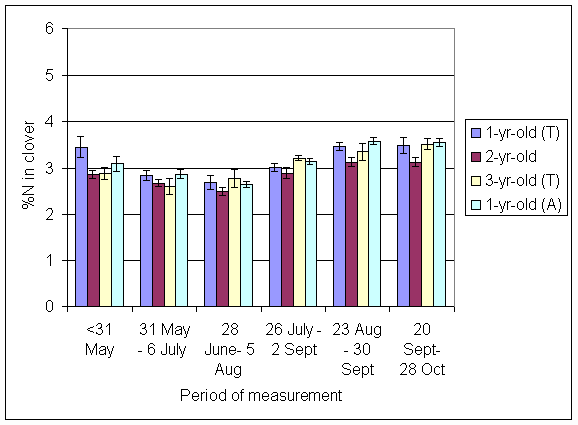
Figure .16 Concentration of nitrogen in clover herbage expressed as a percentage of dry matter. 1-, 2- and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty, in 1999. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 2000, %NC was again significantly lower in 2-year-old leys (REML p<0.001, Figure 4.5.17). %NC showed a significant seasonal pattern (REML, p<0.001, Figure 4.5.17), similar to previous years. Three-year-old leys showed the greatest seasonal variation, and 1 and 4-year-old leys varied least over the course of the growing season (REML p<0.001, Figure 4.5.17). %NC was also higher in the 66% ley rotation in 1-year-old leys (REML p<0.001). Average %NC in 2000 at Tulloch was 4.34, 3.79, 4.38 and 4.21 in 1-, 2-, 3- and 4-year-old leys, respectively.
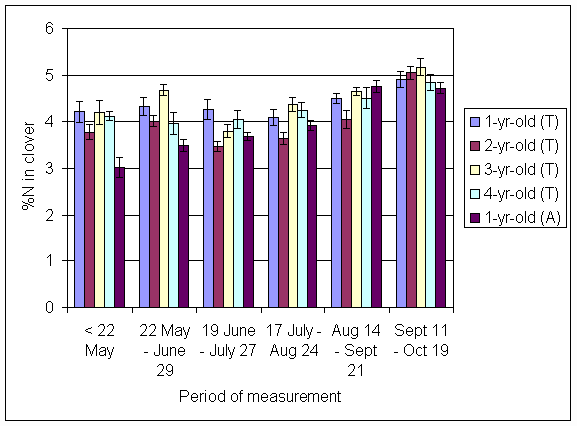
Figure .17 Concentration of nitrogen in clover herbage expressed as a percentage of dry matter. 1-, 2-, 3- and 4-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 2000. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
When only 1-year-old leys were examined, %NC was found to be lowest in 1999 and highest in 1997 and 2000 (REML p<0.001). The seasonal variation was greatest in 1998 (REML p<0.001). There was no significant effect of rotation on 1-year-old leys overall.
In 2-year-old leys, %NC was lower in 1999 (REML p<0.001). The seasonal pattern in 2-year-old leys varied between years, with most seasonal variation in 2000 and least in 1999 (REML p<0.001).
In 3-year-old leys, there was significantly higher %NC in 1999 than in 2000 (REML p<0.001). The seasonal pattern was also significantly different, between the two years: in 1999 %NC was at a minimum in July, whereas in 2000 %NC was high in July and fell to a minimum in August (REML, p<0.001, Figure 4.5.16 and Figure 4.5.17). %NC was higher in the 66% ley rotation than the 50% ley rotation in 3-year-old leys (REML p=0.01), especially early in the growing season (REML p<0.001) and in 1999 (REML p=0.01).
The concentration of N in clover was significantly higher in 2000 than in 1999 at Aldroughty (ANOVA p>0.001, Figure 4.5.16 and Figure 4.5.17). %NC in 1-year-old leys at Aldroughty was not significantly different from %NC in 1-year-old leys at Tulloch. The seasonal patterns of the concentration of N in clover were also similar at Tulloch and Aldroughty in 1999 and 2000 (Figure 4.5.16 and Figure 4.5.17). Average N concentration at Aldroughty was 3.038% in 1999 and 4.15% in 2000
When data from all years and ages of leys was considered, clover yield varied significantly with year and/or age (REML p<0.001) and season (REML p<0.001). The seasonal pattern of growth also varied with year and/or age (REML p<0.001). There was no overall effect of rotation on clover yield, but in some years and/or ages of ley there were differences between the two rotations (REML p<0.001). Data from each year and age of ley were re-analysed individually:
In 1997, the dry matter yield of clover showed a significant seasonal trend (REML p<0.01) which was very similar to the seasonal trend of N fixation (Figure 4.5.5 and Figure 4.5.18). There was no effect of rotation type on clover yield in 1997. Total clover yield in 1997 was 1211.3 kg dry
matter ha-1. This is a low estimate because sampling began one month later in 1997 than in other years.
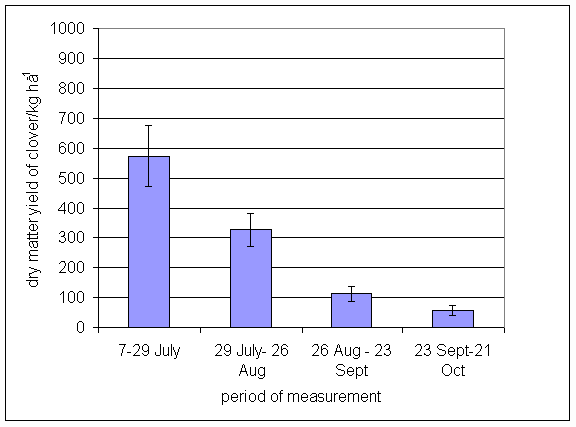
Figure .18 Clover herbage dry matter yield at Tulloch in 1-year-old leys in 1997. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1998, clover yield did not vary significantly between 1- and 2-year-old leys. There were similar seasonal trends to N fixation (Figure 4.5.6 and Figure 4.5.19), and these were significantly different in the two ages of ley (REML p<0.001), because the peak of growth in 2-year-old leys occurred later than the peak of growth in 1-year-old leys (Figure 4.5.19). There was no effect of rotation type on clover yield in 1998. Total yields of clover over the sampling period were 756.1 and 869.8 kg dry matter ha-1 in 1- and 2-year-old leys, respectively. Over the whole year, the yields would have been higher, because of growth of clover before the start of the growing season. In 2-year-old leys, the total annual clover yield was 1120.6 kg dry matter
ha-1. It was not possible to calculate this in the 1-year-old leys because of grazing prior to the start of the experiment.
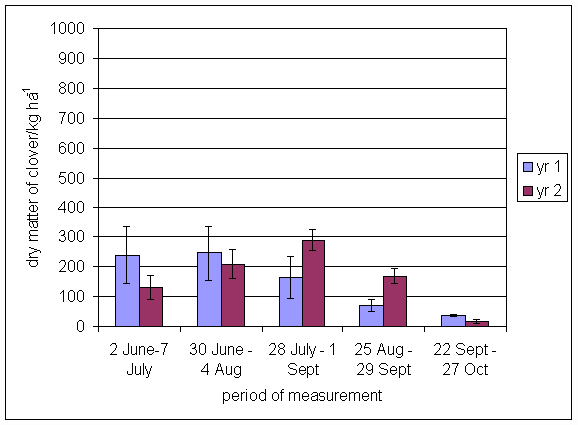
Figure .19 Clover herbage dry matter yield at Tulloch in 1- and 2-year-old leys in 1998. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1999, clover yield was significantly higher in 2-year-old leys than in other ages of ley (REML p<0.001, Figure 4.5.20). There was a clear seasonal trend, with peak clover production in mid – late summer (Figure 4.5.20), closely following the seasonal pattern of N fixation. The seasonal patterns of clover growth were not the same in all ages of ley: yield in 2 and 3-year-old leys declined after midsummer, but yields in 1-year-old leys remained high into August (Figure 4.5.20). Total clover yields for the sampling period were 2020.5, 2809.7 and 1778.4 kg dry matter ha-1 in 1-, 2- and 3-year-old leys, respectively. The total annual clover yield (including the initial sample, in ungrazed plots) was 3681.8 kg dry matter ha-1 and 2491.7 kg dry matter ha-1 for 2-year-old and 3-year-old uncut leys, respectively. There was no significant effect of rotation type on clover yield in 1999.
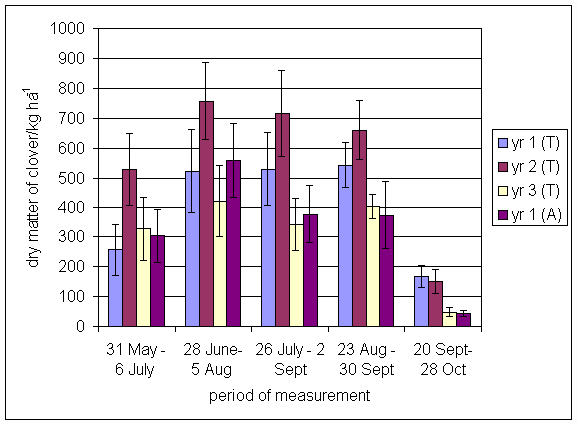
Figure .20 Clover herbage dry matter yield in 1-, 2- and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 1999. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 2000, there was no significant difference between the clover yields in 1,2 and 3-year-old leys, but yields in 4-year-old leys were significantly lower (REML p=0.005). There were strong seasonal trends, similar to those of N fixation (REML p<0.001, Figure 4.5.8 and Figure 4.5.21). Clover yield in 3 and 4-year-old leys started to decline earlier in the year than clover yield in 1- and 2-year-old leys (REML p<0.001, Figure 4.5.21). Clover yields in 1 and 3-year-old leys were higher in the 66% ley rotation than the 50% ley rotation (REML p<0.01). Clover yields for the sampling period were 1909.4, 2032.5, 1878.4 and 985.9 kg dry matter ha-1 for 1-, 2-, 3- and 4-year-old leys respectively. Total annual clover yields (including initial sample, ungrazed plots only) were 2830.8, 2089.1 and 1509.7 kg dry matter ha-1 in 2-year-old, 3-year-old (ungrazed) and 4-year-old leys, respectively.
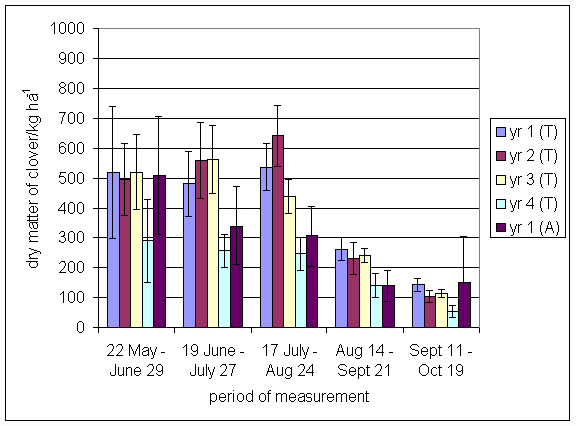
Figure .21 Clover herbage dry matter yield in 1-, 2-, 3- and 4-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 2000. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
When clover yields in 1-year-old leys were compared across all years, yields were lowest in 1998 and highest in 1999 (REML p<0.001). Seasonal variation of clover yield varied from year to year in a manner similar to N fixation (REML p<0.001, Figure 4.5.18, Figure 4.5.19, Figure 4.5.20 and Figure 4.5.21,).
In 2-year-old leys, clover yield was highest in 1999 and lowest in 1998 (REML p<0.001). Clover yield followed similar seasonal patterns to N fixation (Section 4.5.1). There was no effect of rotation on 2-year-old leys. Clover yield was not significantly different in 1999 and 2000 in 3-year-old leys. The seasonal patterns were generally similar to the seasonal patterns of N fixation. 3-year-old leys in the 66% ley rotation had higher clover yields than 3-year-old leys in the 50% ley rotation.
Data from Tulloch on ground cover of clover in Autumn, showed broadly similar patterns of age related variation to the patterns of clover yield observed in this study (Figure 4.5.22), 2-year-old leys generally had the highest clover ground cover. There was also considerable variation in clover ground cover between years.
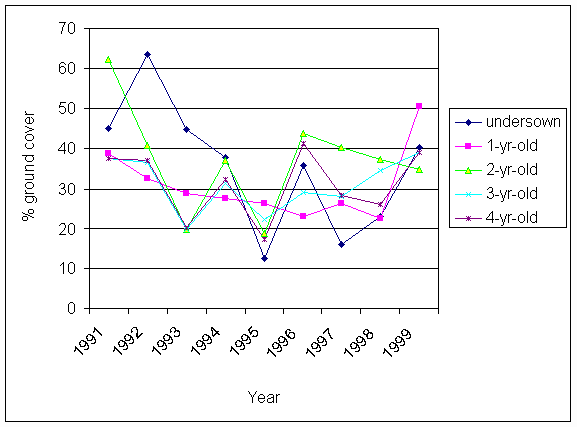
Figure .22 Percentage ground cover of clover in autumn in successive years since the establishment of the trial. Each point is a mean of four samples (SAC data)
Clover yields in 1-year-old leys at Aldroughty were not significantly different from clover yields in 1-year-old leys at Tulloch in 1999 or 2000. The clover yields in 1999 at Aldroughty were not significantly different from the clover yields in 2000 (Figure 4.5.20 and Figure 4.5.21). There was no significant effect of rotation on clover yields at Aldroughty. Total clover yields were 2809.1 kg dry matter ha-1 in 1999 and 1445.9 kg dry matter ha-1 in 2000.
Overall, soil derived N (SDN) in grass and clover varied significantly between different years and/or ages of ley (REML p<0.001). There was a significant seasonal pattern (REML p<0.001), which varied between different years and/or ages of ley (REML p<0.001). The two rotations also had different levels of soil derived N overall (REML p<0.01), especially in certain years and/or ages of ley (REML p<0.001). To understand these patterns better, each year and age of ley were examined individually:
In 1997, Soil derived N declined over the course of the growing season (REML p<0.001, Figure 4.5.23). There was no significant effect of rotation on the leys studied in 1997. Soil derived N over the whole growing season in 1997, 1-year-old leys was 109.83 kg N ha-1. The total annual SDN would have been higher than this, because sampling began late in 1997. 95.6% of SDN was accounted for by N in grass (grass N yield).
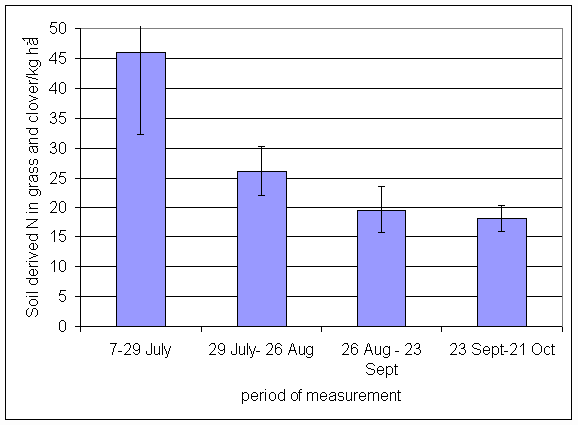
Figure .23 Soil derived N in grass and clover herbage in 1-year-old leys at Tulloch in 1997. Values represent the total accumulation of non-fixed N during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1998, soil derived N was similar in 1- and 2-year-old leys (Figure 4.5.24). There was a general decline in SDN over the course of the growing season (REML p<0.001, Figure 4.5.24), although in August, SDN was high in 2-year-old leys and low in 1-year-old leys (REML p<0.001, Figure 4.5.24). Total SDN was 59.4 kg N ha-1 in 1-year-old leys and 69 kg N ha-1 in 2-year-old leys. These are likely to be low estimates for total annual SDN, as there was considerable grass growth prior to the first sample. The total grass N yield (including the initial sample) in 2-year-old leys was 106.3 kg N ha-1 (1-year-old leys had been grazed prior to the start of sampling, so early season grass yields couldn’t be measured). Grass N yield accounted for 94.1% and 97.6% of SDN in 1- and 2-year-old leys, respectively.
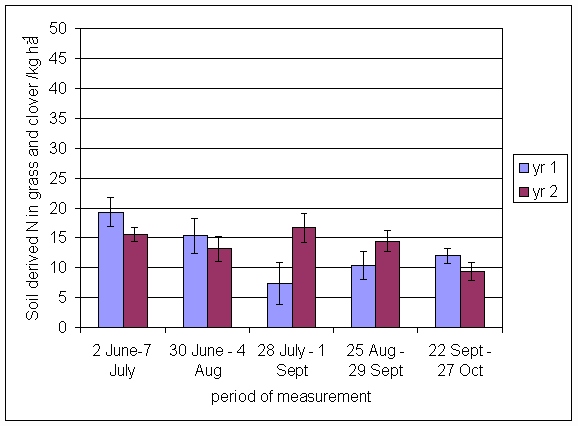
Figure .24 Soil derived N in grass and clover herbage in 1- and 2-year-old leys at Tulloch in 1998. Values represent the total accumulation of non-fixed N during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1999, SDN was highest in 3-year-old leys (REML p<0.01, Figure 4.5.25). SDN was 47.22, 46.3, and 65.19 kg N ha-1 in 1-, 2- and 3-year-old leys respectively. Grass N yield accounted for 93.9% of SDN in 1-year-old leys 90.3% of SDN in 2-year-old leys and 95.1% of SDN in 3-year-old leys. These are low estimates for total annual SDN, as there was considerable grass growth early in the season. Total annual grass N yield (including the initial sample) was 72.7 kg N ha-1 in 2-year-old leys and 89.1 kg N ha-1 in 3-year-old cut leys. SDN was low at the beginning and end of the growing season (REML p<0.001). The largest seasonal variation was seen in the 3-year-old leys (REML p<0.001, Figure 4.5.25). There was no significant effect of rotation on SDN in 1999.
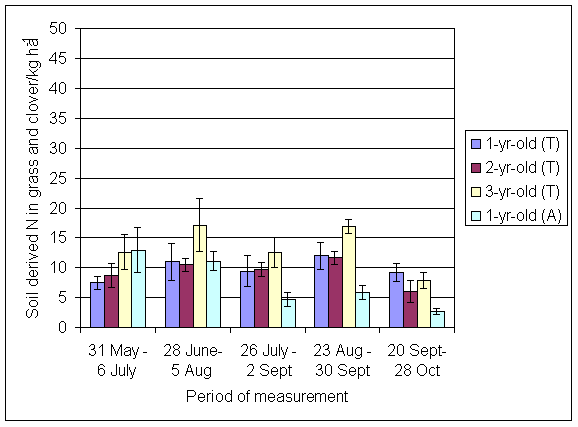
Figure .25 Soil derived N in grass and clover herbage in 1-, 2- and 3-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 1999. Values represent the total accumulation of non-fixed N during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 2000, SDN was lowest in 2-year-old leys and highest in 3-year-old leys (REML p<0.001, Figure 4.5.26). SDN was 111.8, 72.7, 139.0 and 114.0 kg N ha-1 in 1-, 2-, 3- and 4-year-old leys, respectively. Grass N yield accounted for 92.6% of SDN in 1-year-old leys, 91.6% of SDN in 2-year-old leys, 90.8% of SDN in 3-year-old leys and 96.5% of SDN in 4-year-old leys. Total annual SDN was probably considerably higher than the figure given above. Total grass N yield (including initial sample) was 105.2 kg N ha-1 in 2-year-old leys, 165.9 kg N ha-1 in 3-year-old cut leys and 171.8 kg N ha-1 in 4-year-old leys. SDN was highest in late summer (REML p<0.001, Figure 4.5.26), but the seasonal pattern varied greatly between different ages of ley. In 4-year-old leys SDN reached a maximum in late August-early September, approximately one month after younger leys (REML p<0.001, Figure 4.5.26).
SDN was higher in leys in the 66% ley rotation, than the 50% ley rotation (REML p<0.001) especially in 3-year-old leys (REML p=0.01, Figure 4.5.27).
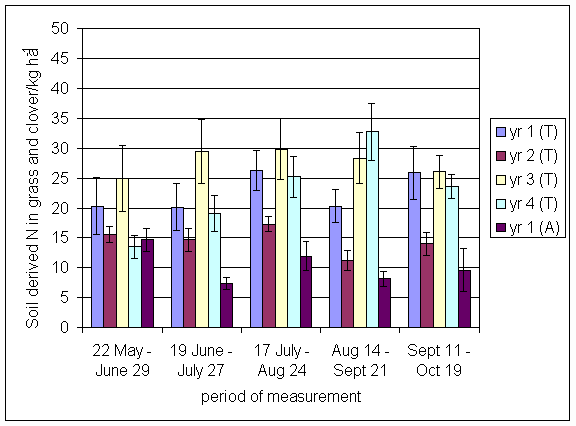
Figure .26 Soil derived N in grass and clover herbage in 1-, 2-, 3- and 4-year-old leys at Tulloch and 1-year-old leys at Aldroughty in 2000. Values represent the total accumulation of non-fixed N during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
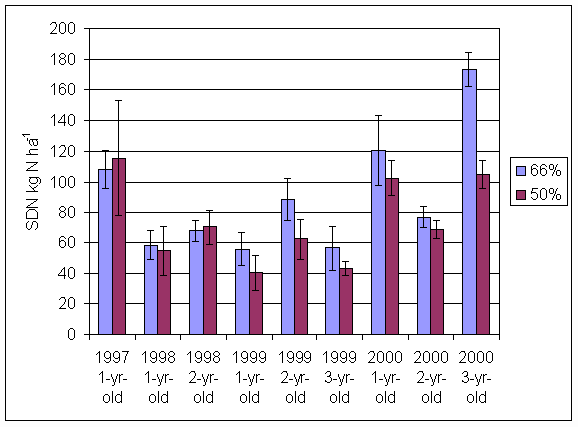
Figure .27 Soil derived N in grass and clover herbage (kg N ha-1) in leys in the 66% ley rotation and the 50% ley rotation. Columns represent total soil derived N over the growing season (mean of 6 subplots). Bars represent standard errors.
In the 1-year-old leys, SDN was highest in 1997 and 2000 and lowest in 1998 and 1999 (REML p<0.001, Figure 4.5.23-Figure 4.5.26). There was no significant seasonal trend when all 1-year-old leys were considered, but there was a clear seasonal trend in 1997 (REML p<0.001, Figure 4.5.23).
There was no effect of rotation on SDN when only 1-year-old leys were considered.
In the 2-year-old leys, SDN was lowest in 1999 (REML p<0.001, Figure 4.5.25-Figure 4.5.27). SDN was low at the beginning and end of the growing season and peaked in late summer in 2-year-old leys (REML p<0.001, Figure 4.5.24-Figure 4.5.26), but in 2000 SDN rose slightly at the end of the year (REML p<0.001).
In the 3-year-old leys, SDN was significantly higher in 2000 than in 1999 (REML p<0.001, Figure 4.5.25 and Figure 4.5.26). SDN was low at the beginning and end of the growing season in both 1999 and 2000 (REML p<0.001) but in 1999, SDN was also low in late summer (REML p<0.001), Figure 4.5.25 and Figure 4.5.26). SDN was higher in the 66% ley rotation (REML p<0.001), especially early in the year (REML p<0.001). The difference was greatest early in the growing season in 2000 (REML p<0.005).
SDN in 1-year-old leys was significantly higher in 2000 than 1999 at Aldroughty (ANOVA p<0.05, Figure 4.5.25 and Figure 4.5.26). There were no significant differences between SDN at Tulloch and Aldroughty in 1999. In 2000, SDN was significantly higher at Tulloch than Aldroughty (ANOVA p<0.001, Figure 4.5.26).
When data from all years and ages of leys was examined, the nitrogen concentration of grass (%NG) varied significantly between years and/or ages of ley (REML p<0.001). There was a significant seasonal component to the variation (REML p<0.001) which varied with year and/or age of ley. %NG was also affected by the type of rotation (REML p<0.01), and this effect was stronger in some years and/or ages of ley than others (REML p<0.001). In order to clarify this, the data for each year and age of ley were examined separately.
In 1997, %NG was significantly higher in late summer and autumn than in early summer (REML p<0.001, Figure 4.5.28). There was no effect of rotation type on %NG in 1997. Average %NG (calculated as total grass Nyield / total grass dry matter yield) was 3.38%
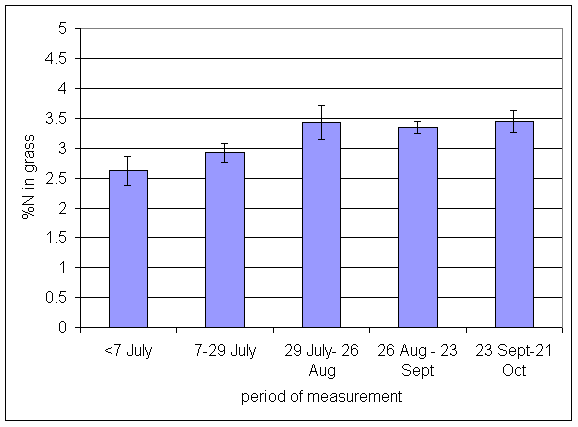
Figure .28 Concentration of nitrogen in grass herbage expressed as a percentage of dry matter. 1-year-old leys at Tulloch, in 1997. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1998, average %NG in 1-year-old leys was 2.39% and in 2-year-old leys, 2.52%. These values were not significantly different. %NG rose over the course of the growing season (REML p<0.001, Figure 4.5.29). In the 2-year-old leys, %NG was relatively high in June, compared to the 1-year-old leys (REML p<0.001). There was no effect of rotation on %NG in 1998.
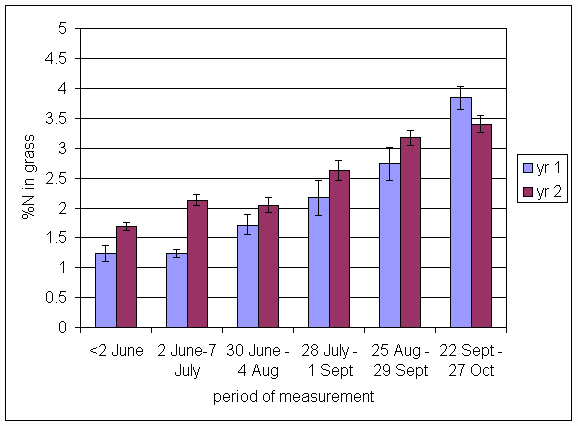
Figure .29 Concentration of nitrogen in grass herbage expressed as a percentage of dry matter. 1- and 2-year-old leys at Tulloch, in 1998. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1999, %NG was significantly higher in 3-year-old leys than in other ages of ley (REML p<0.005, Figure 4.5.30). %NG rose over the course of the growing season (REML p<0.001), and rose most sharply in the 3-year-old leys (REML p<0.01, Figure 4.5.30). %NG was significantly higher in the 66% ley rotation in 1999 (REML p<0.001). Average %NG for the growing season was 1.92, 1,9 and 2.01 in 1-, 2- and 3-year-old leys respectively.
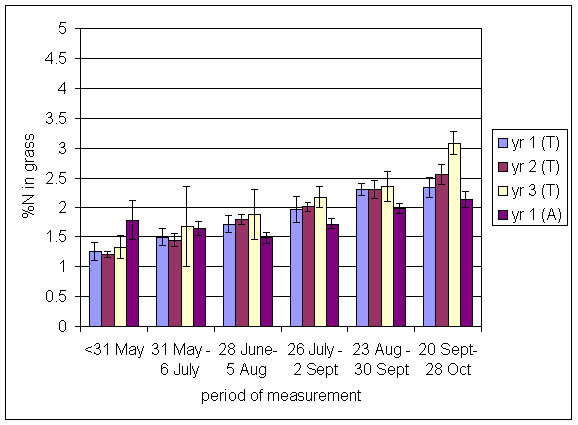
Figure .30 Concentration of nitrogen in grass herbage expressed as a percentage of dry matter. 1-, 2- and 3-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 1999. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 2000, %NG was highest in 3-year-old leys and lowest in 2-year-old leys (REML p<0.001, Figure 4.5.31). %NG rose over the course of the growing season (REML p<0.001, Figure 4.5.31). This seasonal pattern was not the same for all ages of ley: 3-year-old leys had relatively high %NG at the start of the season, but at the end of the year had lower %NG than other ages of ley. 2-year-old leys had low %NG to begin with and high %NG at the end of the year (REML p<0.001, Figure 4.5.31). In 3-year-old leys, %NG was significantly higher in the 66% ley rotation than the 50% ley rotation (REML p<0.01, data in appendix). Average %NG for the whole growing season was 3.04, 2.59, 3.19 and 2.99 for 1-, 2-, 3- and 4-year-old leys, respectively.
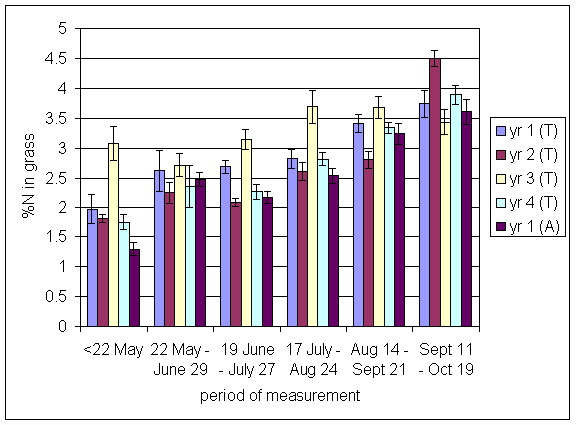
Figure .31 Concentration of nitrogen in grass herbage expressed as a percentage of dry matter. 1-, 2-, 3- and 4-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 2000. Columns represent means of 12 subplots. The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
%NG in 1-year-old leys, was highest in 1997 and 2000 and lowest in 1999 (REML p<0.001, Figure 4.5.28-Figure 4.5.31). %NG rose as the growing season progressed (REML p<0.001) and this increase was most marked in 1998 (REML p<0.001, Figure 4.5.28-Figure 4.5.31). There was no effect of rotation on %NG overall, but %NG was higher in August 1997 in the 66% rotation than the 50% rotation (REML p=0.001).
In 2-year-old leys, %NG was significantly lower in 1999 (REML p<0.001, Figure 4.5.29 -Figure 4.5.31). %NG rose over the course of the growing season in 2-year-old leys (REML p<0.001), and rose most sharply in 2000 (REML p<0.001). There was no effect of rotation on 2-year-old leys.
In 3-year-old leys, %NG was significantly higher in 2000 than 1999 (REML p<0.001, Figure 4.5.30 and Figure 4.5.31). %NG rose significantly over the course of the growing season (REML p<0.001). In 1999, %NG was much lower at the start of the growing season than in 2000 (REML p<0.001). %NG was higher in the 66% ley rotation than the 50% ley rotation in 3-year-old leys (REML p<0.001), especially in 2000 (REML p<0.005).
%NG was significantly higher in 2000 (2.66%) than 1999 (2.17%) at Aldroughty (ANOVA p<0.001, Figure 4.5.30 and Figure 4.5.31). The concentration of N in grass in 1-year-old leys at Aldroughty was higher than at Tulloch in 1999 (ANOVA p<0.05, Figure 4.5.30 and Figure 4.5.31) and lower in 2000 (ANOVA p<0.001, Figure 4.5.30 and Figure 4.5.31).
When the entire data set was analysed, grass yield varied significantly between different years and/or ages of ley (REML p<0.001). Grass yield was low in Autumn (REML p<0.001), but this seasonal pattern varied significantly with year and/or age of ley (REML p<0.001). Grass yield was also affected by the type of rotation in some years and/or ages of ley (REML p=0.001). In order to understand these patterns more clearly, each year and age of ley was examined individually.
In 1997, grass yield declined significantly over the course of the growing season (REML p<0.001, Figure 4.5.32). Total grass yield for the sampling period was 3105.5 kg dry matter ha-1. The total annual grass yield would have been much higher, because sampling began late in 1997. There was no effect of rotation on grass yield in 1997.
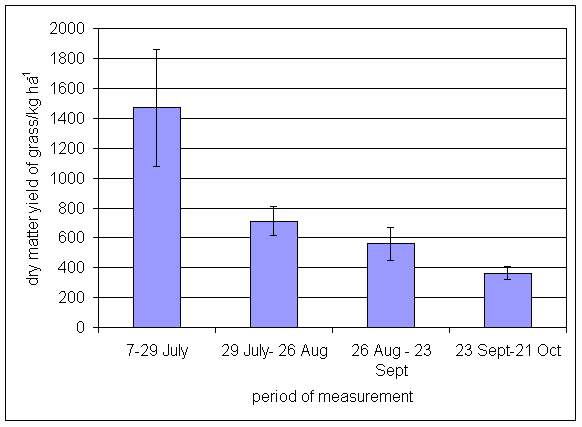
Figure .32 Grass herbage dry matter yield at Tulloch in 1-year-old leys in 1997. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1998, grass yield was 2328.2 and 2658.8 kg dry matter ha-1 for 1- and 2-year old leys, respectively. These values were not significantly different. Grass dry matter yields declined as the growing season progressed (REML p<0.001). Grass yields in late summer were lower in 1-year-old leys than 2-year-old leys (REML p<0.001, Figure 4.5.33). There was no effect of rotation on grass yield in 1998.
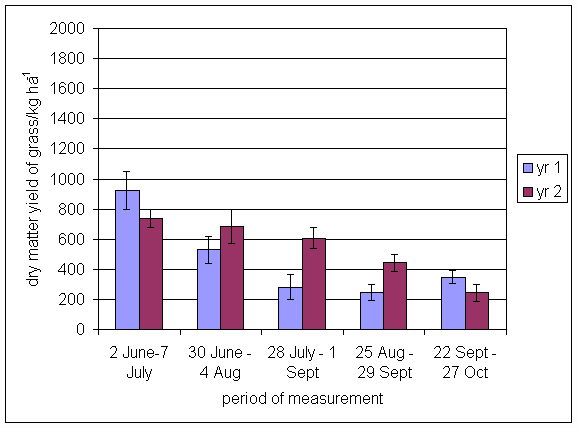
Figure .33 Grass herbage dry matter yield at Tulloch in 1- and 2-year-old leys in 1998. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 1999, grass dry matter yields for the sampling period were 2308.2, 2190.4 and 3033 kg ha-1 in 1-, 2- and 3-year-old leys, respectively. These values were not significantly different at p<0.01. Total annual grass yields (including initial sample, ungrazed plots only) were 4786.1 and 5878.5 kg dry matter ha-1 for 2-year-old and 3-year-old (ungrazed) leys, respectively. Grass yields were at a maximum in July, and subsequently declined (REML p<0.001). The seasonal variation was greatest in 3-year-old leys (REML p<0.001, Figure 4.5.34). There was no effect of rotation on grass yields in 1999.
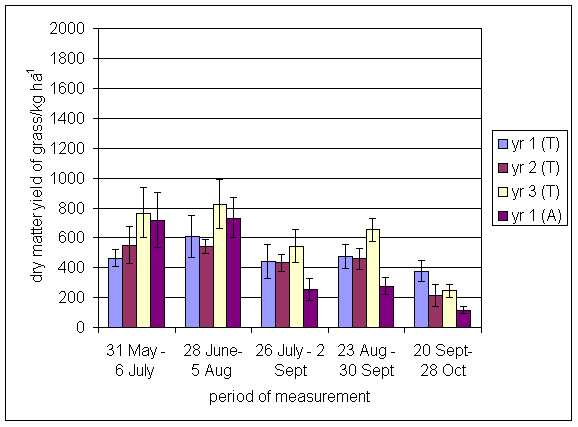
Figure .34 Grass herbage dry matter yield at Tulloch in 1-, 2- and 3-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 1999. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
In 2000, grass yields during the sampling period were 3419.2, 2578.7, 3894.9 and 3693.1 kg dry matter ha-1 in 1-, 2-, 3- and 4-year-old leys, respectively. Grass yields were significantly lower in 2-year-old leys (REML p<0.001, Figure 4.5.35). Grass yields were low at the beginning and end of the growing season (REML p<0.001, Figure 4.5.35). This seasonal pattern varied considerably with age of ley: grass yield in 3-year-old leys peaked in June, whereas grass yields in 4-year-old leys peaked in late August/ early September (REML p<0.001). Grass yield was significantly higher in the 66% ley rotation in 2000 (REML p<0.001, Figure 4.5.36).
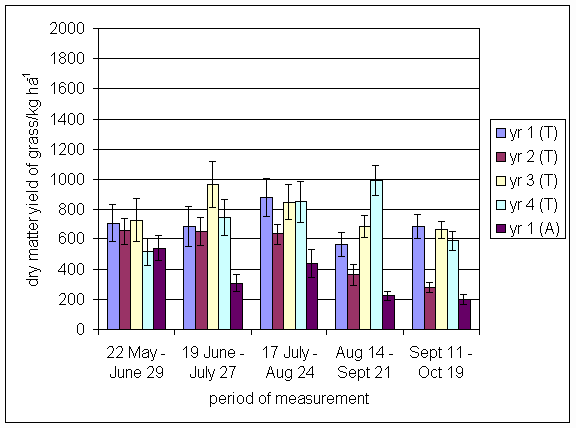
Figure .35 Grass herbage dry matter yield at Tulloch in 1-, 2-, 3- and 4-year-old leys at Tulloch, and 1-year-old leys at Aldroughty in 2000. Values represent the herbage produced during the month of sampling (mean of 12 subplots). The period of measurement is the period from the start of regrowth on the earliest treatment to be harvested, to cutting of the last treatment to be harvested in each month. Bars represent standard errors.
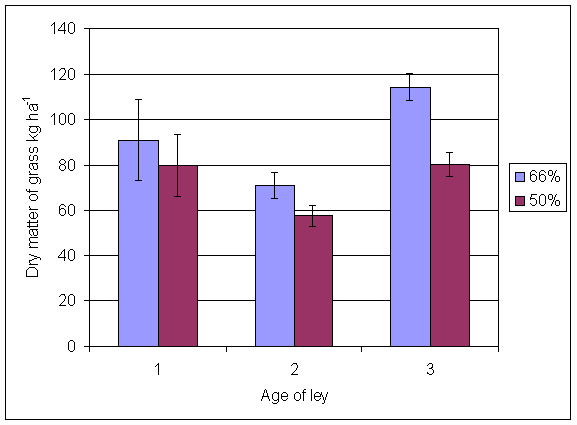
Figure .36 Grass dry matter yields in the 66% and 50% ley rotations in 2000. Columns represent total grass dry matter yield for the sampling period (mean of 6 subplots). Bars represent standard errors.
In 1-year-old leys, grass yields were highest in 1997 and 2000 (REML p=0.001). Grass yields varied seasonally, declining in the autumn (REML p<0.001), and the seasonal pattern varied significantly: in 1997, yields were very high in July, and subsequently relatively low compared to other years (REML p<0.001). There was no effect of rotation on 1-year-old leys.
Grass yields in 2-year-old leys did not vary significantly between years, although the seasonal trends (REML p<0.001) did. Grass yield was lower at the start of sampling in 1999 than in 1998 or 2000 (REML p=0.001). There was no effect of rotation on grass yields in 2-year-old leys.
Grass yields in 3-year-old leys were significantly higher in 2000 than in 1999 (REML p=0.001). Grass yield was at a maximum in July (REML p<0.001), but in 1999 it declined more sharply in late summer/autumn than in 2000 (REML p<0.001). Grass yields were higher in the 66% ley rotation in 3-year-old leys (REML p<0.001), especially early in the growing season (REML p<0.001) and early in 2000 (REML p<0.001).
Grass dry matter yields in 1-year-old leys at Aldroughty were 2102.1 kg ha-1 in 1999 and 1705.8 kg ha-1 in 2000. Dry matter yield of grass was significantly lower in 1-year-old leys at Aldroughty than Tulloch in 2000 (ANOVA p<0.001). Yield of grass was similar in both years at Aldroughty. Grass dry matter yields were similar in 1999 at both sites.
Tulloch
N transfer calculated by isotope method a) was generally low or negative at Tulloch. In 2000 pNdfaIT was similar to pNdfaET in the 1,3 and 4-year-old leys, and lower in the 2-year-old leys (Table 4.5.1).
|
Method |
Age of ley (years) |
1997 |
1998 |
1999 |
2000 |
|
Isotope method a) |
1 |
*6.30 (1.73) |
1.10 (0.66) |
0.78 (0.79) |
1.15 (0.61) |
|
2 |
-0.25 0.18) |
-5.04 (0.95) |
*-3.13 (0.65) |
||
|
3 |
1.20 (0.65) |
1.50 (2.11) |
|||
|
4 |
0.21 (0.35) |
||||
|
Isotope method b) |
1 |
0.04 (1.52) |
1.31 (0.41) |
1.84 (0.62) |
26.7 (13.4) |
|
2 |
-4.17 (0.62) |
-4.01 (1.49) |
*-29.1 (3.43) |
||
|
3 |
1.18 (0.67) |
-0.39 (2.92) |
|||
|
4 |
11.87 (8.43) |
||||
|
N difference method |
1 |
14.8(17.20) |
-1.75 (9.24) |
-13.5 (8.56) |
28.12 |
|
2 |
-36.0 (6.48) |
-58.8 (5.08) |
11.64 |
||
|
3 |
9.69 (8.25) |
19.87 |
|||
|
4 |
31.37 |
Table .1 N transfer at Tulloch (kg ha-1) between 1997 and 2000, calculated by isotope methods a and b, and the N difference method. Numbers in brackets represent standard errors.
*July-October only
N transfer calculated by isotope method b) was also frequently low or negative (Table 22). In 2000, the 1-year-old ley had positive values, the 2-year-old ley had negative values, the 3 and 4-year-old leys had values not significantly different from zero.
N transfer calculated by the N difference method produced a lot of negative values between 1997 and 1999. In 2000, grass N yield in mixtures was greater than grass N yield in monocultures in all ages of ley (Table 22). N transfered in 2000 was approximately 35% of total above ground fixed N in 1-year-old leys, 13% in 2-year-old leys, 21% in 3-year-old leys and 50% in 4-year-old leys. Excluding non-fixed N in clover, N transfer of fixed N for 2000 was 30.72, 4.96, -5.27 and 46.76 % for 1-,2-,3- and 4-year-old leys, respectively. The low and negative N transfer values in 2 and 3-year-old leys, may indicate that in these stages of the rotation, clover is successfully competing with grass for soil N. N fixation above ground calculated by N difference for these plots in 2000 gave values of 62.72, 87.28, 83.15 and 62.82 kg ha-1 for 1-, 2-, 3- and 4-year-old leys, respectively.
There was generally little agreement between the various methods for calculating N transfer. Table 4.3.1 shows the correlations between the various estimates of N transfer for 2000, when the N transfer measurements were felt to have been made with the greatest accuracy.
|
Isotope method a |
Isotope method b |
|
|
Isotope method b |
0.688 |
|
|
N difference method |
0.104 |
0.398 |
Aldroughty
In both 1999 and 2000, N transfer values calculated by isotope method a) were not significantly different from 0. N transfer calculated by isotope method b) was slightly negative in both years, and N transfer calculated by the N difference method was not significantly different from 0 in either year.
|
Method |
1999 |
2000 |
|
Isotope method a) |
-1.40 (0.59) |
-4.36 (6.21) |
|
Isotope method b) |
-2.76 (1.06) |
-3.05 (1.12) |
|
N difference method |
-0.45 (6.39) |
-1.75 (13.90) |
Flowerheads
Figure 4.5.37 shows the number of clover flowerheads per g of clover dry matter on all sampling dates in 2001. Total numbers of flowers and flowering rate averaged over the whole year are shown in Table 4.5.4.
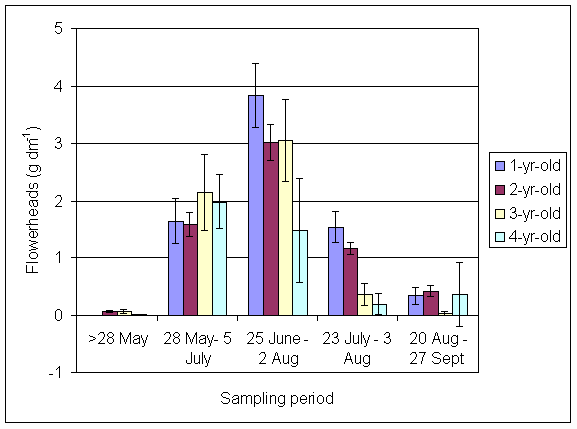
Figure .37 Number of flowers per g dried clover in samples taken for 15N analysis in 2001. Columns represent the mean of 12 subplots. Bars represent standard errors.
Flowering rate was generally highest in 1-year-old leys (ANOVA p<0.001), and in July (ANOVA p<0.001, Table 4.3.1). Flowering was over by the end of September (Figure 4.5.37). There was no direct effect of rotation on the flowering rate of clover but, in 3-year-old leys, flowering rate was higher in the 66% ley rotation (ANOVA p<0.001). The flowering rate was also around 10% higher in the 66% ley rotation in July (p<0.05).
|
Age of ley (years) |
Flowers/g clover dm |
|
1 |
2.0 (0.27) |
|
2 |
1.26 (0.21) |
|
3 |
1.51 (0.45) |
|
4 |
0.62 (0.23) |
Table .4 Number of clover flowers produced per m2 within the subplots used for the 15N dilution study. Numbers in brackets represent standard errors.
In August 2001, only clover in 1-year-old leys had significant numbers of tap-roots. No tap-roots were observed in clover from 4-year-old leys, and tap-roots were rare in 2 and 3-year-old leys (Figure 13).
These results should be taken with caution, as tap-roots were only counted in soil samples from one sampling date. Also, the results do not show what proportion of clover plants had tap-roots, but rather, the average number of tap-roots in 8 soil cores taken from clover microsites.
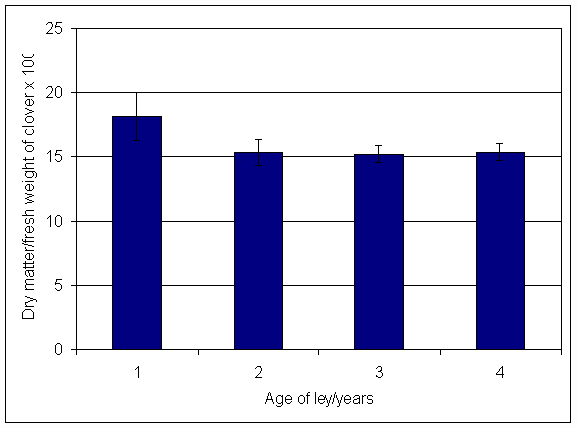
Figure .38 Dry matter content of clover herbage in 2001, expressed as a percentage of fresh weight. Columns represent average dry matter content for the whole year (mean of 4 subplots).
Other differences were also observed between different ages of ley. In 2001, clover in 1-year-old leys was observed to have a higher dry matter content than clover in 2, 3 and 4-year old leys (Figure 4.5.38)
.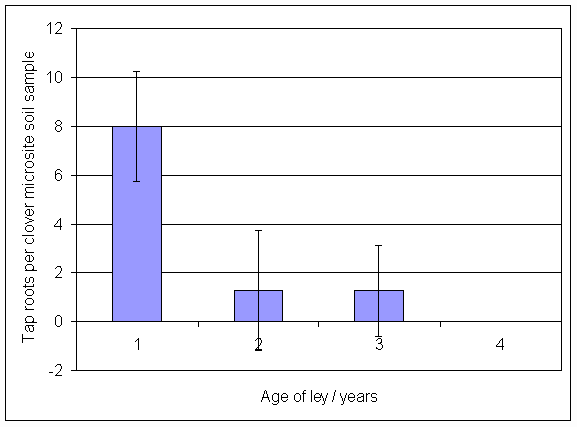
|
Year |
Age of ley |
N in grass in mixture (kg ha-1) |
N in grass monoculture (kg ha-1) |
Fixed N in clover (kg ha-1) |
Soil N in clover (kg ha-1) |
Total N in herbage (kg ha-1) |
|
1997 |
1 |
121.5 |
121.8 |
41.8 |
11.1 |
151.7 |
|
1998 |
1 |
54.3 |
57.7 |
26.2 |
1.6 |
82.2 |
|
2 |
67.4 |
105.1 |
30.3 |
1.7 |
99.4 |
|
|
1999 |
1 |
44.3 |
93.0 |
61.9 |
2.9 |
109.1 |
|
2 |
41.8 |
103.6 |
75.6 |
4.5 |
121.9 |
|
|
3 |
61.2 |
55.5 |
47.6 |
4.0 |
112.8 |
|
|
2000 |
1 |
103.5 |
86.6 |
73.7 |
7.9 |
185.1 |
|
2 |
66.5 |
72.8 |
70.4 |
6.1 |
143.1 |
|
|
3 |
126.1 |
155.0 |
69.2 |
12.8 |
208.2 |
|
|
4 |
110.1 |
86.4 |
33.5 |
5.7 |
149.3 |
|
|
total |
360.2 |
368.8 |
153.2 |
22.5 |
513.2 |
Table .5 N budgets in all years and ages of ley, showing fixed and soil derived N present in all grass and clover mixtures and monocultures. Data from all plots.
|
Age of ley |
N in grass mixture (kg ha-1) |
N in grass monoculture (kg ha-1) |
Fixed N in clover (kg ha-1) |
Soil N in clover (kg ha-1) |
N benefit to grass from clover (kg ha-1) |
|
1 |
110.1 |
86.6 |
53.1 |
4.6 |
23.5 |
|
2 |
76.8 |
72.8 |
75.6 |
7.7 |
4.0 |
|
3 |
147.0 |
155.0 |
76.1 |
15.1 |
-8 |
|
4 |
114.1 |
86.4 |
31.4 |
3.8 |
27.7 |
|
total |
448 |
400.8 |
236.2 |
31.2 |
47.2 |
Table .6 N budget for plots 13 (4-year-old ley), 16 (1-year-old ley), 17 (2-year-old ley) and 18 (3-year-old ley) showing fixed and soil derived N present in all grass and clover mixtures and monocultures.
N budgets for the subplots are shown in Table 4.5.5 and Table 4.5.6. Table 4.5.6 shows the N budget for selected subplots in 2000. lt was difficult to calculate N transfer in this experiment with any confidence. The N transfer calculations at Tulloch in 2000 were an attempt to improve the accuracy of the method, by reducing the distance between the grass-clover subplots, and the grass monocultures. For this reason, this section will mainly focus on the data from Tulloch in 2000.
The data for 2000 shows that harvesting of the subplots in the 1-,3- and 4-year old leys removed more soil N from the system than was fixed by clover, and considerably more than was apparently transferred from clover to grass. Grass yields are a low estimate, as there was considerable early season grass growth, equal to around 50% of the grass yield during the sampling period. For this reason, grass yields will be multiplied by 1.5 and clover yields by 1.3 in the following calculations: In the cut leys the net removal of soil N in grass and clover over the growing season (taking into account N fixation below ground) was 49.4, 181.9 and 110.7 kg N ha-1 for 2, 3 and 4-year-old leys, respectively. Transfer of fixed nitrogen from clover calculated as N difference was 4, 0 and 27.7 kg N ha-1 for 2, 3 and 4-year-old leys, respectively. These represent conditions under the exclusion cages. In the field, the effects of grazing and FYM applications must also be taken into account:
Applications of manure to cut plots in 2000 were 23, 10 and 15 tonnes ha-1 in 2-, 3-, and 4-year-old cut leys, respectively. Assuming that composted organic FYM contains 25% dry matter and 5.9 % N (Watson, 2003), this means that 339.25, 147.5 and 221.25 kg N were applied as manure per hectare in 2-, 3- and 4-year-old cut leys, respectively. According to Granstedt (1995), approximately 58% of this will be lost to the atmosphere, making the manure inputs 142.5, 61.95 and 92.9 kg N ha-1 in 2-, 3- and 4-year-old cut leys, respectively. This makes the approximate balance of N (inputs - output) 97.1, -127.9 and 9.94 kg N ha-1 in 2, 3 and 4-year-old cut leys, respectively (including N fixation below ground, estimated at 70% of fixed N in herbage). This calculation does not take leaching into account.
In grazed leys around 25% of the nitrogen removed by grazing animals is likely to be retained by the animals, the rest returned as excreta (Petersen et al., 1956), although 58% of this excreta N is likely to be lost by volatilisation and leaching (Petersen et al., 1956; Granstedt, 1995). This would amount to a net removal of 59.3 and 106.18 kg N ha-1 of soil derived N from grass and 3.14 and 10.3 kg N ha-1 of soil derived N from clover in 1- and 3-year-old leys, respectively. Assuming that 75% of the fixed N in the clover herbage will be returned as dung and urine by the grazers, and 58% of this will be lost to air and water, this means that 40.2 and 39.56 kg fixed N ha-1 will be returned to the soil in 1 and 3-year-old leys, respectively (including N transferred from clover to grass, in 1-year-old leys). In addition, N fixed in roots, stolons and stubble will be equal to approximately 70% of the fixed N in leaves and shoots of clover (Jorgensen and Ledgard, 1997). This will add an extra 37.16 and 53.3 kg N ha-1 to 1- and 3-year-old leys respectively. This gives a total N balance of 14.92 and –33.9 kg N ha-1 for 1- and 3-year-old leys, respectively. Ledgard (1991) estimated that in a sward grazed by dairy cattle in New Zealand, 22% of fixed N was transferred above ground from clover to grass. In the same study, below-ground transfer was estimated at 70 kg N ha-1 (26% of fixed N).
This method can also be used to estimate the N budget for plots 3, 10, 13 and 19 over the 4 years of the study, as these plots were 1-year-old in 1997, 2-year-old in 1998 and 3-year-old in 1999. Plots 3 and 13 were also 4-year-old in 2000. Manure applications to 2-year-old leys in 1998 were 30 t ha-1. Three-year-old cut leys in 1999 received 23 tonnes manure per ha and 4-year-old leys received 15 tonnes manure/ha. This means that the 50% ley rotation accumulated 433.8 kg N ha-1 between 1997 and 1999, and the 66% ley rotation accumulated 370.2 kg N ha-1 between 1997 and 2000. The 66% ley rotation accumulated less in the ley, but this was because of the high amount of grass removed in the 4-year-old leys in 2000, and the manure applied to the 50% ley rotation 3-year-old plots in 1999. Much of the N in grass and clover removed from these two rotations would subsequently be returned to the plots in the arable phase in the form of manure. First year oats at Tulloch received no N, 2nd year oats received 12 tonnes, and swedes received 12 tonnes. This means that 177 kg N ha-1 are returned to the 66% ley rotation and 354 kg N ha-1 are returned to the 50% ley rotation in the form of manure. 3.5-3.9
These estimates do not take into account grazing and decay of clover over winter. The evidence from the grass N yields (Table 4.5.6) suggests that N is accumulating over the first three years of the ley.
This section of the experiment was intended to test the following hypotheses:
N fixation did not show a simple steady decline with age of ley. N fixation was generally highest in 2-year-old leys. In 2000 there was evidence of reduced N fixation in 4-year-old leys. The variation in N fixation between years was mainly due to variation in the yield of clover, and to a lesser extent, variation of clover N concentration and pNdfa. Clover N concentration was lowest in 2-year-old leys and highest in 3-year-old leys in 1999 and 2000. Clover N concentration varied considerably more than pNdfa and therefore had more of an effect on N fixation. Significant variation in clover N concentration between treatments has not been observed in many other studies of this type, and most studies have observed that clover N concentration is unaffected by addition of N (e.g. Boller and Nosberger, 1987, Høgh-Jensen and Schjoerring, 1994, Jorgensen et al. 1999). Høgh-Jensen et al. (2002) found that low availability of phosphorus could reduce the N concentration of clover herbage.
Grass yield and N concentration did not simply increase with age of ley either. In 1999 and 2000, these values were relatively low in 2-year-old leys and relatively high in 3-year-old leys. Soil derived N was often similar in 1- and 2-year-old leys. pNdfa did not steadily decline with increasing age of ley. It was relatively low in 3-year-old leys, but similar in 1-, 2- and 4-year-old leys.
In general, differences between the two rotations were small. N fixation was not reduced by the extra year of ley in the 66% ley rotation, instead it was actually often higher in 1 and 3-year old leys. The elevated N fixation in these cases was due mainly to higher clover dry matter yields, but also partly to higher N concentration in the clover. In contrast, pNdfa was often reduced in the 66% ley rotation in 1 and 3-year-old leys. It is possible that these differences may have been due to grazing of the 3-year-old leys in the 66% ley rotation prior to sampling, rather than to any long term effects on soil fertility.
SDN, grass yield and grass N concentration were also often higher in the 66% ley rotation in 1 and 3-year-old leys. SDN and grass yield were significantly higher in the 66% ley rotation overall. This provides some evidence for higher available N in the 66% rotation, but the effects may be due partly to the management of the leys prior to sampling, as the effects of rotation were most apparent in grazed leys.
Over all year and age treatments, total N fixation ranged from 26.2 kg N ha-1 in 1998 1-year-old leys to 75.6 kg N ha-1 in 1999 2-year-old leys. This was mainly due to large variations in the annual yield of clover per hectare (756 kg ha-1 in 1998 1-year-old leys and 2810 kg ha-1 in 1999 2-year-old leys). Average clover N concentration ranged from 2.87 in 1999, 2-year-old leys to 4.38 in 2000, 3-year-old leys. Average pNdfa varied considerably less (ranging from 0.854 in 2000, 3-year-old leys to 0.952 in 1999 1-year-old leys). Variation of clover yield clearly accounts for most of the annual/age related variation of N fixation at Tulloch. Variation of pNdfa had remarkably little effect on N fixation. Variation of clover N concentration was significant, but high clover N concentration often coincided with low clover yield, so the effect of clover N concentration on N fixation was often masked.
Total annual soil derived N in grass and clover ranged from 26.2 kg ha-1 in 1998, 1-year-old leys to 139 kg ha-1 in 2000, 3-year-old leys). Grass yield ranged from 2190.4 kg ha-1 in 1999 2-year-old leys to 3894.9 kg ha-1 in 2000 3-year-old leys. Grass N concentration varied from 1.9% in 1999, 2-year-old leys to 3.19% in 2000 3-year-old leys. Most of the soil derived N was in the grass, with a smaller fraction in the clover: Soil derived N in clover comprised less than 3% of total soil derived N in 1998, but in the other years soil derived N in clover could account for up to 10% of total soil derived N. This suggests that annual changes in soil N are more likely to cause changes in the grass:clover ratio than change the amount of soil N utilised by clover. Hansen and Vinther (2001) measured soil derived N in clover that was less than 2% of soil derived N. Danso et al. (1988) observed that soil derived N in white clover was only 3.1% of total soil derived N in a white clover-fescue pasture. In contrast, Boller and Nosberger (1987), calculated soil derived N in clover, without added N in 1-year-old leys at 73 and 97 kg ha-1 in 1984 and 1985, respectively, and total grass N was 82 and 95 kg ha-1 in 1- and 2-year-old leys, respectively. This means that in their experiment, SDN in clover was 47.1-56.5% of total SDN (using the methods of this study), suggesting that in their experiment, clover was competing far more effectively with perennial ryegrass for soil N than in our experiment. Hogh-Jensen and Schjoerring (1997) observed that soil derived N in white clover accounted for between 5.9 and 17.2% of total soil derived N.
Annual variationGrowth of grass and clover and the ratio of grass to clover are likely to be affected by the amount of sunlight available in any particular year. The competition between grass and clover for light is likely to be affected by the sward height, as ryegrass grows taller than clover when permitted to (Harris, 1987). Ryegrass grows at slightly lower temperatures than clover (Chestnutt and Lowe, 1970), and clover in Britain needs temperatures of 9o C. to fix nitrogen (Munro, 1970). The result of this is that clover tends to start growing later in the season than ryegrass (Ollerenshaw and Baker, 1981), and early season competition between grass and clover is likely to be affected by availability of soil nitrogen as well as temperature. Soil moisture also affects growth of grass and clover. Clover is shallower rooting than ryegrass and therefore more prone to drought (Harris, 1987, Caradus, 1990). It would be expected that variations in rainfall between seasons would affect the growth of grass and clover (e.g. Schils et al., 1999), but a comparison of the N fixation and soil N data with the rainfall and temperature data for the site does not reveal any obvious patterns. 1998 and 2000 were wetter years than 1997 and 1999, and 2000 was colder than the other years studied (Figure 3.1.2-Figure 3.1.5). Neither rainfall or temperature were simply related to annual variation of N fixation, clover yield, clover N concentration, pNdfa, soil derived N in herbage, grass yield or grass N concentration.
Clover N concentration and pNdfa followed clear seasonal patterns that were remarkably consistent in different sites, ages of ley, and years. pNdfa tended to decline late in the growing season, and a similar pattern was observed by Boller and Nosberger (1987). N concentration of clover was lowest in July and highest in September. In 2001, flowering rate was also highest in July, except in 4-year-old leys, which peaked in June. Assuming flowering rate followed a similar pattern in previous years, it would be negatively correlated with N concentration during the growing season. If intact white clover flowerheads prior to seed formation have a lower N concentration than leaves and petioles, this could explain the seasonal variation of clover N concentration and the variations of this pattern between different ages of ley. Warembourg et al. (1997) observed that flowers of red clover had a C:N ratio of approximately 15, lower than any other part of the plant, save nodules. However, the C:N ratio of flowering stems varied enormously from 10 to 70, and 100 when dead. Hogh-Jensen et al. (2001) observed that N concentration in clover was higher in petioles than in any other plant part. It is therefore possible that flowering might cause an overall decline in white clover N concentration.
Regular cutting increases clover N concentration (Elgersma et al. 2000), probably by suppressing flowering and stimulating new growth, so the N concentration values and flowering rates may not actually reflect the real situation in the field, especially in cut leys. If regular cutting increases clover N concentration, then it is likely that grazing might have the same effect. This could explain the elevated clover N concentration early in the season in 3-year-old leys in the 66% ley rotation, as these leys were grazed prior to the start of sampling.
Seasonal variation of grass N concentration may have been due in part to the flowering cycles of the grasses. Generally grass N concentration rose over the course of the growing season. Similar trends were observed by Elgersma et al. (2000).
Flowering of grasses was not measured in this experiment.
N fixation and pNdfa followed roughly opposite patterns to grass yield with respect to age of ley. N fixation was generally highest in 2-year-old leys, grass yield, soil derived N and grass N concentration were lowest in 2-year-old leys. pNdfa was lowest in 3-year-old leys, and grass yield was highest in 3-year-old leys. However, there was no clear inverse correlation between these factors. Dry matter of clover followed a similar pattern to N fixation (Figure 4.5.5-Figure 4.5.8, Figure 4.5.18-Figure 4.5.21).
It was hypothesised that the pattern of clover N concentration in different ages of ley could be explained by differences in the rate of flowering of clover with age of ley, possibly related to the life cycle of clover. When a clover plant makes the transition from a tap-rooted rosette to a creeping stoloniferous form, it would be expected that there would be changes in the proportion of stolon, leaf, flower and petiole that could cause changes in the N concentration of the sampled herbage (Warembourg et al., 1997). Clover in leys of different ages did show different rates of flowering. It should be stressed that this was only apparent in the controlled conditions under an exclosure cage. In the field, it was obvious that there was far more clover in flower in cut leys than in grazed leys, as sheep grazing consumed a large number of flowerheads. Flowering rate tended to decline with age of ley, and so did not appear to correlate with the clover N concentrations observed in different ages of ley in 2000, as clover flowers are rich in N (Warembourg et al., 1997). Thomas (1987) observed that clover plants fed entirely on mineral N produced more flowers than those fixing N, when grown with 16h of daylight in every 24 hours (Thomas, 1987).
Leys of different ages had different pNdfa values, but this had little effect on the N fixation values, which closely followed the pattern of clover yield (Figure 4.5.1-Figure 4.5.4, Figure 4.5.5-Figure 4.5.8, Figure 4.5.18-Figure 4.5.21). Grass N concentration followed a similar pattern to clover N concentration in leys of different ages, which could mean that both clover and grass are responding in a similar way to soil conditions in leys of different ages. From the limited data available, it seems that clover in this system loses its taproots between the first and second years of ley (when the clover plants are between two and three years old). The senescence of taproots immediately precedes the peak of clover yield, and a sharp reduction in clover N concentration. Frequency of taproots appears to follow a similar pattern to flowering of clover, both declining with age of ley in August. Pederson (1989) observed that clover plants grown with additional N had thicker taproots, more, longer and thicker stolons and more flowerheads than those without additional N. Ryle et al. (1981b) also observed enhanced growth of clover plants with added N, although this was in an experiment in which clover was grown as a monoculture, so the effects of elevated soil N and competition on clover morphology could not be observed. Neither of these experiments ran for long enough to observe loss of taproots, and the clover plants were not grown with grass, so the response of the clover to intense competition from grass in an N rich soil could not be measured. It would be expected that with additional nutrients and no competition, clover would show a general increase in size. It would be interesting to repeat the experiment using a grass clover mix, and a variety of N treatments, over a 4-year period. The disappearance of taproots could be observed either with a minirhizotron, or using destructive sampling. Stolon mass, flowering rate and/or seed yield of the clover, could also be measured, to test the hypothesis that clover alters its reproductive strategy in response to soil N and competition.
Data on ground cover of various species at Tulloch (Figure 4.5.22) broadly supports the evidence of an age-related pattern observed in this study. Overall, 2-year-old leys have most clover. However when the pattern is viewed over all years since conversion, it becomes clear that there have been considerable fluctuations over time. This is to be expected, as the plots were going through their first six-year rotation cycle since conversion while these measurements were being taken. For some reason, 1-year-old leys are less subject to these fluctuations than other ages of ley. This may be because the clover content in 1-year-old leys is determined more by the seeding rate than by any ecological factors. High clover seeding rates can improve white clover establishment, but the effect is overridden after a couple of years (Frame and Newbould, 1986).
It was not possible to directly measure the effect of cutting and grazing on nitrogen fixation, clover yield, N concentration of clover, pNdfa, soil derived N in herbage, grass yield or grass N concentration in this study. These variables were only measured in the experimental subplots which were all cut once every four weeks, and so unaffected by the cutting and grazing treatments of the surrounding plots. However, certain rotation effects were only detected mainly in 3-year-old leys (Age of ley x Rotation interactions), and were most prominent early in the year. It is possible that these result from grazing of the 3-year-old leys in the 66% rotation before sampling began. It is reasonable to suggest that in this system, grazing increases soil derived N, increases grass N concentration and dry matter yield, and reduces pNdfa. All of these effects could be caused by the above ground N transfer of N from clover to grass, and elevated soil N. Alternatively these effects could be due to the type of rotation. Schils et al. (1999) observed less white clover ground cover in a management system using rotational grazing and cutting than one with cutting only.
Across all years and ages of ley, there was evidence that N concentration of grass was elevated in the 66% ley rotation. There was also evidence of elevated N fixation, %NC, SDN and %NG, and reduced pNdfa in the 66% ley rotation, mainly in 1 and 3-year-old leys, in some years. There was no difference between the two rotations in their clover N concentrations. Other evidence from the site showed that there was slightly more K in the 66% ley rotation and this effect was most pronounced in 1999 and 2000 (data in appendix).
Differences between the two rotations tended to be small. N fixation provided by the extra year of ley in the 66% ley rotation was not great, although the extra grass yield could be seen as a benefit to be offset against the loss of 1 years arable crop from the rotation.
N fixET, pNdfa, grass N yield, clover dry matter and average clover N concentration were similar at Tulloch and Aldroughty. There were significant differences between grass N concentration and yield at Aldroughty and Tulloch. This may reflect differences in soil fertility or rainfall between the two sites. Between 1997 and 1999, Aldroughty had significantly lower P (REML p<0.001) and K (REML p<0.001) than Tulloch, and significantly higher magnesium (REML p<0.001, data in appendix).
A deficiency of any of these nutrients could reduce N fixation. Legumes are known to have a higher demand for P & K than non-legumes, so a deficiency of these nutrients would be expected to reduce the clover:grass ratio The low grass N concentration may have been due to the relatively high proportion of broad-leafed weed species (Rumex spp., thistle, volunteer potato etc.) included in the grass fraction at Aldroughty, compared to Tulloch. The flowering rate of clover was not measured at Aldroughty, so a comparison could not be made.N fixation tended to be higher in block 1 at Tulloch than in block 2 (Data in appendix). This may be because block 2 was slightly higher than block 1, more sloping and drier. Ledgard et al. (1987)
found lower N fixation by white clover on sloping sites, although this may have been due to competition from other legume species, which were not at Tulloch. The increased fixation manifested as an increase in clover yield, N concentration and pNdfa were similar in the two blocks. There were no differences in flowering rate of white clover between different blocks at Tulloch. Nitrogen concentration of clover and grass was not significantly different between blocks.Isotope method a) only considers differences between pNdfaIT and pNdfaET, and not differences in N yield of grass mixtures and monocultures. It is rarely used in the literature. In this experiment, differences between pNdfaIT and pNdfaET were negligible
. Method a) compares isotopic enrichment in clover and grass in mixtures and monocultures, but does not consider differences in N yield of grass in mixtures and monocultures. Isotope method b) produced high estimates of N transfer for 1 and 4-year-old leys (36 and 47% of total N fixed, respectively), but 2-year-old leys had negative N transfer values, and 3-year-old leys had non-significant N transfer, calculated by this method. The N difference method also estimated N transfer to be highest in 1- and 4-year-old leys (35 and 50% of total N fixed respectively), but gave large estimates for the 2 and 3-year-old leys when non-fixed N in clover was taken into account (13 and 21 %, respectively). There is therefore some agreement between isotope method b) and the N difference method. The N difference method measures difference in nitrogen yield between grass in mixtures and monocultures, but this does not prove that any additional nitrogen in the grass in mixtures actually originated in the clover. Grass grown in mixtures often had a similar or even lower dry matter yield than grass in monocultures, but a higher N concentration. The effect of clover on grass is probably not a simple one. Discrepancies between the isotope and N difference methods could indicate that clover benefits ryegrass in other ways besides increasing nitrogen availability. Nitrogen released from clover roots might increase the turnover of soil organic matter and release soil nitrogen for grass to use. Any transfer of nitrogen from grass to clover might also complicate the picture, although there is little evidence for this happening in the literature.With all these methods, variation within the plots could still have caused considerable errors, and for the sake of accuracy, grass monoculture subplots should be immediately adjacent to the mixed subplots. The grass monoculture subplots were created by weeding out the clover, prior to sampling. This may well have left fragments of root and nodules to decay in the monocultures, increasing soil N and reducing estimates of N transfer.
There may have been N transfer by the above ground pathway which could not have been measured in this experiment, and likewise, there may also have been N transfer occurring over the winter.
The high levels of soil N, high grass N yield and grass N concentration in the 3-year-old leys at Tulloch, suggests that there may have been considerable N transfer from clover to soil taking place over winter at the end of the 2nd year of the ley rotation. This may have been due to the large volume of clover at this stage, a proportion of which would inevitably decay or be grazed. Alternatively, high levels of soil N could be released from the clover by the senescence of the clover tap-roots which occurs after 1-2 years of growth (Caradus, 1990). In order to measure total N transfer over the whole course of the ley phase, it would be necessary to maintain grass monocultures for 3-4 years. In an organic system, where herbicides cannot be used (Soil Association, 2000a), this obviously creates problems. Most studies used either grass monoculture sown specially for the purpose, or monocultures created using herbicides. Removing clover by hand may have caused disturbance to the soil, or left fragments of clover to decay into the soil, but herbicide treatments could also result in the decay of clover tissue in control plots.
The NfixET values are probably therefore a low estimate, even when N fixation below stubble height, and N fixation outside the growing season are included. Considerable N transfer within the growing season occurred in some ages of ley at least, but the values for NfixIT and pNdfaIT (and therefore N transfer), could not be relied upon, so below ground N transfer is still largely an unknown quantity. If the N transfer values obtained by the difference method are assumed to be reliable, then total N fixation including transfer, below ground fixation, and fixation outside the sampling period for 2000, can be estimated at 214, 164, 146 and 109 kg N ha-1 for 1-, 2-, 3- and 4 year-old leys, respectively. The equivalent estimates for the previous years, range from 71 in 2-year-old leys in 1998, to 176 in 2-year-old leys in 1999. These estimates do not take into account N transfer over winter, or fixed N immobilised in soil, so the true figure could be even higher.
Estimates of N transfer vary considerably: Hatch and Murray (1994) used 15N labelling to measure transfer of N from clover to grass, and found that N transfer only occurred under these conditions if the roots of the clover were damaged. There is some evidence that different clover varieties transfer different amounts of N. Laidlaw et al. (1996) found that small leaved varieties such as Kent wild white transferred more fixed N (27.66% of total N fixed), than large-leaved varieties such as Aran (17.66%. of total N fixed). Thorsted et al. (2002), observed that variety S184 transferred more of its fixed N to oats grown alongside it, than other clover varieties such as Aran which yielded slightly more.
Jorgensen et al. (1999) used 15N dilution methods to measure N transfer from white clover to ryegrass. In the seeding year (when clover was undersown with spring barley), N transfer was negligible, but in the first and second production years, N transfer was 19 and 28 kg N ha-1, respectively, similar to the values obtained for 1-year-old leys in our study. Other studies have also found low N transfer in the seeding year, and increasing N transfer subsequently: Laidlaw et al. (1996) observed that N transfer was on average of 14.6% of fixed N (excluding root and stubble) in the seeding year but rose to 31.8% in the third year of growth. Elgersma and Hassink (1997) also using an N difference method over 4 years, found that N transfer was low in the seedling year (average 14.7% of total N fixed), but was 25.5, 33.8 and 24.4% of N fixed in the 2nd, 3rd and 4th years of growth, respectively. McNeill and Wood (1990), using 15N dilution, estimated N transfer from white clover to be 21.5 – 25% of total N fixed. Boller and Nosberger (1987) found that N transfer from white clover to perennial ryegrass, estimated by isotopic methods, ranged from 4.6 – 30.3% of total N fixed in the seedling year, and 14.9 – 15.5% in the 2nd year of growth. Høgh-Jensen and Schjoerring (2000) measured N transfer from white clover to perennial ryegrass using both leaf labelling and 15N dilution. Leaf labelling produced transfer values of 45 and 75 kg N ha-1 in the 1st and 2nd production years, respectively. Estimates using the 15N dilution technique were approximately 50% lower.
In contrast, this study found low N transfer values (measured by all three methods) in the 2- and 3-year-old leys, but higher values in the 4-year-old leys. The N transfer values produced by the N difference method seem to be reasonably consistent with those found in other studies, but the isotope-based estimates were not.
Soil chemistry of grass-clover swards
Aims
The cyclical replacement hypothesis predicts that as the age of ley increases, soil N should increase and clover should retreat to a diminishing number of low N microsites (the term "microsites" is used here to mean patches within the sward at the scale of individual grass and clover plants). Soil N is also predicted to increase over the course of each growing season. Depletion of nutrients such as Phosphorus (P), Potassium (K) and Magnesium (Mg) would potentially have the same effect as elevated N: giving grass a competitive advantage over clover. It would be expected that the 66% ley rotation would have higher levels of readily available soil N because this rotation has a longer fertility building ley phase and correspondingly shorter arable phase.
Soil was sampled and analysed to compare concentrations of available soil N levels over time, in different ages of ley, under different management regimes, and in different rotations, to see if any of the variations in grass and clover behaviour could be attributed to variation of soil nitrogen. Phosphorus, potassium and magnesium levels were also examined, to a more limited extent for a similar purpose.
Soil N under grass and clover microsites was also analysed and compared over time, to test the hypothesis that clover colonises microsites which are low in soil N, and elevates soil N locally, before being out-competed by grass. As clover is sensitive to the combination of low temperatures and high soil N, it can be predicted that clover will die off back from microsites with high levels of available soil N during the colder months. Root and nodule turnover is likely to be highest when clover is actively growing, so it would be expected that any elevation of soil N in microsites occupied by clover would be most obvious during the summer.
Soil was sampled approximately every 2 months from the trial rotation ley plots at Tulloch (Section 3.1). Plots were not sampled in the autumn immediately after harvest of the oats. Samples were taken on a grid basis, from the top 150 mm of soil. From each plot, 8 soil cores were taken from microsites containing only grass and weeds, and 8 cores were taken from microsites containing clover. Grass cores and clover cores were bulked to provide one grass soil sample and one clover soil sample for each plot. Cores were crumbled and any plants and roots were removed.
A sample of fresh soil from each sample (30-35g) was weighed, dried at
105 oC, and weighed again in order to measure the dry matter content of the soil. 10g of each bulked soil sample was accurately weighed into a screw topped plastic bottle and mixed with 50 ml of 0.5 m K2SO4. The bottles were then placed in a mechanical end-over-end shaker for 2 hours. The soil solutions were then filtered through a fluted filter paper (Whatman No. 42). and analysed for NH4-N and NO3-N as described below. A 50 ml sample of K2SO4 was also included as a blank. A 5 ml aliquot of each filtered extract was accurately pipetted into an autoclaveable universal bottle. 1 ml of persulphate oxidiser, made from 1.34g K2S2O8 + 0.3g NaOH in 100 mls of water, was added to each aliquot, and the mixture was autoclaved at 110oC for 30 minutes (Cabrera and Beare, 1993). The processed extract was then analysed for NO3-N at a dilution of 1.5 ml of extract to 0.09 ml of 0.1M NaOH as described below.
The extracts and the digested aliquots were accurately pipetted into 2 ml autoanalyser cups and analysed for nitrate-N and ammonium N using a Carlo Erba continuous flow autoanalyser (CE Instruments, Milan, Italy). The autoanalyser converts nitrate to nitrite in the presence of copper and cadmium and then measures the nitrite calorimetrically, at 540 nm by a diazotization reaction. Ammonia is measured by reacting ammonia with hypochlorite ions to form monochloramine, which is reacted with sodium salicylate in the presence of sodium nitroprusside to form an emerald-green, indo-phenol type compound which is measured calorimetrically at 660 nm. The autoanalyser produces measures of nitrate and ammonia as ppm N in solution.
The nitrate-N concentration of the digested samples gave a value for total soluble N (TSN). Soluble organic nitrogen (SON) was calculated from TSN –(ammonium + nitrate-N). Soil samples from 17th August 2001, were also analysed for total extractable P, K and Mg. Samples were analysed immediately whenever possible, or frozen for analysis at a later date.
Standards for nitrate were prepared as follows: 0.9025g Potassium nitrate (dried at 98oC for 1 hour), was dissolved in 1000 ml of de-ionised, distilled water, to produce a 125 ppm stock solution. This stock solution was then diluted to produce standards ranging from 0.1 to 2 ppm NO3--N.
Standards for ammonium were prepared as follows: 0.4722g of ammonium sulphate (dried overnight at 98oC was dissolved in 1000 ml deionised water, to give a 100 ppm stock solution. This stock solution was diluted to produce standards ranging from 0.1 to 2 ppm NH4+-N.
Weight of oven dried soil = (weight of container + oven dried soil) – weight of container. [.1]
Weight of moisture = (weight of container + fresh soil) – (weight of container + oven dried soil) [.2]
Dry weight content = ((weight of container + dry soil)-weight of container) / ((weight of container + fresh soil) – weight of container) [.3]
Weight of oven dried soil/g = fresh sample weight/(1+(% moisture/100)) [.4]
where the fresh sample weight is 10g
In order to convert the output of the autoanalyser (ppm per sample) into kg N ha-1 in the top 150 mm of soil, the following calculation was made:
![]() [.5]
[.5]
Where the dilution factor is 50, the fresh sample weight is 10g and the sample depth was 15 cm
Bulk density was calculated as:
Bulk density = soil dry wt/volume of fresh soil [.6]
Sampling was performed on 28th January 1999, 8th April 1999, 21st June 1999, 16th August 1999, 12th October 1999, 17th January 2000, 28th February 2000, 6th April 2000, 4th July 2000, 27th September 2000, 29th November 2000, 4th April 2001, 18th June 2001 and 17th August 2001.
Nitrate-N, ammonium-N, SON and TSN data from all sampling dates was transformed and analysed by REML. Model terms were: age of ley, management type (cutting or grazing), season (month when sample was taken), rotation (50% or 66%), vegetation (grass or clover microsite) and year (1999, 2000 or 2001). Soluble organic N and ammonium data from June 1999 was omitted as the high ammonium values on this date (10-20 kg N ha-1, compared to c.5 kg N ha-1 in April and August 1999) suggested contamination of the K2SO4 solution. Samples where SON was negative (i.e. TSN values were inconsistent with nitrate-N and ammonium values) were also omitted. Nitrate-N values were rooted to the fourth power and ammonium and TSN were transformed by Log Fisher, to normalise the data.
Nitrate-N was generally higher under clover patches than under grass patches (REML p<0.01). This effect was far more pronounced in summer than in winter (REML p<0.001, Figure 5.4.1). Nitrate-N was approximately twice as high in summer 2000 as in summer 1999 (Figure 5.4.1, REML p<0.001).

Figure .1 Nitrate-N in soil under grass and clover microsites in all ley plots at Tulloch, between February 1999 and August 2001. Points represent means of 14 samples. Bars represent standard errors.
Nitrate-N was significantly lower in 1999 than in 2000 (REML p<0.001), especially during the growing season (REML p<0.001). Nitrate-N was 3-4 times higher during the winter than in the growing season (REML p<0.001, Figure 5.4.1 and Figure 5.4.2).
Nitrate-N was highest in 3-year-old leys and lowest in 2-year-old leys. (p<0.001). During the growing season, 3-year-old leys had approximately twice as much nitrate-N as 2-year-old-leys, but the difference was smaller early in the year (REML p<0.001). The age related pattern varied from year to year: nitrate-N in 1 and 4-year-old leys was relatively high compared to other ages of ley in 2000 and 2001, and relatively low compared to other ages of ley in 1999, (REML p<0.001, Figure 5.4.2).
During early summer, grazed leys had approximately 50% higher nitrate-N than cut leys (REML p<0.001, data in appendix). Nitrate-N levels were similar in both rotations.
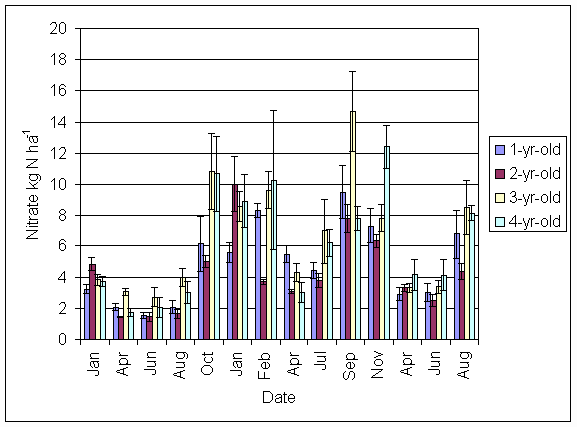
Figure .2 Nitrate-N in soil in 1-, 2-, 3- and 4-year-old leys at Tulloch between January 1999 and August 2001. Columns represent means of 8 samples (1-, 2- and 3-year-old leys) and 4 samples (4-year-old leys). Bars represent standard errors.
Soil ammonium-N levels were highest in 2000 and lowest in 1999 (REML p<0.001, Figure 5.4.3), and varied from month to month, being 2-3 times as high at the end of the growing season (REML p<0.001), although this effect was less pronounced in 1999 (REML p<0.001, Figure 5.4.3).
Ammonium-N was highest in 3 and 4-year-old leys, approximately 50% higher than in 1 and 2-year-old leys (REML p<0.001, Figure 5.4.3). Ammonium was not significantly different in cut and grazed leys. Management (cutting or grazing) did not significantly affect ammonium. Clover and grass patches did not have significantly different ammonium levels. The 66% ley rotation had up to 50% higher ammonium than the 50% ley rotation (REML p<0.001, Figure 5.4.4).
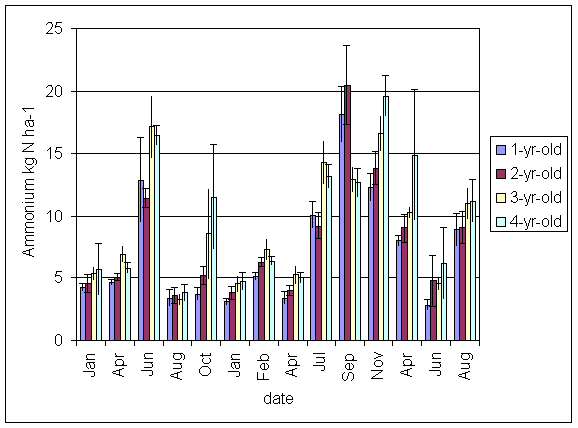
Figure .3 Ammonium-N in soil in 1-, 2-, 3- and 4-year-old leys at Tulloch between January 1999 and August 2001. Columns represent means of 8 samples (1-, 2- and 3-year-old leys) and 4 samples (4-year-old leys). Bars represent standard errors.
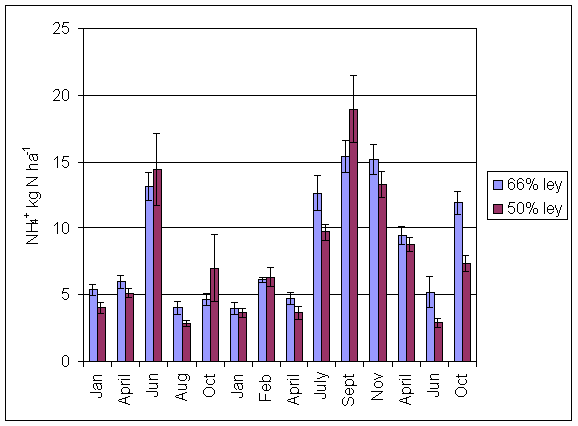
Figure .4 Ammonium-N in the 50% and 66% ley rotations at Tulloch (1–3-year-old leys only). Columns represent means of values for 12 plots. Bars represent standard errors.
SON varied significantly between monthly samples (REML p<0.001, Figure 5.4.5), but there was no obvious seasonal pattern. SON was higher in 1999 and 2001 than in 2000 (REML p<0.01, Figure 5.4.5), although the lack of an obvious seasonal pattern makes the data hard to interpret. There was no direct effect of vegetation type on SON, but on some sampling dates, SON was higher under clover, especially early in the growing season. SON under grass patches was more variable than SON under clover (REML p<0.01, Figure 5.4.5). SON was not significantly different in different ages of ley, different rotations, or different management regimes.
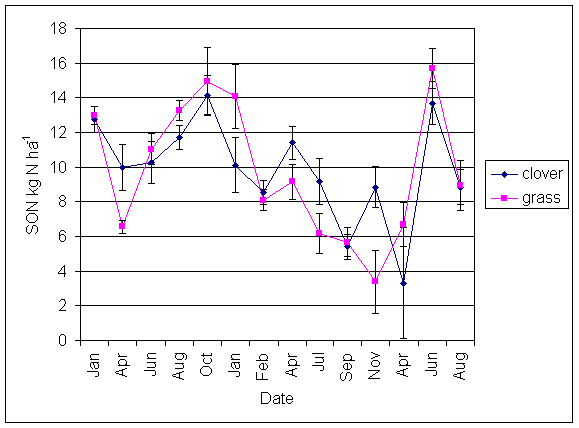
Figure .5 Soluble organic nitrogen under grass and clover microsites in all ley plots at Tulloch, between February 1999 and August 2001. Points represent means of 14 samples. Bars represent standard errors.
TSN was higher in 2000 than in other years, but the difference between years was not very large (REML p<0.001, Figure 5.4.6). TSN was up to 50% higher in 3 and 4-year-old leys than in 1 and 2-year-old leys (REML p<0.001, Figure 5.4.7). TSN was lowest in April (approximately 20 kg N ha-1) and highest in the autumn (approximately 35-40 kg N ha-1 p=0.001, Figure 5.4.6). This seasonal pattern varied between years, and TSN was unusually low in August 1999 (p=0.001, Figure 5.4.6 and Figure 5.4.7). TSN was not significantly affected directly by management, rotation or vegetation type. During the summer TSN was up to 30% higher in clover microsites than in grass microsites (p=0.001, Figure 5.4.6).
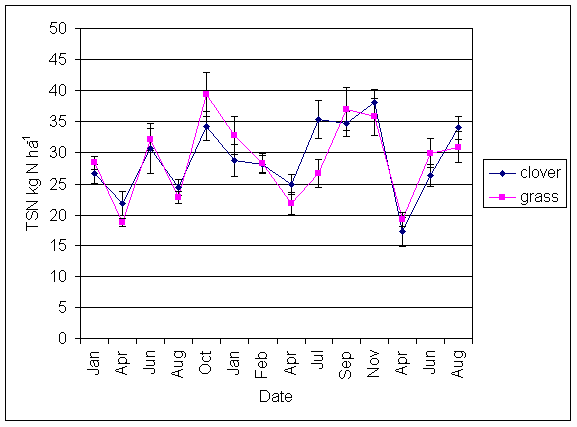
Figure .6 Total Soluble N under grass and clover microsites in all ley plots at Tulloch, between February 1999 and August 2001. Points represent means of 14 samples. Bars represent standard errors.
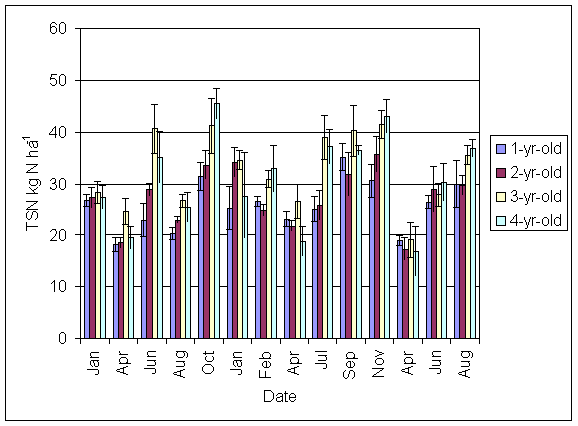
Figure .7 Total Soluble N in soil in 1-, 2-, 3- and 4-year-old leys at Tulloch between January 1999 and August 2001. Columns represent means of 8 samples (1-, 2- and 3-year-old leys) and 4 samples (4-year-old leys). Bars represent standard errors.
Grass and clover microsites had similar levels of extractable P and Mg in August 2001. P and Mg were also similar in cut and grazed leys and did not vary significantly between different ages of ley or different rotations. K levels were significantly higher in grass microsites than clover microsites in August 2001 (REML p <0.001), especially in 2 and 3-year-old leys (REML p <0.01, Figure 5.4.8). K levels were similar in cut and grazed leys and did not vary significantly between different ages of ley or different rotations.
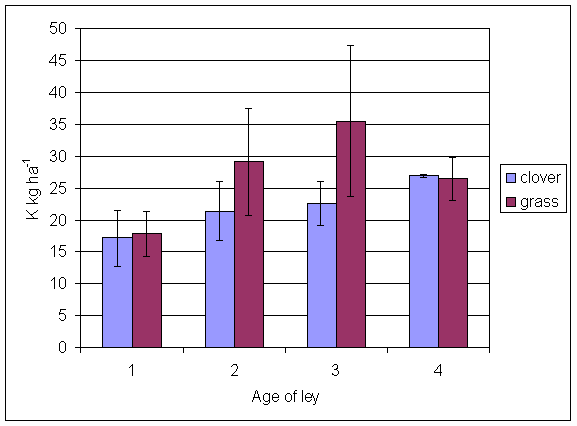
Figure .8 Potassium levels in soil from under grass and clover microsites in 1-, 2-, 3- and 4-year-old leys in August 2001. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors.
This part of the study tested the following hypotheses:
Nitrate-N was highest in 3 and 4-year-old leys, and lowest in 2-year-old leys. In 1-year-old leys, nitrate-N was also high on some sampling dates. Nitrate did not therefore show a steady increase with age of ley, although it was clearly higher in older leys (Figure 5.4.2).
Ammonium-N levels on almost all sampling dates were highest in 4-year-old leys and lowest in 1-year-old leys. Ammonium-N was clearly increasing with age of ley (Figure 5.4.3). Soluble organic N showed no clear pattern with age of ley. Total Soluble N was highest in 3-year-old leys (Figure 5.4.7).
Overall, soil N was accumulating over the lifetime of the ley, although the low soil nitrate-N in 2-year-old leys does not fit this pattern. Benefits to the soil from a fourth year of ley were not obvious. Four-year-old leys often had lower soil N than 3-year-old leys, possibly because the high grass yield in the 4-year-old leys removed large amounts of N from the system, so an extra year of ley may not greatly improve soil fertility. It is also possible that denitrification and leaching removed large amounts of N from 4-year-old leys. This confirms the findings of Johnston et al. (1994), who studied the effects of 1–6-year ryegrass-clover leys on soil fertility, and concluded that increasing the length of the ley phase beyond three years did not significantly increase the availability of nitrogen or subsequent crop yields. Granstedt (1992) also found that when clover content was allowed to fall below 30-50% of ley biomass, as it did in the 4-year-old leys in 2000, then offtake of N in grass is likely to exceed N input by N fixation.
Other data from the site indicates that P, K and Mg showed no consistent pattern with respect to age of ley, when data from different years was examined (Figure 5.5.2, Figure 5.5.3, Figure 5.5.4). pH tended to decline with increasing age of ley (Figure 5.5.5). It was difficult to distinguish the effects of ley age from the effects of management: all 1-year-old leys were grazed and all 2 and 4-year-old leys were cut. Cut leys had composted farmyard manure spread on them in the early spring, and this would have affected their nutrient levels. However, there was no evidence of elevated N, P, K or Mg levels in the cut leys. Nitrate-N was significantly lower in cut leys than in grazed leys in early summer, and no other significant differences between the soil in cut and grazed leys were observed.
There was little difference between the two rotations in terms of their levels of nitrate-N, SON or TSN, but on most sampling dates, there was higher ammonium-N in leys of the 66% ley rotation than in the 50% ley rotation. This was true in all ages of ley, and was not affected by any seasonal trends. There was no significant effect of cutting or grazing on ammonium-N, so it is likely that this represents a long-term effect of the rotations on soil fertility. The 66 % ley rotation had only two years of arable crops in each cycle, and only one year without clover. On the other hand, removal of N in grass in the 4-year-old leys greatly exceeded N fixation in 2000. If the values of N fixation and grass N yield observed in 4-year-old leys were typical of previous years, then the 66% ley rotation might ultimately be less fertile than the 50% ley rotation.
Nitrate-N was higher under clover patches than under grass patches over all sampling dates, and especially in summer, confirming hypothesis c. Total soluble N was also higher under clover patches in summer, and SON was also significantly higher under clover patches than under grass patches on some sampling dates. There was no difference between different ages of ley in this respect.
Differences were also observed between grass and clover microsites in the levels of K in summer. It is possible that K becomes depleted underneath clover patches, over time while nitrogen is elevated. Faerge and Magid (2003) suggest that the supply of potassium is a key limiting factor for growth and N fixation by organic white clover leys, although this was based on studies of farms in Denmark, so it may not be representative of the soil conditions in Scotland. Low clover content in grass-clover swards has been linked to potassium deficiency (Evans et al., 1986). All this suggests that white clover has a higher demand for potassium than grass. The alternative explanation for this data is that clover plants may be more likely to colonise microsites that are high in nitrate-N and low in potassium. This latter hypothesis seems unlikely, as the differences between clover and grass microsites seem to increase over time.
Differences in pH, calcium and phosphate have also been measured between clover and non-clover microsites in a number of upland sites in North Wales. Not all of these differences were consistent across all sites. The strongest correlation seemed to be between clover and calcium, but this may have been due to the positive relationship between calcium and pH (Snaydon, 1961). It is not possible to distinguish cause and effect in most of these cases: nutrient levels or pH may be high or low because of clover, or clover may be growing in a particular place because of the soil conditions. In some cases though it is likely that clover is affecting the soil: Matthew et al. (1995) observed that clover microsites had greater soil aeration, higher grass N concentration, and greater herbage mass than microsites without clover. In this case, the most likely explanation is that clover is elevating soil N in its immediate vicinity. The rate of mineralisation of N is greater under clover, grass mixtures than under grass monocultures, probably because of the low C:N ratio of clover residues (Elgersma and Hassink, 1997). This increased mineralisation of soil organic N, by clover may be a more important source of nitrogen for grass than directly transferred fixed N, especially in soils rich in organic matter (Mengel, 1996), like the soil at Tulloch. Mineralisation of soil organic matter by clover may explain the discrepancies between N transfer as calculated by the isotope methods, and the N difference methods (Section 3.6.8). Taken together this evidence suggests that clover grows preferentially in microsites with particular soil conditions, and that it also modifies the soil conditions locally, for example by increasing the availability of soil N.
Nitrate under grass patches was higher than nitrate under clover patches on some sampling dates in winter, but the differences were not significant. If cyclical replacement occurred over a single growing season, clover would invade low N microsites early in the growing season, and elevate the soil N levels over the summer, before being replaced by grass late in the season, and retreating to a few low N microsites. There were considerable variations between samples gathered on the same date, especially in grazed leys, probably as a result of dung and urine patches, and this may have masked changes in soil N caused by the clover. This method only sampled soil from randomly chosen clover patches, it did not measure changes in soil N under individual clover patches, or closely examine changes in the distribution of clover patches in the field. The invasion experiment (Section 5) examines changes in the distribution of clover under experimental conditions, and addresses some of these questions.
This section of the study has measured the availability of soluble forms of nitrogen, in order to compare soil fertility in different ages of ley, year, season, rotation etc. It should be emphasised that these measurements of soil N do not indicate the total amount of nitrogen in soil, rather, they measure how much of the soil nitrogen is in soluble forms. This gives an estimate of the availability of nitrogen to grass and clover. Ammonium and nitrate can be used readily by grass and clover, but the extent to which grass and clover can use SON is not known. SON comprised between 15 and 50% of the total soluble soil N under leys at Tulloch. Measurements of soluble soil N are not always predictably related to total soil N (Beauchamp et al., 2003), so the changes in soluble N measured in these experiments may not accurately represent the N balance of the system. In addition to the soluble forms of N, soils contain N within the soil organic matter pool. This pool can be categorised as:
(Bjarnason, 1988).
Soil organic matter in leys at Tulloch comprised 8-12% of soil dry matter. There were differences between ages of ley in some years, but there was no clear overall pattern (Figure 5.5.1).
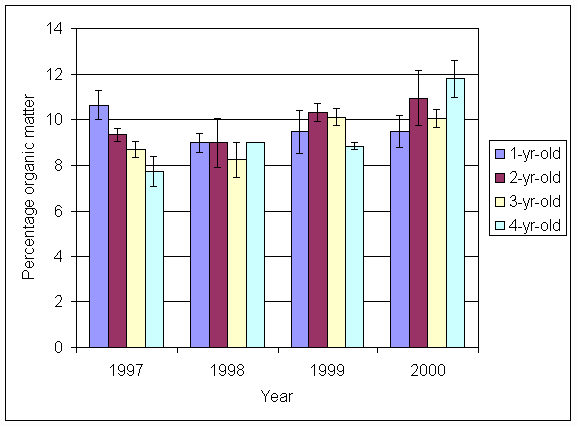
Figure .1 Percentage organic matter in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
Annual variation of soil nutrientsNitrate-N and ammonium were both lower in 1999 than in 2000 and 2001, although SON and TSN did not follow this pattern. TSN was significantly higher in 2000 than in other years. SON was not characterised in this experiment, so it is not possible to state how much of this dissolved N was available to grass and clover. Little is known about the ability of clover to utilise organic N (Caradus, 1990). As ammonium was highly variable, and some of this variation may have been due to daily variations in temperature, rainfall etc., nitrate-N probably provides the clearest picture of the levels of soil N available to clover. Other data from the site indicates that over the period of study, soil P levels were highest in 1999 and lowest in 1997 and 2000 (Figure 5.5.2). K, Mg and pH did not change significantly between 1997 and 2000 (Figure 5.5.4 and Figure 5.5.5).
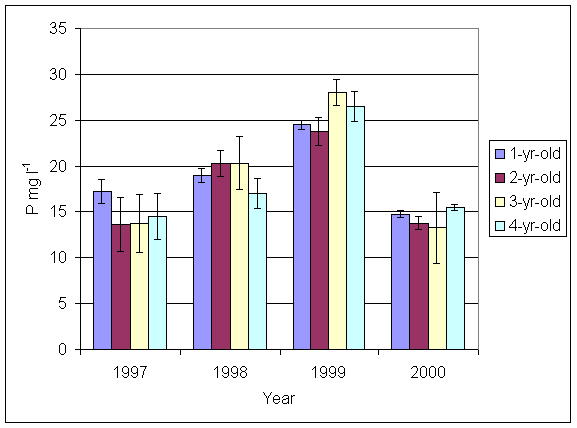
Figure .2 Phosphorus levels in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
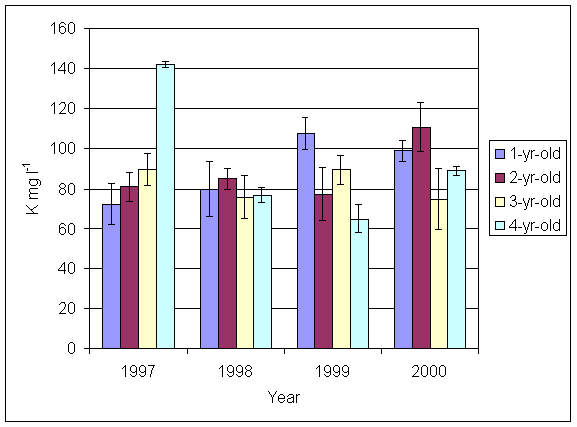
Figure .3 Potassium levels in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
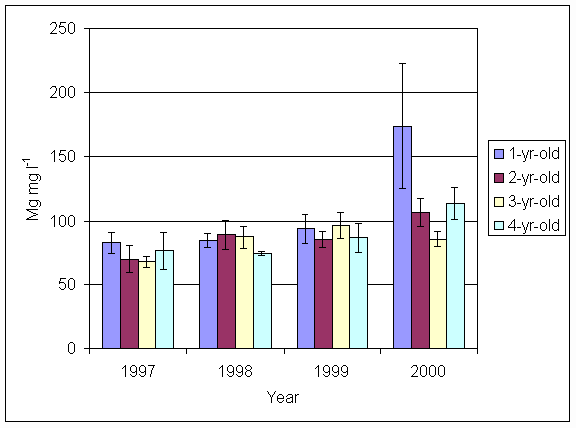
Figure .4 Magnesium levels in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
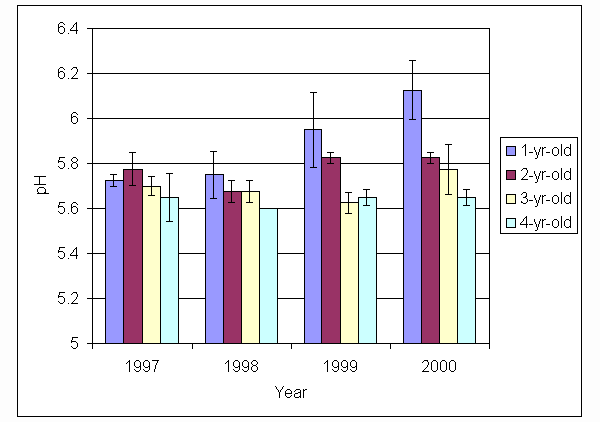
Figure .5 pH levels in soil from 1-, 2-, 3- and 4-year-old leys at Tulloch. Columns represent means of 4 samples (1-, 2- and 3-year-old leys) and 2 samples (4-year-old leys). Bars represent standard errors (SAC data).
Nitrate-N, ammonium-N and TSN were all highest in autumn and winter. This is probably because of uptake of soil N by actively growing grass and clover during the growing season. Elevated soil mineral N in autumn under ryegrass-clover set aside swards has been observed by Chalmers et al. (2001), in the absence of any manure or fertiliser applications, strongly suggesting that some of the elevated mineral N results from rhizodeposition by clover. Eriksen et al. (1999) have also observed elevated nitrate under grass-clover leys in winter. As in this study, nitrate levels were higher in winter in the 2-year-old leys than in the 1-year-old leys. SON did not follow this pattern, although there were large differences between SON levels on different sampling dates. SON was high between August and December 1999 and low between August 2000 and April 2001. It is not clear what caused these variations, it may have been due to variation in the rate of production of SON by plant roots, uptake of SON by roots, or mineralisation of SON by microbes. There were no obvious relationships between SON and rainfall or temperature.
Nitrate-N did not change significantly under different management systems when samples from all months were considered, probably because cutting of leys only took place during the growing season. However, nitrate-N was significantly affected by a combination of management and season: during the summer months, nitrate-N was higher under grazed leys. Over winter there was little practical difference between management of "cut" and "grazed" leys, so it is fair to conclude that grazing by sheep elevated slightly the level of soil nitrate-N in this system, possibly by the transfer of fixed N to the soil in dung and urine. Ammonium, SON and TSN did not change significantly under different management systems in this system.
Nitrate-N was approximately twice as high in April 2000 as in April 1999. TSN was also lower in 1999 than 2000. pNdfa was approximately 5% lower in 2000 than in 1999, which could be taken as evidence that pNdfa was being weakly suppressed by high soil N. Variations in pNdfa between leys of different ages and years only became apparent late in the growing season, whereas variations in soil N were detectable throughout the growing seasons. Therefore, if suppression of pNdfa by elevated soil N occurs at this site, it must be a delayed effect. Danso et al. (1988) applied 80 kg N ha-1 to a white clover-fescue pasture in two applications over a five-month period, and saw no effect on the pNdfa until the final sample, when pNdfa in the high N treatment was 5.3% lower, showing that these results are not completely unprecedented.
The reduced pNdfa in 2000 did not result in reduced N fixation. The reason for this was that the N concentration of clover increased substantially (c.30%) between 1999 and 2000, while clover yield was generally similar in both years, except in 2-year-old leys. The increase in clover N concentration between 1999 and 2000 could not have been simply due to increased uptake of soil N, as this would have suppressed pNdfa far more than was observed. If available soil nitrogen reduces pNdfa and increases clover N concentration, some other factor must be influencing the dry matter yield of clover, and consequently the N fixation. From the evidence in the literature, applications of N do not seem to increase N concentration of clover, and do not always increase the N concentration of grass: Danso et al. (1988) applied 80 kg N ha-1 of urea to a white clover-fescue pasture, and saw no effect on the N concentration of the fescue or the clover. Herrmann et al. (2001) applied 80 and 160 kg N ha-1 to white clover leys, and found similar N concentrations in clover, in both treatments, and in grass in both treatments. Boller and Nosberger (1987) added 120-150 kg N ha-1 to white clover perennial ryegrass leys and observed no change in N concentration of either clover or grass. In contrast, Ledgard et al. (2001) observed an increase in grass N concentration from 3.2% to 3.8% when N fertiliser was increased from 0–400kg N ha-1. McAuliffe et al. (1958) observed an increase in clover N concentration from 3.83 to 4.52% when 90 kg N ha-1 was applied, and Vinther (1998) observed that urine application caused grass N concentration to increase from 2.6-3.1 to 4.0-4.8%, while clover N concentration increased slightly from 4.2-4.4 to 4.5-4.9 with the addition of urine.
In our experiment, grass N concentration was around 50% higher in 2000 than in 1999, and this was probably due to the higher soil mineral N levels in 2000. Pearson correlations were obtained for the Tulloch data from 1999 and 2000. Model terms were soil nitrate-N in April, soil ammonium-N in April, SON in April, TSN in April, total annual N fixation, average annual pNdfa, average annual %NC, total annual clover dry matter yield, total annual grass dry matter yield, average annual grass N concentration and total annual SDN. Soil nitrate-N at Tulloch in April was positively correlated with grass dry matter (Pearson 0.663, p<0.001), N concentration in grass (Pearson 0.66, p<0.001), N concentration in clover (Pearson 0.646, p<0.001), and SDN (Pearson 0.72, p<0.001); and negatively correlated with pNdfa (Pearson -0.526, p<0.01). TSN at Tulloch in April was positively correlated with grass dry matter (Pearson 0.503, p<0.01), N concentration of grass (Pearson 0.466, p<0.05), N concentration in clover (Pearson 0.461, p<0.05) and SDN (Pearson 0.72, p<0.001); and was negatively correlated with pNdfa (Pearson -0.557, p<0.005, Figure 5.6.1). SON at Tulloch in April was also correlated with N concentration of grass (Pearson 0.397, p<0.05).
Grass yield was generally higher in 2000 than in 1999, except in 2-year-old leys. Soil derived N was approximately twice as high in 2000 as in 1999. Annual variation of soil derived N therefore appears to be closely related to annual variation of available soil N. To a lesser extent, the same seems to be true of grass N concentration and grass yield in this system. These conclusions are tentative, as comparisons of soil N and plant behaviour could only be made over 2 years.
Other data from the site indicated that soil phosphate levels were almost twice as high in 1999 as in 2000 (Figure 5.5.2), but this doesn’t seem to have affected N fixation. K, Mg and pH were similar in both years (Figure 5.4.8, Figure 5.5.4 and Figure 5.5.5).
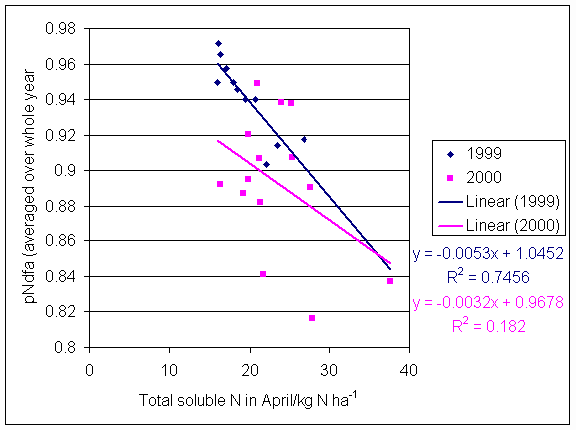
Figure .1 Proportion of nitrogen derived from the atmosphere (average over whole year, mean of three subplots) and total soluble nitrogen (values for April) for 1999 and 2000. Each point represents one plot
High soil N in any particular year would be expected to increase grass yield and grass N concentration and decrease pNdfa (and consequently elevate the soil derived N). The high grass yield would be expected to suppress clover yield. Therefore years with high grass yield, and grass N concentration should also have low pNdfa and low clover yield, if this annual variation is due to variations in soil N. Clover yield did not appear to follow this pattern. The highest grass yields and grass N concentrations, in 1997 and 2000, coincided with high clover yields. The lowest grass yields and grass N concentrations were observed in 1997 when clover yield was also high. However, pNdfa did seem to behave roughly as predicted, low in 1997 and 2000, but high in 1998 and 1999. Clover N concentration seemed to follow the same pattern of annual variation as grass N concentration, but other studies have not always found this to be the case: Carranca et al. (1999) observed subterranean clover (Trifolium subterraneum) and grasses over two successive growing seasons. Grass N concentration increased significantly between 1992/3 and 1993/4 but clover N concentration did not. Over the same period, grass dry matter yield declined and clover dry matter yield increased significantly. Nitrogen fixation was similar in both seasons, although pNdfa, measured by isotope dilution was higher in 1993/4. Soil N was not measured in both years, so it is not possible to say which if any of these changes were attributable to soil N.
N concentration of all parts of white clover plants may be increased by P and K deficiency, even if growth and morphology of the clover plant is not affected (Hogh-Jensen et al., 2001). In contrast, Bailey and Laidlaw (1999) observed that P deficiency caused clover plants to maintain leaves and petioles at the expense of stolons. K deficiency had less effect, but encouraged the development of stolons at the expense of leaves and petioles (Bailey and Laidlaw, 1999). The high soil K in 1999 could therefore explain the low N concentration of clover in that year. Overall, K was not however, related to the N concentration of clover, but P was negatively correlated with clover N concentration (Table 5.6.1), confirming the findings of Hogh-Jensen et al. (2001).
The age related patterns of soil nitrate-N were reflected in the soil derived N values and also in the N concentration of clover and grass. This age-related pattern of soil nitrate-N also varied between 1999 and 2000 in a similar way to the age-related patterns of soil derived N, grass N concentration, grass yield and clover N concentration. In 1999 these factors were all relatively high in 3-year-old leys and low in 1- and 2-year-old leys; in 2000 these factors were low in 2-year-old leys and high in 1 and 3- and 4-year-old leys (Figures 3.5.16, 3.5.17, 3.5.24, 3.5.25, 3.5.29, 3.5.30, 3.5.33,
3.5.34 and Figure 5.4.2). Dry matter of clover and N fixation did not closely follow this pattern. It is fair to conclude that both age-related variation and annual variation of clover N concentration, grass N concentration, pNdfa and soil derived N, are largely caused by variations in available soil N. Annual and age related variation of clover yield (and therefore N fixation) cannot be linked directly to soil N. The age related variations in plant behaviour and soil N can be tentatively explained as follows:In the first year of the ley, soil nitrate-N is moderately low and pNdfa is high. Ground cover and yield of clover are determined more by the sowing density, than by any ecological factors.
The following year, soil N falls, presumably because removal of N by leaching and grazing in the first year exceeded the input of fixed N into the soil. Grass shows signs of N deficiency, and consequently clover spreads at the expense of grass to dominate much of the pasture. The pNdfa falls slightly, probably because of the reduced competition from grass for soil N.
The following year, soil N increases as a result of above-ground and below-ground transfer, and decay of clover over winter. Some of the additional soil N is taken up by clover, causing a further decline in pNdfa. N concentration of clover increases, although the exact cause of this is not clear. Grass is unable to rapidly re-colonise microsites high in soil N as many of these are physically occupied by clover, and grass cannot spread as quickly as clover, so grass yield does not increase in line with the increase in available soil nitrate-N. Grass N concentration increases, because of the increased availability of soil N.
The following year, grass spreads and colonises microsites high in soil N, suppressing clover growth and N fixation. Grass yield increases, availability of soil N decreases and pNdfa rises. This is in general agreement with the idea of a two-year cycle in the effect of soil N on clover and grass, resulting from a one-year time-lag in the effects of grass and clover on each other, as proposed by Loisseau et al. (2001), based on experiments with clover and grass, grown in sandy soil in central France.
The situation where pNdfa is low, soil nitrogen is high, grass yield is low and grass and clover N concentrations are high is inherently unstable, as it indicates that clover is growing under conditions which favour grass. When this is observed in any particular year and/or age of ley, it indicates that grass is soon to increase at the expense of clover.
Soil derived N in clover comprised between 2 and 10% of total soil derived N. This figure was highest in 2 and 3-year-old leys. This suggests that under certain circumstances (for instance when clover content of the sward is high), clover can compete for soil N to a limited extent. This could also explain why the age related patterns of grass yield, clover yield and consequently N fixation are only approximately related to age related patterns of soil nitrate-N. Low nitrate-N in 2-year-old leys causes clover yield to increase, as clover rapidly spreads, and colonises new areas of the sward under these conditions and grass growth is suppressed. Soil pH also affected clover yield in this experiment (Table 5.6.1), and was probably at least partly responsible for the differences in clover yield between different ages of ley. Soil pH declined with age of ley in 2000, from 6.1 in 1-year-old leys to 5.65 in 4-year-old leys, and the equivalent clover dry matter yields were 1909.4 and 985.9 kg ha-1, respectively. This is in close agreement with the findings of Bailey and Laidlaw (1999), that increasing soil pH from 5.4 to 6.1, doubled the dry matter yield of clover. The mechanism for this seemed to be that a rise in pH increased the availability of P to the clover (Bailey and Laidlaw, 1999). This evidence came from pot experiments using clover monocultures. In the field, with competition from grass, high P and K may have different effects. These changes in pH may be long term effects of clover on soil (Raven and Smith, 1976, Tang, C., 1998, Monaghan et al. 1998).
Seasonal variation of N fixation did not seem to be directly related to seasonal changes in soil nitrate-N. Seasonal variation of dry matter of clover (and consequently N fixation), seemed to be more strongly affected by seasonal variation of ground temperature than seasonal changes of soil N. This is shown by the fact that the seasonal pattern of clover yield varied between years, but not between ages of ley. This may be in part due to a timelag in the effect of soil N on clover, as described earlier. Seasonal variation of grass yield was also not obviously related to seasonal variation of soil nitrate-N, although the seasonal pattern varied between different ages of ley in the same year, so it cannot have been entirely due to climatic variation. In grass-clover swards, additions of mineral N generally increase grass yield and leaf area index and reduce pNdfa, but the effect of 100 kg N ha-1 (Laidlaw and Withers, 1998) or even 400 kg N ha-1 (Hogh-Jensen and Schjoerring, 1994) may only become apparent late in the growing season.
In this study, the decline in pNdfa over the growing season seemed to mirror the increase in soil nitrate-N over the same period, although differences between soil N in leys of different ages were more obvious early in the season than differences in pNdfa. The increase in soil nitrate-N over the growing season was most marked in 3-year-old leys, and the seasonal decline in pNdfa was also steepest in 3-year-old leys. Soil nitrate-N rose more sharply over the growing season in 2000 than in 1999, and pNdfa fell more sharply in 2000 than in 1999 (Figures 3.5.12 and 3.5.13). Similar inversely related seasonal patterns of white clover pNdfa and soil N have been observed in New Zealand (Ledgard et al., 1987).
Clover N concentration followed a roughly similar seasonal pattern to soil nitrate-N. This may have been partly due to seasonal changes in the proportions of leaves, petioles, stolons and flowerheads in the samples. In summer, white clover stolons typically have a nitrogen concentration of 2-2.75%, petioles 2.5-3.25% and laminae 4-5% (Hogh-Jensen et al., 2001) However, in this study, the initial sample and the last sample both consisted almost entirely of leaves and petioles, and yet the final sample had a higher N concentration than the initial sample. This strongly suggests that there are also changes occurring in the N concentration of the plant tissues. The relationship between seasonal changes in clover N concentration and seasonal changes in soil nitrate-N (if there is one) is not obvious.
Seasonal variation of grass N concentration followed a similar pattern to soil nitrate-N, generally rising over the course of the growing season. The seasonal pattern varied between years and in different ages of ley, but these patterns weren’t obviously related to the seasonal patterns of soil nitrate-N in different years and ages of ley.
Soil derived N varied little over the course of the growing season in 1999 and 2000, except in the 4-year-old leys. Seasonal changes in soil derived N could not be clearly related to soil N and were probably due in a large part to the effect of the weather on uptake of N by grass, although there were differences in the seasonal pattern between ages of ley, sampled over the same growing season. Seasonal variation of grass yield was also not obviously related to seasonal variation of soil nitrate-N, although as with soil derived N, the seasonal pattern varied between different ages of ley, so it cannot have been entirely due to the weather either.
The slightly higher ammonium levels in the 66% ley rotation could explain the slightly reduced pNdfa, clover yield and N fixation, as well as the elevated soil derived N, grass yield and grass N concentration in 1-year-old leys in the 66% rotation. Rotation effects would become more pronounced with time, accounting for the year/rotation interactions. The two rotations at Tulloch were relatively similar, each having a cycle of 6 years, with 2 years of cereals, and a sequence of cut and grazed leys. Greater differences might have been observed if a ley-arable rotation was compared to a stockless rotation. A comparison between a rotation with 3 years of cut leys and one with three years of grazed leys might have shown clearer differences between these two types of management.
Pearson correlations were obtained for the Tulloch data from 1997-2000. Variables examined were year, pH, K, P, Mg, soil organic matter (som), plot number, ley age, grass dry matter yield (dmg), clover dry matter yield (dmc), N concentration of grass (%Ng), N concentration of clover (%Nc), pNdfa and N fixation (Nfix) (Table 5.6.1). N fixation was positively correlated with pH, Mg, SOM and clover yield. pNdfa was negatively correlated with DMG, %NG (and hence grass N yield, Figure 5.6.2, Table 5.6.1), %NC (Figure 5.6.3) and SDN, but not related to pH, K, P, Mg or soil organic matter. DMG, %NG, %NC and SDN were all positively related to nitrate-N, so it is likely that these correlations represent the response of pNdfa to soil mineral N.
Riffkin et al. (1999) surveyed 71 dairy pastures in South West Victoria, Australia, and found that pNdfa could be predicted from soil type (medium or light), rhizobial numbers, total soil N, soil K, soil P, crude protein level in ryegrass and numbers of the nematode pratylenchus. Crude protein level in plants is related to tissue N concentration (Allison, 1973), so there is some agreement with this study. Riffkin et al. (1999) also found that N fixation could be predicted from soil type, density of Heterodera and Ditylenchus nematodes, soil pH, applied P, soil P, soil K, TSN, crude protein levels in white clover and ryegrass and digestible dry matter of ryegrass. Surprisingly, use of N fertiliser (up to 121 kg ha-1) did not have any discernible effect on pNdfa or N fixation (Riffkin et al., 1999).
At Tulloch, Mg was positively correlated with pH, both P and Mg were higher at the lower end of the site (block 1). Clover yield was positively correlated with P and pH. N concentration of grass was negatively correlated with P, and it was also positively correlated with grass yield. N concentration of clover was negatively correlated with P and clover yield, and positively correlated with grass yield and grass N concentration. N fixation was positively correlated with pH, Mg, SOM and clover yield. SDN was negatively correlated with P, and positively correlated with grass yield, N concentration of grass, N concentration of clover and N fixation.
It is difficult to identify causal relationships in all these cases, but it appears as though high soil P causes clover to grow at the expense of grass, reducing both grass yield and N concentration. As grass N yield is the main component of SDN, this is also reduced. P also reduces N concentration in clover, with the effect that the increased clover yield doesn’t result in higher N fixation. High N concentration in clover may be a sign of plant stress rather than health. High soil N reduces pNdfa and increases grass yield, grass N concentration, SDN and clover N concentration, and this explains the correlations between these factors. N fixation is enhanced by a relatively high soil pH, and this also increases the availability of Mg. It is not clear from this study whether SOM increases N fixation, N fixation increases SOM, or both, or if both N fixation and SOM are affected by a third factor. Soil nutrients, such as P and K and Mg may be limiting factors for clover growth and N fixation at some sites but not at others. At Tulloch, variations in the level of soil P affected some aspects of clover growth but did not have a significant effect on N fixation overall.
|
year |
pH |
K |
P |
Mg |
som |
plot No. |
ley age |
dmg |
dmc |
%Ng |
%Nc |
pNdfa |
N fix |
|
|
pH |
ns |
|||||||||||||
|
K |
ns |
ns |
||||||||||||
|
P |
- ns |
- ns |
ns |
|||||||||||
|
Mg |
+ * |
+ *** |
ns |
ns |
||||||||||
|
som |
ns |
ns |
ns |
ns |
ns |
|||||||||
|
plot No. |
ns |
ns |
ns |
- ** |
ns |
ns |
||||||||
|
ley age |
+ ** |
- * |
ns |
ns |
ns |
ns |
ns |
|||||||
|
dmg |
ns |
ns |
ns |
ns |
ns |
ns |
ns |
+ * |
||||||
|
dmc |
+ ** |
+ * |
ns |
+ * |
ns |
ns |
ns |
ns |
ns |
|||||
|
%Ng |
ns |
ns |
ns |
- ** |
ns |
ns |
ns |
ns |
+ *** |
ns |
||||
|
%Nc |
ns |
ns |
ns |
- ** |
ns |
ns |
ns |
ns |
+ ** |
- ** |
+ *** |
|||
|
pNdfa |
ns |
ns |
ns |
ns |
ns |
ns |
ns |
ns |
- *** |
ns |
- *** |
- *** |
||
|
Nfix |
+ *** |
+ ** |
ns |
ns |
+ ** |
ns |
ns |
ns |
ns |
+ *** |
ns |
ns |
ns |
|
|
sdn |
+ * |
ns |
ns |
- ** |
ns |
ns |
ns |
ns |
+ *** |
ns |
+ *** |
+ *** |
- *** |
+ * |
Table .1Correlations between soil and plant factors at Tulloch 1997-2000 (ns= not significant, * = p<0.05, ** = p<0.01, *** = p<0.001)
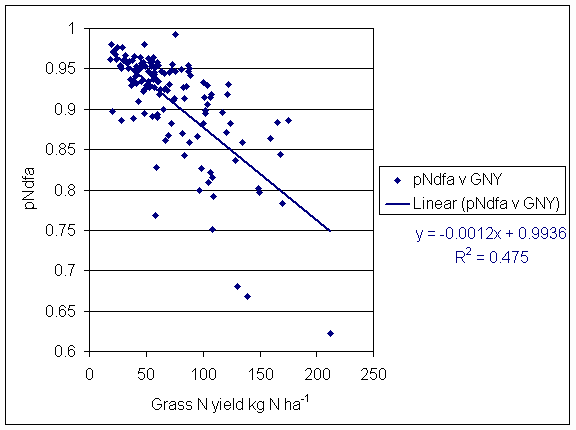
Figure .2 Correlation between proportion of clover N derived from the atmosphere (pNdfa) and grass N yield. Each point represents one subplot and sampling date.
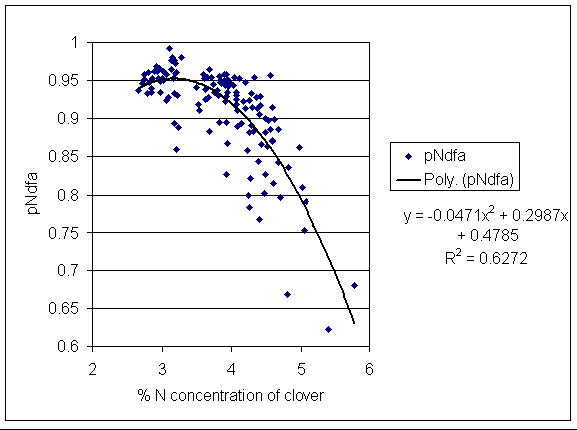
Figure .3 Correlation between N concentration of clover (%) and proportion of clover N derived from the atmosphere (pNdfa). Each point represents one subplot and sampling date.
There were strong positive relationships between soil N and the growth and chemical composition of grass and clover, and negative relationships between soil N and pNdfa. Soil N did not, however have a simple relationship with clover yield or N fixation. It is possible that both grass and clover N concentration rise with increasing soil N, but pNdfa was only weakly negatively correlated with clover N concentration (Figure 5.6.3). The increased clover N concentration observed in some years cannot simply be due to increased uptake of soil N, it may instead be a response to stress, or a change in the allocation of biomass to leaves, flowers, stolons and roots. Effects of management on N fixation could not be measured in this study, but nitrate-N was elevated by grazing during the summer months. Ground cover data from Tulloch indicated that the cut leys had more clover than grazed leys (Figure 4.5.22), confirming the findings of Schils et al. (1999).
Effects of clover and grass on soil nitrogen Introduction
This experiment was intended to measure transfer of nitrogen from clover to grass and to the soil, under experimental conditions. Although experiments similar to this one were performed in the 1930’s (Virtanen 1930, Virtanen et al. 1936), studies of this kind on white clover are remarkably scarce. Data on rhizodeposition and N transfer is often contradictory, with different experimental methods producing different results: Hatch and Murray (1994), and Murray and Hatch (1996) measured no transfer of fixed N from white clover to grass, using a 15N labelling method, unless the clover roots were damaged. McNeill and Wood (1990) labelled clover plants using 15N2 in a sealed chamber with ryegrass plants. No 15N was detected in the ryegrass plants after 129 days of growth, indicating that no fixed N was transferred from clover to grass under those conditions in the time of the experiment. Frey and Schuepp (1992) observed that mycorrhizae facilitated transfer of N between berseem (Trifolium alexandrinum L.) and maize. In contrast, Rogers et al. (2001), found that the presence of mycorrhizae did not affect N transfer between ryegrass and clover. Høgh-Jensen and Schjoerring (2001) used leaf labelling to measure rhizodeposition, and calculated that the amount of N deposited in soil by clover roots in mixed swards was almost 50% of the amount harvested in leaves and shoots. Field based 15N experiments usually produce lower estimates of N transfer than leaf labelling, generally between 0 and 35% of total fixed N (Laidlaw et al. 1996, Boller and Nosberger, 1987, Høgh-Jensen and Schjoerring, 1997, Høgh-Jensen and Schjoerring, 2000, Ledgard and Steele, 1992). The 15N dilution experiments in this study (Section 3.5.8) produced widely varying estimates of N transfer, that were mostly low or negative. In contrast, N difference calculations based on the same samples gave estimates similar to those in the literature.
Evidence from the field showed that soluble soil N levels were higher under clover patches than under grass patches, especially during the summer (Section 5.4). This does not necessarily mean that clover is elevating soil N by root and leaf turnover or root exudation. The soil sampling experiment only sampled random clover patches on each sampling date. It did not record changes in the distribution of the clover, or levels of total soil N under individual clover patches. A preliminary experiment measuring effects of clover and grass on soil N was performed in 2000. Details are given in the appendix.
This section of the study was intended to test the following hypotheses in the laboratory:
Perennial ryegrass (Condessa) and white clover (Avoca) seeds were sown in pots containing 300g of either acid washed sharp sand, or sieved soil from Tulloch (dry matter). Each pot received 3 seeds of one or both species. Clover seeds were innoculated by mixing the seeds with a mixture of wet peat and rhizobial innoculum, and allowing the mixture to dry before sowing.
Pots were made from 2 L carbonated drinks bottles; cut to form open topped plant pots 200 mm high. These pots were pierced at the bottom and the sides with a standard pattern of holes. The lower part of the pot was covered with tin foil, to prevent overheating of the soil. The pots were intended to contain the clover and grass plants in order to prevent leaves and seeds from dropping out of the pots. No N was applied to any of the treatments. Pots were placed in trays, out of doors, and watered from below with standard N free nutrient solution (Dart and Pate, 1959), consisting of 2.3 mM K2HPO4 l-1, 0.8 mM MgSO4.7H20 l-1, 5.8 mM CaSO4 l-1, 4.6 m M H3BO3 l-1, 9 m M MnSO4.H2O l-1, 1.0 m M CoSO4.7H2O l-1, 0.8 m M ZnSO4.7H20 l-1, 0.1 m M Na2MoO4.2H2O l-1, 0.3 m M CuSO4.H2O l-1 and 5 m M Fe l-1 as citrate. There were 7 treatments, each consisting of 8 replicates:
|
Treatment code |
Medium |
Plants |
|
1a |
Sieved soil |
Clover |
|
1b |
Sieved soil |
Clover + grass |
|
1c |
Sieved soil |
Grass |
|
1d |
Sieved soil |
None |
|
2a |
Sand |
Clover |
|
2b |
Sand |
Clover + grass |
|
2c |
Sand |
Grass |
Table .1 Experimental treatments in the pot experiment to test effects of clover and grass on soil
Soil and N-free sand treatments were included to control for the N sparing effect (Section 1.4.1): Treatments 2a, 2b and 2c contained no N beyond that added in the form of seeds.
Rhizodeposition from clover grown in sand could therefore be calculated as the difference between N in sand in treatments 2a/b and N in sand from 2c. N levels and grass yields from treatment 2c were expected to be very low, because the grass in this treatment would be reliant on seed reserves for N.
The soil treatments were intended to distinguish the effects of clover and grass on soil N. The observed elevation of soil N under clover patches in the field, could have resulted from rhizodeposition by clover or from lower uptake of N by clover than grass (the N sparing effect). Treatment 1d was included as a control, to measure changes in soluble soil N resulting from mineralisation of soil organic matter over the growing season. By comparing soluble N levels in 1d with treatments 1a, 1b and 1c, it should be possible to calculate net rhizodeposition of N by clover and uptake of N by grass.
Grass and clover were harvested at the end of the experiment, separated and weighed. Roots were removed from the sand or soil and the soil or sand was analysed for nitrate, ammonium and TSN as described in section 3.3.2. The experiment ran from 01.03.01 to 30.10.01
CalculationsNitrate-N, ammonium-N, SON and TSN values in ppm were multiplied by the following formula in order to convert the output of the autoanalyser (ppm per sample) into mg N per g dry soil:
![]() [.1]
[.1]
Dry matter yields of grass and clover are shown in Figure 6.4.1. Clover yielded better than grass in sand, but grass yielded better in soil. Grass grown with clover did not yield more than grass grown as a monoculture. Yields of grass grown in sand were extremely low. Total N in grass and clover had not been measured at the time of writing.
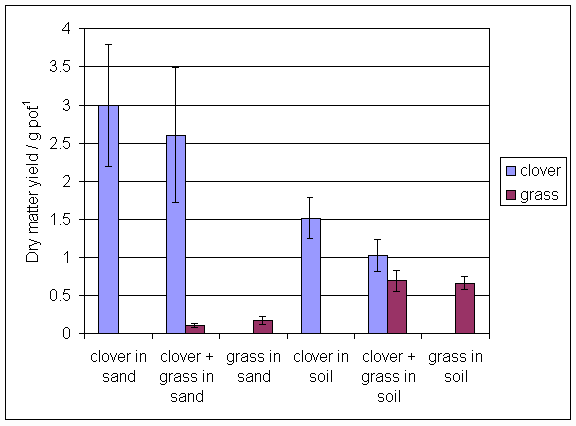
Figure .1 Dry matter yields of grass and clover in main root exudation experiment. Columns represent means of 8 replicates. Bars represent standard errors.
Nitrate-N levels were not significantly affected by the type of vegetation in the pots. Nitrate was also similar in soil and sand treatments
(Figure 6.4.2). Nitrate levels were considerably lower than those observed in the field: Nitrate-N levels in October 1999 and November 2000 in the top 150 mm of soil at Tulloch were on average 4.9 mg N/g dry soil-1.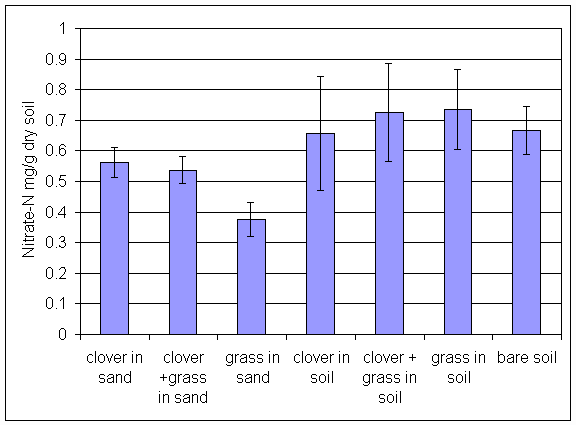
Figure .2 Nitrate-N levels in soil from all treatments in the main root exudation experiment. Columns indicate means of 8 replicates. Bars indicate standard errors.
Soil ammonium-N was significantly lower in sand pots than in soil pots (REML p<0.001), and was lower in pots containing grass than pots with clover or bare pots
(REML p=0.03). Ammonium-N levels are shown in Figure 6.4.3. Ammonium-N levels in the soil treatments were in a similar range to those observed in the autumn in the field at Tulloch: Ammonium- N levels in the top 150 mm of soil at Tulloch were on average 4.1 and 9.2 mg N (g dry soil)-1, in October 1999 and November 2000.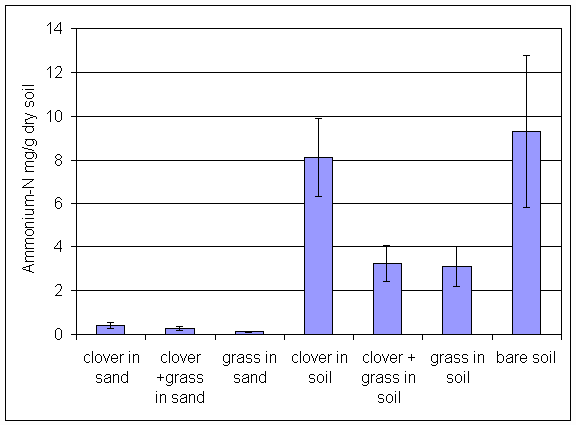
Figure .3 Ammonium-N levels in soil from all treatments in the main root exudation experiment. Columns indicate means of 8 replicates. Bars indicate standard errors.
TSN was higher in soil pots than sand pots (REML p<0.001)
, but TSN levels were not affected by the type of vegetation in the pots (Figure 6.4.5). TSN levels in the soil treatments were slightly higher than those observed in autumn in the field: TSN levels in the top 150 mm of soil at Tulloch were on average 22.7 mg N (g dry soil)-1, in October 1999 and November 2000.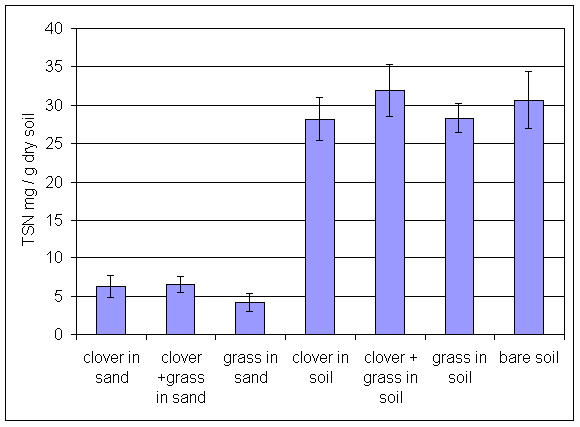
Figure .4 TSN levels in soil from all treatments in the main root exudation experiment. Columns indicate means of 8 replicates. Bars indicate standard errors.
SON was also higher in soil pots than sand pots (REML <0.001), but was not significantly affected by the type of vegetation in the pots (Figure 6.4.4). SON was considerably higher in the soil treatments than in the field in autumn: SON levels in the top 150 mm of soil at Tulloch were on average 9.0 and 3.7 mg N (g dry soil)-1, in October 1999 and November 2000.
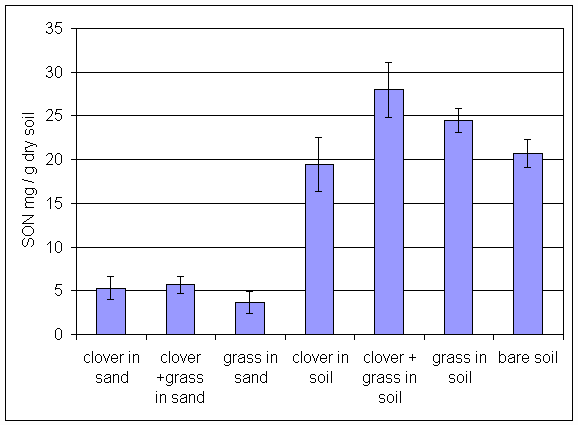
Figure .5 SON levels in soil from all treatments in the main root exudation experiment. Columns indicate means of 8 replicates. Bars indicate standard errors.
This experiment was intended to test the following hypotheses:
In the sand treatments, nitrate-N, ammonium-N, SON and TSN were all higher in treatments containing clover than those with grass only, but the differences were not significant. In the soil treatments, ammonium was significantly higher in clover monoculture treatments, than in treatments containing grass, but not significantly higher than the bare soil treatment. Nitrate-N, TSN and SON were not significantly higher in the presence of clover than in bare soil or grass monocultures. The soluble nitrogen levels did not closely resemble the conditions in the field, in particular, there was higher SON and lower nitrate-N in the pots than in the field.
Clover thrived growing in sand with 0N, but under these conditions ryegrass suffered and was extremely stunted, whether grown with or without clover. When grown in soil, grass did not seem to benefit from the presence of clover.
The presence of nitrogen in the grass-only sand treatments was surprising. It could be due to non-symbiotic N fixation by bacteria, as suggested by Virtanen et al. (1936), and it is possible that the use of N free nutrient solution encouraged the growth of free-living N fixing microrganisms (Liengen, 1999). A number of studies have found lower N transfer during the first year of clover growth than in subsequent years (e.g. Høgh-Jensen and Schjoerring (1997), Høgh-Jensen and Schjoerring (2000), Virtanen (1930), Jorgensen et al. (1999), Laidlaw et al. (1996), Elgersma and Hassink (1997)). It is possible that clearer evidence for N transfer might have been obtained if the experiment had been continued over a second growing season.
For these reasons, firm conclusions cannot be drawn from this experiment.
The soil chemistry experiment showed that there was higher nitrate and TSN under clover than grass in summer. The distribution of clover patches in the sward was not recorded, and the soil samples were not taken from precisely the same grass or clover patches in successive samples, so changes in the spatial pattern of clover in response to available soil nitrogen could not be observed.
It is known that the distribution of clover in a pasture changes relatively rapidly (e.g. Thorhallsdottir, 1990a,b) and that the amount of clover in the sward changes with the level of nitrogen in the soil. It is possible, therefore that the distribution of clover patches in a sward is determined at least partly by the distribution of soil N. Distribution of soil N is affected by grazing, dung and urine, and also by rhizodeposition of fixed N by clover. The effect of dung and urine on soil N, and soil heterogeneity is considerable: Afzal and Adams (1992) estimated that after 120 grazing days at a stocking density of 3.0 cows/ha, 19% of a pasture will be covered by dung and urine patches. As plants up to 15 mm from a dung patch can be affected, up to 50% of a pasture could be affected by dung and urine in any year in an intensively grazed pasture (Afzal and Adams, 1992, Vinther, 1998).
Rhizodeposition of fixed N by the clover may limit the time that clover can grow in a microsite before grass takes over, and the accumulation of rhizodeposited N triggers the movement of the clover plant to a new microsite. If this is the case, then confining a white clover plant to a single microsite for a year should result in an N enriched microsite, which is highly susceptible to invasion by grass. Likewise, a microsite containing only grass which has been repeatedly harvested should become depleted in N and susceptible to invasion by clover.
Containing clover and grass in the same area (experiments C and D) is intended to exaggerate the effects of the two species on the soil. According to the cyclical replacement hypothesis, clover is expected to elevate soil N while simultaneously depleting potassium, phosphate and other nutrients, up to a threshold point at which grass invades and dominates over the microsite. Grass is expected to deplete soil N, up to a threshold point at which clover invades and dominates the microsite. By preventing the invasion of clover and grass into new microsites, clover is expected to elevate soil N more than it would in the field, and grass is expected to deplete soil N more than it would in the field.
This part of the study tests the following hypotheses:
Methods
This section of the study consisted of four experiments. Experiment A was intended to observe the invasion of clover plants into microsites containing grass and bare soil, either fertilised with N equivalent to a urine patch or unfertilised. Experiment B was intended to measure invasion of perennial ryegrass into microsites containing clover, either fertilised with N equivalent to a urine patch or unfertilised. Experiment C was intended to observe the invasion of clover into microsites containing grass, either harvested or unharvested, after one growing season in which the clover and grass have been artificially contained in isolation. Experiment D was intended to observe the invasion of ryegrass into microsites containing clover, either harvested or unharvested, after one growing season in which the clover and grass have been artificially contained.
The experiment was performed out of doors in a wooden box 1.2m × 1.5m × 0.5m, open at the top and filled with sand. Drainage holes were made in the bottom of the box, and the top was left open. Each experiment consisted of three sections of plastic pipe 300 mm in diameter, 300 mm in length, buried in the sand, and filled with sieved Tulloch soil mixed with sand in a ratio of 1 part soil to 3 parts sand, to correct for the flush of soluble N resulting from drying, and re-wetting of the soil. Each pipe section was divided radially into 8 by stiff plastic dividers, with a circular central region 100 mm in diameter, thus creating 9 microsites of equal area (Figure 7.3.1). The central microsite was divided from the rest either by a cylinder of muslin coated with wax (Drew, 1975), to allow root penetration, but not diffusion of water or nutrients. To create the central microsite, rectangular pieces of muslin, 300 mm × 320 mm were dipped into molten wax and then allowed to cool. These were then rolled into tubes, and sealed along the edges with silicone sealant. Silicone sealant was used to attach the plastic dividers to the muslin cylinder and the inside edge of the pipe section, to create water-tight seals.
In experiments C and D, plastic sheet was placed around the muslin/wax membrane for the first year, to prevent any invasion in or out of the central region, during that time. The plastic sheet was made into cylinders of the same dimensions as the muslin wax cylinders, and sealed with silicone sealant.
Experiment A
Three white clover seedlings (Avoca) were transplanted into the central microsites of each pipe section. These central microsites were designated AC1. The remaining microsites in each pipe section were divided into two groups of 4 microsites: those on the north side and those on the south side of the pipe section (this was to control for the direction of sunlight). The 4 microsites in each group were designated at random to 4 treatments: AGI, AG2, AB1 and AB2. Treatment AG1 was planted with 3 perennial ryegrass seedlings (Condessa). Transplants received 100ml of distilled water. No N was applied. Treatment AG2 was planted with 3 perennial ryegrass seedlings and treated with 27.78 ml of artificial urine. The artificial urine was prepared from 14.22g urea, 1.00g allantoin, 2.88g hippuric acid, 0.35g creatinine, 5.77g glycine and 0.23 g ammonium chloride per litre of distilled water (Cuttle and Bourne, 1993, Doak, 1952). This mixture was equivalent to one sheep urination or 30 g N m-2 (Cuttle and Bourne, 1993, Doak, 1952). Treatment AB1 was left bare and received 100 ml of distilled water, but no N was applied. Treatment AB2 was left bare and treated with 27.78 ml of artificial urine. All treatments were cut on 6.9.00, 18.10.00, 24.5.01 and 13.8.01. Herbage from each microsite was separated into grass and clover, and the grass and clover fractions were dried and weighed. Numbers of flowerheads in the clover fraction were recorded. Any seedlings of weeds or grass that were not deliberately planted were pulled up and cut and mulched into the microsite in which they were found.
Experiment B
Three perennial ryegrass seedlings were transplanted into the central microsite of each pipe section. This microsite was designated as treatment BC1. The remaining microsites in each pipe section were divided into two groups of 4 microsites: those on the north side and those on the south side of the pipe section, as in experiment A. The 4 microsites in each group were designated at random to 4 treatments: BCI, BC2, BB1 and BB2.
Treatment BC1 was planted with 3 white clover seedlings. 100 ml of distilled water was applied to the seedlings after transplanting. No N was applied. Treatment BC2 was planted with 3 white clover seedlings and received 27.78 ml of artificial urine, as in experiment A. Treatment BB1 was left bare and received 100 ml of distilled water but no N. Treatment BB2 was left bare and received 27.78 ml of artificial urine.
All treatments were cut on 6.9.00, 18.10.00, 24.5.01 and 13.8.01. Herbage from each microsite was separated into grass and clover, and the grass and clover fractions were dried and weighed. Numbers of flowerheads in the clover fraction were recorded.
Any seedlings of weeds or grass that were not deliberately planted were pulled up and cut and mulched into the microsite in which they were found. Any clover stolons that grew over the plastic partitions between the peripheral microsites, were cut at the point at which they crossed the barrier, and then chopped and returned to the microsite in which they originated.
Experiment C
Three white clover seedlings were transplanted into the central microsite of each pipe section. No N was applied. The herbage was cut on 6.9.00, 18.10.00, 24.5.01 and 13.8.01, and separated into grass and clover, dried and weighed. Numbers of flowerheads in the clover fraction were recorded. This treatment was designated CC1.
The remaining microsites in each pipe section were divided into two groups of 4 microsites: those on the north side and those on the south side of the pipe section, as in experiment A. The 4 microsites in each group were designated at random to 4 treatments: CGU1, CGU3, CGH1 and CGH3.
Treatment CGU1 was planted with three perennial ryegrass seedlings, and no N was applied, instead 100 ml of distilled water was applied to the transplants. CGU1 was cut down to ground level on 6.9.00 and 18.10.00, and the cuttings were returned to the soil surface. On 24.5.01 and 13.8.01, CGU1 was cut and the herbage was separated into grass and clover fractions, dried and weighed. Numbers of flowerheads in the clover fraction were recorded.
Treatment CGU3 was planted with three perennial ryegrass seedlings. N was applied at a rate of 5 g N m-2, by dissolving 0.185 g of (NH4)2SO4 in 100 ml of water for each microsite. CGU3 was cut down to ground level on 6.9.00 and 18.10.00, and the cuttings were returned to the soil surface. On 24.5.01 and 13.8.01, CGU3 was cut and the herbage was separated into grass and clover fractions, dried and weighed. Numbers of flowerheads in the clover fraction were recorded.
Treatment CGH1 was planted with three perennial ryegrass seedlings. No N was applied, but 100 ml of distilled water was applied after transplanting. CGH1 was harvested on 6.9.00, 18.10.00, 24.5.01 and 13.8.01. Herbage was separated into grass and clover fractions, dried and weighed. Numbers of flowerheads in the clover fraction were recorded.
Treatment CGH3 was planted with three perennial ryegrass seedlings. N was applied as in treatment CGU3. CGH3 was harvested on 6.9.00, 18.10.00, 24.5.01 and 13.8.01. Herbage was separated into grass and clover, dried and weighed. Numbers of flowerheads in the clover fraction were recorded.
The plastic barrier around the central microsite was removed on 28.1.01. Any seedlings of weeds or grass that were not deliberately planted were pulled up and cut and mulched into the microsite in which they were found.
Experiment D
Three perennial ryegrass seedlings were transplanted into the central microsite of each pipe section. No N was applied. The herbage was cut on 6.9.00, 18.10.00, 24.5.01 and 13.8.01, and separated into grass and clover, dried and weighed. Numbers of flowerheads in the clover fraction were recorded. This treatment was designated DG1.
The remaining microsites in each pipe section were divided into two groups of 4 microsites: those on the north side and those on the south side of the pipe section, as in experiment A. The 4 microsites in each group were designated at random to 4 treatments: DCU1, DCU3, DCH1 and DCH3.
DCU1 was planted with three white clover seedlings. DCU1 was cut down to ground level on 6.9.00 and 18.10.00, and the cuttings were returned to the soil surface. On 24.5.01 and 13.8.01, DCU1 was cut and the herbage was separated into grass and clover fractions, dried and weighed. Numbers of flowerheads in the clover fraction were recorded.
Treatment DCU3 was planted with three perennial ryegrass seedlings. N was applied as in experiment C. DCU3 was cut down to ground level on 6.9.00 and 18.10.00, and the cuttings were returned to the soil surface. On 24.5.01 and 13.8.01 DCU3 was cut and the herbage was separated into grass and clover fractions, dried and weighed. Numbers of flowerheads in the clover fraction were recorded.
Treatment DCH1 was planted with three perennial ryegrass seedlings. No N was applied. 100 ml of distilled water was applied to the transplants. DCH1 was harvested on 6.9.00, 18.10.00, 24.5.01 and 13.8.01. Herbage was separated into grass and clover fractions, dried and weighed. Numbers of flowerheads in the clover fraction were recorded.
Treatment DCH3 was planted with three perennial ryegrass seedlings. N was applied as in Experiment C. DCH3 was harvested on 6.9.00, 18.10.00, 24.5.01 and 13.8.01. Herbage was separated into grass and clover, dried and weighed. Numbers of flowerheads in the clover fraction were recorded. The plastic barrier around the central microsite was removed on 28.1.01.
Any seedlings of weeds or grass that were not deliberately planted were pulled up and cut and mulched into the sector in which they were found. Any clover stolons that grew over the plastic partitions between the peripheral microsites, were cut at the point at which they crossed the barrier, and then chopped and returned to the microsite in which they originated.
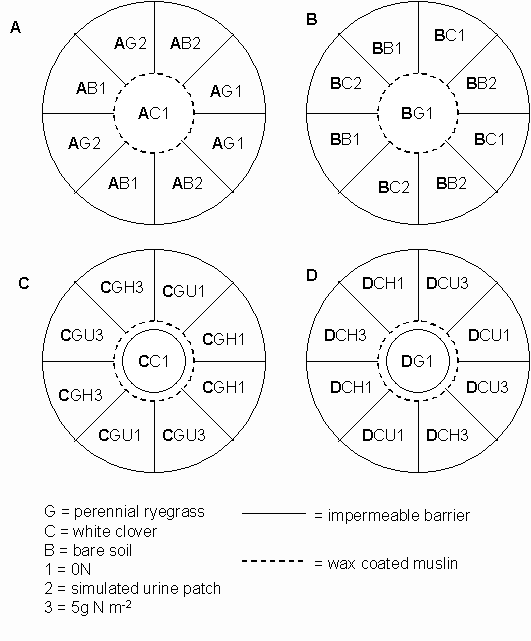
Figure .1 Treatments in the invasion experiments. Each circle represents one of the replicates of experiment A, B, C or D. Letters in bold are codes for experiments A, B, C and D.
Invasions
Clover that colonised bare microsites or microsites initially planted with grass was designated as invading clover. Grass that colonised bare microsites or microsites initially planted with clover was designated as invading grass. Figure 7.4.1 shows yields of invading clover in microsites which originally contained only grass or bare soil. Treatment BG1 (central microsite of experiment B, containing unfertilised grass harvested on all sampling dates), was invaded on the first sampling date, and subsequently colonised by clover to a far greater extent than other treatments (Figure 7.4.1). The other grass and bare soil microsites were not invaded by clover until the final sampling date, if at all.
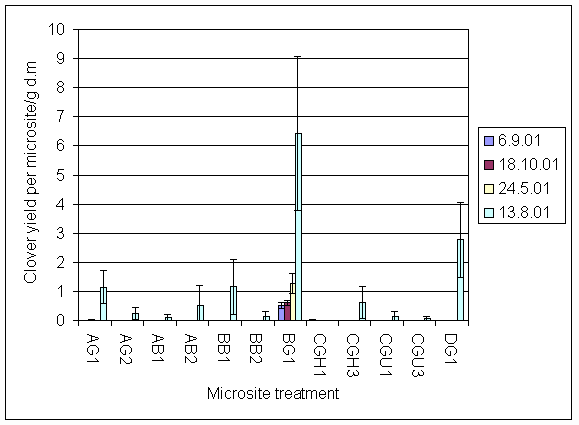
Figure .1 Yield of invading clover on all four sampling dates AG1 = experiment A, grass, unfertilised; AG2 = experiment A, grass + urine; AB1 = experiment A, bare soil, unfertilised; AB2=experiment A, bare soil + urine; BB1=experiment B, bare soil, unfertilised; BB2 = experiment B, bare soil + urine; BG1 = experiment B, grass, unfertilised; CGH1 = experiment C, grass, harvested, unfertilised; CGH3 = experiment C, grass, harvested + 5 g N m-2; CGU1 = experiment C, grass, unharvested, unfertilised; CGU3 = experiment C grass, unharvested + 5 g N m-2; DG1 = experiment D, grass, unharvested, unfertilised.
Grass showed very little tendency to invade bare or clover microsites. Invasion by grass was sometimes transient, i.e. grass invaded a microsite and then died off in that microsite. The average mass of invading grass was not significantly greater than 0 in any treatment
(Figure 7.4.2).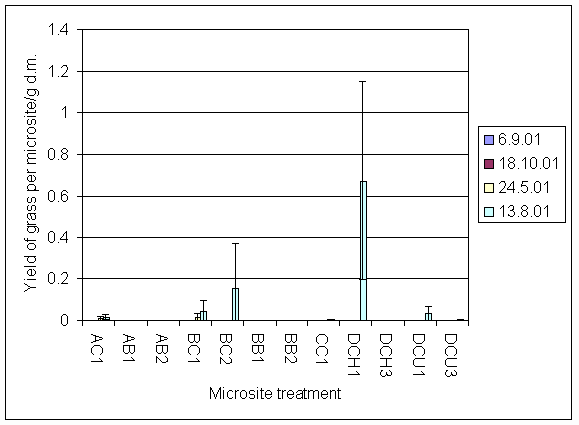
Figure .2 Yield of invading grass on all four sampling dates. AC1 = experiment A, clover, unfertilised; AB1 = experiment A, bare soil, unfertilised; AB2 = experiment A, bare soil + urine; BC1 = experiment B, clover, unfertilised; BC2 = experiment B, clover + urine; BB1 = experiment B, bare soil, unfertilised; BB2 = experiment B, bare soil + urine; CC1 = experiment C, clover, unfertilised; DCH1 = experiment D, clover, harvested, unfertilised; DCH3 = experiment D, clover, harvested + 5 g N m-2; DCU1 = experiment D, clover, unharvested, unfertilised; DCU3 = experiment D clover, unharvested + 5 g N m-2.
Clover yields in the microsites initially planted with clover were highest in the final sample in most cases. Clover in the central microsites of experiment C (CC1) yielded poorly (Figure 7.4.3) and also did not invade other microsites much in comparison to other clover treatments (Figure 7.4.1
).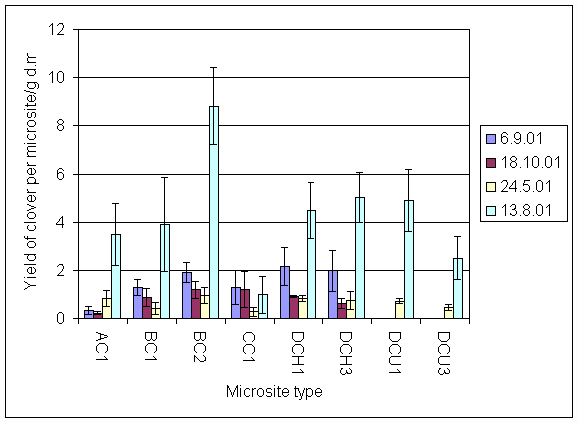
Figure .3 Yield of clover in microsites initially planted with clover on all four sampling dates AC1 = experiment A, clover, unfertilised; BC1 = experiment B, clover unfertilised; BC2 = experiment B, clover + urine; CC1 = experiment C, clover, unfertilised; DCH1 = experiment D, clover harvested, unfertilised; DCH3 = experiment D, clover, harvested + 5 g N m-2; DCU1 = experiment D, clover, unharvested, unfertilised; DCU3 = experiment D clover, unharvested + 5 g N m-2.
In the microsites initially planted with grass, the highest grass yields were in the central microsites of experiment B. Grass yields were higher in this microsite than in any other on all sampling dates (Figure 7.4.4). In the second year of sampling, the total grass yields in the cut and mulched treatments (CGU1 and CGU3) were higher than the total grass yields in the previously harvested treatment (CGH1 And CGH3, ANOVA p<0.05). Grass yields in urine treated microsites (AG2) were higher than grass yields in untreated microsites (AG1), but the difference was not significant. Smaller additions of N had no effect on grass yield.
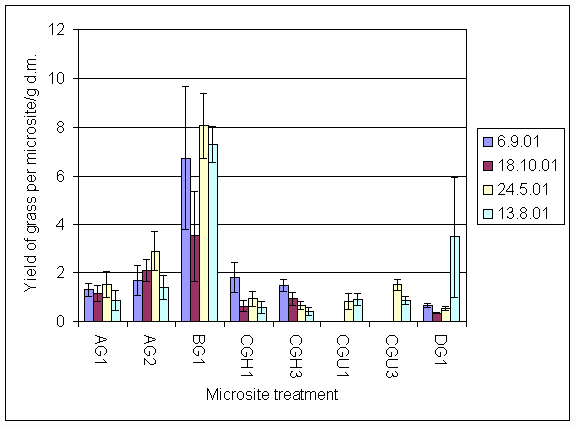
Figure .4 Yield of grass in microsites initially planted with grass on all four sampling dates. AG1 = experiment A, grass, unfertilised; AG2 = experiment A, grass + urine; BG1 = experiment B, grass, unfertilised; CGH1 = experiment C, grass, harvested, unfertilised; CGH3 = experiment C, grass, harvested + 5 g N m-2; CGU1 = experiment C, grass, unharvested, unfertilised; CGU3 = experiment C grass, unharvested + 5 g N m-2; DG1 = experiment D, grass, unharvested, unfertilised.
The number of clover flowerheads followed a similar pattern to clover yield, both in invaded microsites and in microsites initially planted with clover (Figure 7.4.5, Figure 7.4.6, Figure 7.4.1 and Figure 7.4.3). There were no significant differences in flowering rate (number of flowers per g clover dry matter) between treatments.
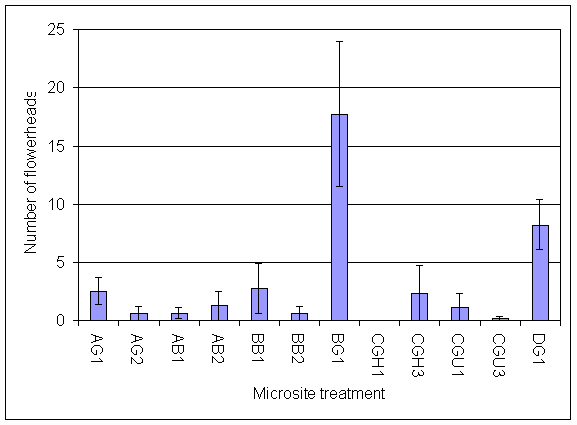
Figure .5 Number of flowerheads of invading clover on 13.8.01 AG1 = experiment A, grass, unfertilised; AG2 = experiment A, grass + urine; AB1 = experiment A, bare soil, unfertilised; AB2 = experiment A, bare soil + urine; BB1=experiment B, bare soil, unfertilised; BB2 = experiment B, bare soil + urine; BG1 = experiment B, grass, unfertilised; CGH1 = experiment C, grass, harvested, unfertilised; CGH3 = experiment C, grass, harvested + 5 g N m-2; CGU1 = experiment C, grass, unharvested, unfertilised; CGU3 = experiment C grass, unharvested + 5 g N m-2; DG1 = experiment D, grass, unharvested, unfertilised.
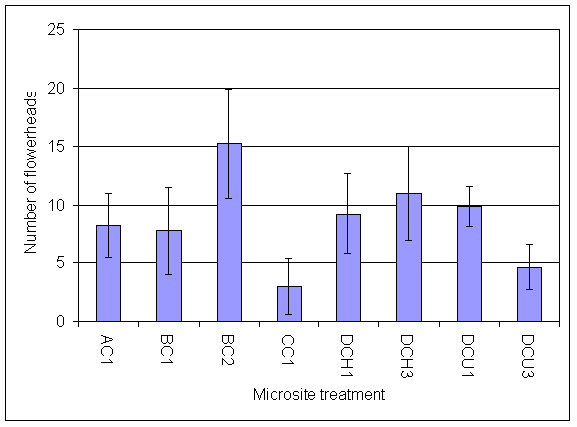
Figure .6 Number of flowerheads of clover in microsites initially planted with clover on 13.8.01. AC1= experiment A, clover, unfertilised; BC1 = experiment B, clover, unfertilised; BC2 = experiment B, clover + urine; CC1 = experiment C, clover, unfertilised; DCH1 = experiment D, clover, harvested, unfertilised; DCH3 = experiment D, clover, harvested + 5 g N m-2; DCU1 = experiment D, clover, unharvested, unfertilised; DCU3 = experiment D, clover, unharvested + 5 g N m-2.
In experiment A, the average mass of invading clover was significantly higher in unfertilised grass than in grass patches fertilised with 30 g N m-2 at the final sampling date (ANOVA p=0.01). The total yield of invading clover over both growing seasons was higher in unfertilised than fertilised grass (ANOVA p=0.04), and unfertilised grass was invaded before fertilised grass and bare soil. The numbers of fertilised grass zones and unfertilised grass zones invaded were the same. The mass of grass in urine treated microsites was greater than that in untreated microsites, but the difference was not significantly different.
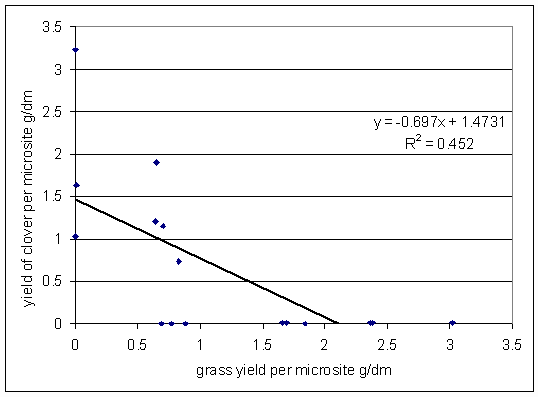
Figure .7 Yields of grass and invading clover, experiment A. 13.8.01. Each point represents the grass and clover yields for one of the microsites initially planted with grass (treatments AG1 and AG2).
The average mass of invading clover in unfertilised bare ground was not significantly different from that of clover invading bare ground with 30 mg of N. Mass of invading clover was significantly higher in unfertilised grass than in unfertilised bare soil (ANOVA p=0.02). Grass microsites with low grass yields were invaded by clover to a greater extent than those with high grass yields (Figure 7.4.7). Mass of invading clover in fertilised grass and fertilised bare soil were not significantly different. Half of the central clover zones were invaded by grass (Table 7.4.1).
|
Date |
|||||
|
Initial treatment |
Invading species |
6.9.00 |
18.10.00 |
24.5.01 |
13.8.01 |
|
AG1 |
Clover |
0 |
12.5 |
0 |
75 |
|
AG2 |
Clover |
0 |
0 |
0 |
75 |
|
AC1 |
Grass |
0 |
0 |
25 |
50 |
|
AB1 |
Grass |
0 |
0 |
0 |
0 |
|
Clover |
0 |
0 |
0 |
37.5 |
|
|
AB2 |
Grass |
0 |
0 |
0 |
0 |
|
Clover |
0 |
12.5 |
0 |
25 |
|
Table .1 Experiment A: % of artificial microsites invaded by species alien to that microsite on four sampling dates and five treatments. AG1 = grass, untreated; AG2 = grass + artificial urine; AC1 = Clover, untreated; AB1 = bare soil, untreated; AB2 = bare soil + artificial urine.
In experiment B, grass invaded less than half of the zones and the mass of invading grass was negligible (Table 7.4.2). Clover invaded all of the central grass zones, before the first sampling date. Urine treated clover patches had higher clover yields in the final sample (ANOVA p=0.02), but not in earlier samples. Some clover stolons spread through the central zone into bare microsites, but the average yield of clover in these zones was not significantly greater than 0. No grass successfully colonised bare microsites. Yield of invading grass was not significantly greater than 0 in any treatment.
|
Date |
|||||
|
Initial treatment |
Invading species |
6.9.00 |
18.10.00 |
24.5.01 |
13.8.01 |
|
BC1 |
Grass |
0 |
0 |
12.5 |
37.5 |
|
BC2 |
Grass |
0 |
0 |
0 |
37.5 |
|
BG1 |
Clover |
100 |
100 |
100 |
100 |
|
BB1 |
Grass |
0 |
0 |
12.5 |
0 |
|
Clover |
0 |
0 |
0 |
37.5 |
|
|
BB2 |
Grass |
0 |
0 |
0 |
0 |
|
Clover |
0 |
0 |
0 |
37.5 |
|
Table .2 Experiment B: % of artificial microsites invaded by species alien to that microsite on four sampling dates and five treatments. BC1 = clover, untreated; BC2 = clover + artificial urine; BG1 = grass, untreated; BB1 = bare soil, untreated; BB2 = bare soil + artificial urine.
In Experiment C, mass of clover was generally low. Clover invaded 35% of the surrounding zones and mass of invading clover was also low. Grass did not significantly invade the central clover zone (Table 7.4.3., Figure 6.4.1)
However, when clover was grown alongside patches of grass which had been harvested for a year and unfertilised, and patches which had been harvested for a year and fertilised with 5 g N m-2 the clover was more likely to invade the fertilised grass patch. Cutting and mulching of grass appeared to deter invasion by clover, regardless of whether 5 g N m-2 was applied (Figure 7.4.1, Table 7.4.3). Total yields of clover and grass were not significantly affected by harvesting and N application (Figure 7.4.3., Figure 7.4.4). Dry matter of invading clover in harvested untreated grass zones in experiment C, was not significantly different from dry matter of invading clover in untreated grass in experiment A.
|
Date |
||||
|
Initial treatment |
Invading species |
24.5.01 |
13.8.01 |
|
|
CGH1 |
Clover |
12.5 |
25 |
|
|
CGH3 |
Clover |
0 |
50 |
|
|
CGU1 |
Clover |
0 |
50 |
|
|
CGU3 |
Clover |
0 |
25 |
|
|
CC1 |
Grass |
0 |
25 |
|
Table .3 Experiment C: % of artificial microsites invaded by species alien to that microsite on the final two sampling dates (after removal of the plastic barriers). CGH1 = grass, harvested, unfertilised; CGH3 = grass, harvested + 5 g N m-2; CGU1 = grass, unharvested; CGU3 = grass, unharvested + 5 g N m-2; CC1 = clover, harvested, unfertilised.
In experiment D, total yield of invading grass in 2001 was significantly higher in untreated, harvested clover than in other treatments (ANOVA p=0.02, Figure 7.4.2). The majority of untreated, harvested clover microsites were invaded by grass, whereas none of the harvested clover microsites, with applied N were in this experiment. All of the central grass microsites were invaded by clover, although clover was slower to invade these microsites, than the central grass microsites in experiment B (Table 7.4.4,Figure 7.4.1)
|
Date |
|||
|
Initial treatment |
Invading species |
24.5.01 |
13.8.01 |
|
DCH1 |
Grass |
12.5 |
62.5 |
|
DCH3 |
Grass |
0 |
0 |
|
DCU1 |
Grass |
12.5 |
37.5 |
|
DCU3 |
Grass |
0 |
12.5 |
|
DG1 |
Clover |
0 |
100 |
Table .4 Experiment D: % of artificial microsites invaded by species alien to that microsite on the final two sampling dates (after removal of the plastic barriers). DCH1 = clover, harvested, unfertilised; DCH3 = clover, harvested + 5 g N m-2; DCU1 = clover, unharvested; DCU3 = clover, unharvested + 5 g N m-2; DG1 = clover, harvested, unfertilised.
This experiment was set up to test the following hypotheses:
Hypothesis a) stated that under controlled conditions, clover will elevate soil N and increase the yield of grass grown nearby. Clover did elevate the yield of grass grown alongside it in these experiments. The yield of grass in BG1 (grass grown surrounded by clover) on the first two sampling dates was considerably higher than the yield of grass in DG1 (grass grown surrounded by clover with the roots separated by a plastic barrier, Figure 7.4.2, Figure 7.4.4, Table 7.4.2). Grass yields in the central microsites of experiment D were low, but rose in the final sample following invasion of all grass plots in D by clover. Allowing the roots and stolons of grass and clover to freely mix clearly increased the yield of grass. This confirms the observation of Thorhallsdottir (1990a), that T.repens grew alongside gaps in the pasture less often than would be expected due to chance, perhaps because of its stimulating effect on nearby grasses (Marriott and Zuazua, 1996). Clover yields were lowest in the central clover microsites of experiment C. Possibly this is because the clover roots were constricted for the first season of growth. In one replicate of this microsite, the clover died off altogether. Clover in experiment D, which was also constrained in the first year, yielded better (Figure 7.4.3). The reasons for this are not obvious, but it may be due to the initially high ratio of grass to clover in experiment C. Grass yields in the harvested microsites in experiment C were low and declined over the course of the experiment. This is probably because harvesting of the grass in the isolated microsites depleted soil nutrients, and may also be related to the poor growth of clover in experiment C.
Hypothesis b) stated that clover will preferentially invade microsites that have not received urine or have been depleted in N by repeated cropping of grass. Clover was just as likely to invade grass microsites affected by urine as grass microsites with no urine (Table 7.4.1), but the yield of invading clover was significantly less in the urine treated microsites (Figure 7.4.1). Cropping of grass, however, did not increase the likelihood of clover invading a microsite and did not significantly increase the yield of invading clover. Clover yield was not reduced by the addition of urine: clover yields were highest in microsites that had been sown with clover and treated with urine. This is probably an experimental artefact, due to the lack of grass in these plots, which would otherwise have grown at the expense of clover.
Hypothesis c) stated that grass will preferentially invade microsites that have received urine, and have not been depleted in N. Grass was no more likely to invade urine treated microsites (whether occupied by clover or bare soil, but this was probably because ryegrass didn’t change its distribution very much over the course of the experiment (Figure 7.4.2, Table 7.4.1). It is possible that if the experiment was continued for longer, invasion of urine affected patches by ryegrass may have been observed, however, the effects of urine patches on pasture are generally short lived. The effects of urine on white clover pNdfa are greatest after 40 days, and clover growth starts to recover after 120 days (Vinther, 1998), so it is unlikely that ryegrass would spread vegetatively into a urine patch before the extra nitrogen in the patch had leached out of the soil. In the field, ryegrass could replace clover in a microsite by the germination of seeds, or the growth of small grass plants already present in the microsite, previously suppressed by the clover. Vinther (1998) showed that applications of urine increased grass yield by between 42 and 76%, depending on the time of year. This study confirmed this: grass treated with urine yielded approximately twice as much dry matter as grass untreated by urine. The increased grass yield resulting from urine applications was, however, small compared to the increased grass yield that resulted from the presence of clover: treatment BG1 had approximately 4 times as much grass as other microsites (Figure 7.4.4). Vinther (1998) also observed that urine applications also reduced the proportion of clover by 32-39%. This was not observed in experiment B: Clover actually grew slightly better when treated with urine. However, this was probably due to the lack of competition from grass. In experiment B, urine was applied to clover monocultures, and ryegrass was very slow to invade.
Hypothesis d) stated that both clover and grass will preferentially invade vacant microsites in preference to occupied microsites. Surprisingly, clover did not behave as a pioneer species, as it was more likely to invade grass microsites than bare microsites. Only one instance of clover invading a bare microsite was observed in experiment A. Some invasion of bare microsites by clover was observed in experiment B (stolons spreading via the central microsite), indicating that invasion of bare microsites was possible under these circumstances. Grass was also more likely to invade clover microsites than bare microsites, disproving hypothesis d. There is some support for this in the literature: Turkington and Joliffe (1996) observed that under certain circumstances, white clover could benefit from the presence of perennial ryegrass, and Thorhallsdottir (1990 a,b) observed that white clover grew alongside L.perenne, Cynosurus cristatus (crested dogstail) and Anthoxanthum odoratum (sweet vernal grass) more frequently than would be expected due to chance in the field. In artificial swards, clover showed a preference for invading Poa trivialis and L.perenne microsites. The dry matter of invading clover was greater in microsites initially sown with grasses, than in initially bare microsites, but this may have been due to rapid invasion and vacation of these microsites during the 27 months of the experiment (Thorhallsdottir, 1990b).
The available literature strongly supports the idea that white clover will proliferate on bare soil: Harris (1987) states that "The apparent antagonism between white clover and stoloniferous or rhizomatous species possibly results from competition for suitable rooting sites by the prostrate stems of these species as they wander through the sward. Where the associated species are tufted, bare soil is ample between the tufts, and in these inter-tuft areas white clover stolons are mostly found". In these experiments, the ryegrass grew mainly in discreet clumps, but the clover was more likely to successfully invade these clumps than the bare soil in between them. It would be interesting to repeat this experiment with a non-leguminous creeping perennial such as buttercup (Ranunculus spp.) in addition to white clover.
Most studies of this kind have found that white clover stolons elongate and branch in response to increased light intensity, and divert their energy from stolon to leaf and petiole production when shaded (De Kroons and Hutchings, 1995, Heraut-Bron et al. 2001, Marriott et al. 1997a, Faurie et al. 1996, Thompson, 1993). Stolon length and branching were not recorded in this study, but the presence of stolons in a microsite was recorded, even if there was no herbage above stubble height. There was no evidence of increased proliferation of stolons in bare microsites (Table 7.4.1, Table 7.4.2). The percentage of microsites invaded and the biomass of invading clover were not very closely correlated, suggesting that the proliferation of leaves and roots may be as important as the growth of stolons in the colonisation of microsites by white clover, as suggested by De Kroons and Hutchings, (1995).
The results of these experiments did not fully support the cyclical replacement hypothesis. Ryegrass was not capable of rapidly colonising soil microsites by vegetative growth, although it did invade clover microsites, more than bare microsites, the mass of invading grass was too low to be significant. Urine patches (whether clover or bare soil) were no more likely to be colonised by grass, and the addition of small amounts of N also didn’t make microsites any more susceptible to invasion by grass (Figure 7.4.2, Table 7.4.1). On the contrary, addition of small amounts of N and/or mulching of clover seemed to make microsites less susceptible to invasion by grass. It is possible that in a more natural situation, grasses may regenerate from seed more frequently than in this experiment. Also the creation of microsites containing only clover in this experiment was artificial. In the field, clover microsites would contain some grass. Such a microsite could change from clover to grass if grass plants already present within the microsites grew and suppressed the clover. Invasion of the microsite by grasses might not be necessary for the vegetation to change.
Clover and grass were therefore capable of distinguishing between different types of microsite, and invading some more than others. Some microsites were invaded by clover, but not successfully colonised. It may be that stolons grow in a largely random fashion in all directions, but shading and/or unsuitable soil conditions restrict their growth and/or branching. Cutting and mulching of vegetation, and the presence or absence of vegetation seemed to have more effect on invasion than the addition of N. Subsequent establishment and growth of the invading species could, however, be affected by the addition of N.
Discussion
This thesis set out to examine the interactions between grass, clover and soluble soil N in the ley phase of an organic ley arable rotation. Specifically it attempted to measure the effects of a grass-clover ley on soluble soil N in the field, and the effects of grass and clover on soluble soil N under experimental conditions. It also examined the changes in grass and clover yield, N fixation, proportion of N in clover derived from fixation, grass N concentration and clover N concentration over the course of the ley phase, between rotations, and over time. These variations were compared with the changes in soil N, to see to what extent they were related.
Hypotheses 1 and 4 were tested in Section 2. Hypotheses 2 and 3 were tested in Sections 2 and 3. Hypothesis 5 was tested in Sections 4 and 5. Hypotheses 6, 7 and 8 were tested in Section 5.
Hypothesis 1Mineral N (nitrate-N + ammonium-N) concentrations were higher in soil in 3 and 4-year old leys than in 1-year-old leys at Tulloch, confirming hypothesis 1. Other studies have observed increases in total soil N and soil mineral N over the course of a grass/legume ley (Hossain et al., 1996), something that is not always observed in grass-only leys (Korsaeth et al., 2003). Nitrate-N did not however, increase steadily over the lifetime of the ley. This may have been because of management of the leys. Grazing of 1- and 3-year-old leys may have increased the availability of mineral N in these stages of the rotation. 2-year-old leys in both rotations were cut for silage. It is also possible that soil N and N fixation are involved in a feedback cycle with a one year time lag: N fixation elevates soil N, which inhibits N fixation in the following year, and this in turn results in lower soil N in the subsequent year. This is similar to the effect postulated by Loiseau et al. (2001). The highest levels of nitrate-N were observed in 3-year-old leys, and the lowest in 2-year-old leys. This may have been related to the management of the leys, as 2-year-old leys in both rotations were cut for silage. In this study, clover content of the sward varied less than total sward herbage N yield over the period studied, and this contradicts the observation of Ledgard (2001), that variations in the clover content of the sward are greater than variations in total herbage yield.
The N fixation data suggests that inputs of fixed N from white clover are likely to decline after the third year of ley, and that the fourth year of ley growth in this system could be an N depleting stage of the rotation, depending on the amount of manure applied. This confirms the findings of Johnston et al. (1994) and Clement and Williams (1962), that a ley phase longer than 3 years did not significantly improve crop yields, or availability of N.
Estimates of the N budget based on the experimental subplots at Tulloch, suggest that 1-year-old leys should have a small positive N balance (input – removal), of approximately 15 kg N ha-1, 2-year-old leys 97 kg N ha-1, 3-year-old cut leys –128 kg N ha-1, 3-year-old grazed leys –34 kg N ha-1 and 4-year-old leys 9.9 kg N ha-1. This explains the increase in soil mineral N between years 2 and 3 of the crop rotation, and is roughly consistent with the other changes in soil mineral N observed over the course of the crop rotation. The N balance would, in almost all cases, be negative, if manure was not returned. Ledgard (2001) suggests that grass-legume systems are essentially self-regulating, with regard to N, and this means that losses of N through leaching are likely to be small. Self-regulation of the type hypothesised by Ledgard (2001) was not observed in this study. This system differed from the type of system studied by Ledgard (2001) in a number of ways: The system featured both cut and grazed swards. The cut swards received manure, and although the manuring rate was adjusted to take fluctuations in sward clover content, it could still have potentially unbalanced the system. Also it should be remembered that the estimates of N fixation were made from small subplots which did not receive the same treatment as the rest of the sward (no manure or excreta, cut monthly). The subplots were also moved between years, so they may not precisely represent the situation in the field as a whole.
Most of the N in soil is bound up in soil organic matter (Bjarnasson, 1988). A small fraction of this is released into solution as nitrate and ammonium, by the processes of mineralisation. The rate of mineralisation is an indicator of soil fertility, and is related to the amount of N in the soil, and the C:N ratio of the organic inputs (Alvarez et al. 1998, Bjarnasson, 1988). Clover residues, dung and urine would therefore increase mineralisation of N, more than grass residues.
There was no evidence from this study that N fixation or clover yield was negatively correlated with soil nitrate-N, ammonium-N, TSN or SON. Clover yield followed different seasonal, annual and age related patterns to soil N. pNdfa was negatively correlated with soil nitrate and TSN, but this did not significantly affect N fixation, because N fixation was more strongly affected by clover yield than pNdfa or %NC. There is no evidence from this that soil N is having an immediate and direct effect on N fixation as the cyclical replacement hypothesis implies. This is in contrast to the findings of Hansen et al. (2002), who observed negative correlations between clover dry matter yield and mineral N (Pearson 0.250, p<0.05) and clover N yield and mineral N (Pearson 0.278, p<0.05) on an organic farm in Denmark. Some of the differences may be due to the fact that Hansen et al. (2002) studied a two-year-old ley previously grazed by cattle, whereas at Tulloch, 2-year-old leys were ungrazed prior to sampling. Ledgard (2001) observed that pNdfa was reduced in grazed swards, owing to N uptake from excreta. This study confirmed this: Soil nitrate-N was observed to be higher in grazed leys than in cut leys, and pNdfa was also slightly lower in swards that had been grazed prior to sampling. As in this study, Hanson et al. (2002) observed a negative correlation between pNdfa and nitrate-N (Pearson –0.403, p<0.01). The situation on the Danish farm differed from Tulloch in that pNdfa was lower (average 0.6), more variable, and negatively correlated with clover yield (Pearson –0.304, p<0.05). The correlation between N fixation and clover yield was much lower than in this study (Pearson 0.432, p<0.01, Hansen et al., 2002). However, soil extractable P was significantly correlated with clover yield, so it is possible that rather than elevating N locally, clover may deplete P to a point at which it cannot compete with grass. pH was also correlated with clover yield. pH affects clover yield and is also affected by clover, so pH or P could also cause cyclical replacement in a grass-clover sward.
Grass yield was positively correlated with nitrate-N, confirming the findings of Hansen et al. (2002), and was also correlated with TSN. N concentration of grass was positively correlated with nitrate-N, TSN and SON. Soil derived N was significantly correlated with nitrate-N and TSN. Unlike clover, grass was clearly responding to the level of available N in the soil at Tulloch, as predicted by the cyclical replacement hypothesis. The variation of grass N concentration and grass yield (and consequently grass N yield) between different years and ages of ley was closely related to variations in soil mineral N between years and ages of ley.
Soil derived N was closely related to grass N yield, but in a few instances, clover herbage contained significant amounts of soil derived N as well. Other studies have found that soil derived N in clover can be highly significant, indicating that under some conditions clover can compete effectively with grass for soil N (Boller and Nosberger, 1987).
Clover microsites had higher nitrate-N than grass microsites in summer, as predicted by the cyclical replacement hypothesis. This confirms the observations of Antil et al. (2001) and Ledgard et al. (1998) that N mineralisation is elevated in the presence of clover, probably as a result of the low C:N ratio of the plant residues. Evidence that clover was growing in low-N microsites during the winter was less clear, but this may have been due to the highly variable soil N values observed during the winter. Weeda (1967), observed that during the winter months in a New Zealand pasture, white clover was almost entirely restricted to dung patches. The study also drew attention to the high concentrations of calcium, magnesium and potassium in cattle dung. It is possible that the seasonal distribution of clover in pastures is influenced by the distribution of these nutrients, rather than N. Alternatively, winter grazing pressure may have caused a decline in clover everywhere in the sward except in the rejected areas around dung patches.
No significant effects of clover on soil N were observed in the root exudation experiment. This may reflect the unnatural conditions of the experiment (e.g. application of N free fertiliser), that may have encouraged the growth of free-living N fixing bacteria. The experimental conditions may also have excluded organisms such as slugs and root weevils, which could facilitate N transfer in the field. Another possible reason is that the root exudation experiment used clover seedlings in their first year of growth. N transfer estimates are sometimes low in the first year of clover growth (Høgh-Jensen and Schjoerring, 1997). High fertility soils like the one used in this experiment can also reduce the effect of clover/grass mixtures on N mineralisation (Alvarez et al., 1998)
It is possible that there was greater N leaching from microsites containing clover than from those containing only grass. Grassland swards containing legumes lose more N through leaching than swards containing only non-legumes (Scherer-Lorenzen et al., 2003, Alvarez et al., 1998), increasing the diversity of the sward can reduce this effect (Scherer-Lorenzen et al., 2003). Scherer-Lorenzen et al. (2003) found that pasture mixtures containing legumes lost more N through leaching than those containing only non-legumes. In addition, diverse species mixtures containing legumes lost less N to groundwater than mixtures of legumes and a single non-legume species. In contrast, Froment et al. (1999) observed that mineral N under land sown with legumes was highly variable, but in some cases, lower than mineral N in land sown to ryegrass with no fertiliser N, especially in Autumn.
In the field in 2000, when the most accurate comparisons could be made, grass growing in mixtures with clover had higher N yields and N concentrations than grass grown as a monoculture, confirming the findings of Matthew et al. (1995). Grass dry matter yields were, however, similar or lower in mixtures than in monocultures. This probably indicates that the grass growing alongside clover was well supplied with N, but its dry matter yield was restricted by the presence of the clover, and competition for light, water or soil nutrients. Ledgard (2001) observed from a review of the available evidence that in intensively grazed swards, N transfer from grass to clover below ground is of a similar magnitude to N transfer above ground, whereas in extensively grazed swards below ground transfer is more important. In this study, N transfer below ground was estimated at between 0 and 28 kg N ha-1, and N transfer above ground at around 40 kg N ha-1. Both estimates may be subject to large errors and innaccuracies. N transfer below ground may be a much slower process than N transfer through dung and urine (Ledgard, 2001)
In the pot experiments, grass yields were similar in mixtures and monocultures. N concentration of grass in this experiment had not been measured at the time of writing, so it is not possible to say whether N yield of grass was increased by the presence of clover under these conditions. Pot experiments similar to this one, in which plant roots are confined do not always produce comparable results to field sampling in which soil with and without roots is sampled (Haynes and Beare, 1997)
In the invasion experiment there was some evidence that grass benefited from the presence of clover. This may have been due to the N sparing effect in some cases. Clear evidence that clover was elevating soil mineral-N was not observed in the root exudation experiment.
Improved growth of grass when sown in a mixture with clover may not be entirely due to improved soil N status. Clover also improves soil structure, aggregate stability and drainage (Mytton et al., 1993, Haynes and Beare, 1997), and this could benefit the growth of neighbouring plants and subsequent crops after ploughing of the ley.
Successful invasion of clover into microsites containing grass was significantly less likely if the grass had been treated with urine, however, harvesting and removal of grass did not significantly increase the likelihood that a microsite would be successfully invaded by clover. The amount of N removed in the grass harvested in the first year of invasion experiment C, was not measured. If it is assumed that the N concentration of grass in the invasion experiment was similar to that observed in 1-year-old grass monocultures in the field in 2000 (2.397%, 3.34%, 1.423% and 2.1533%, for September, October, May and August, respectively), then the harvesting of grass in the harvested treatments, removed the equivalent of 8.2 and 8.4 g N m-2, in 2000, in the untreated and N treated harvested plots (CGH1 and CGH3), respectively. The addition of small amounts of N did not, therefore, significantly affect the yield of grass. The amount of N removed from the soil in the harvested grass in experiment C, was of the same order of magnitude as the amount of N added to treatments CGU3 and CGH3. The N in grass mulch would not be immediately available to grass or clover, and recovery of N from the mulch in the short term could be as low as 13% (Riley et al., 2003). It is unlikely, therefore, that the preferential invasion of harvested grass microsites by clover is entirely due to differences in soil N levels. A grass mulch could have other effects on the soil, such as preventing rooting of clover stolons, or reducing evaporation of water from the soil surface (Seneviratne et al., 1998). More regular harvesting of ryegrass from earlier in the growing season (more closely simulating grazing) could have resulted in greater soil N depletion than was observed. The addition of urine did appear to increase grass yields, although the difference was not significant overall. This was mainly because of considerable variation in grass yields between microsites.
Although clover invaded bare soil microsites less successfully than microsites containing grass, microsites with low yields of grass were more likely to be invaded than microsites with high grass yields. There is a general consensus in the literature that competition between clover and grass for light is crucial to grass clover interactions, and that shading by grass reduces clover stolon branching (Markuvitz and Turkington, 2000, Wilman and Fisher, 1996, Thompson, 1993). The suppression of clover by grass when N is applied may be because the additional N enables grass to shade out clover (Wilman and Fisher, 1996, Faurie et al. 1996, Laidlaw and Withers, 1998). These results are consistent with this, but the effects seen could also be due to competition between clover and grass for water or soil nutrients other than nitrogen.
Laidlaw and Withers (1998) and Acuna and Wilman (1993) both showed the importance of light competition in determining the ratio of grass to clover. Elevated nitrogen seems to decrease the ratio of clover to grass, by increasing the ability of grass to compete for light. Close cutting increases the number of clover growing points (Wilman and Acuna, 1993). The addition of P can have the same effect (Bailey and Laidlaw, 1999), but this is probably because it enables the clover to compete more effectively for light, as there is no interaction between the effect of P and close cutting (Acuna and Wilman 1993).
The invasion experiment supports the hypothesis that ryegrass in urine affected microsites resists invasion by white clover. It was however not possible to draw firm conclusions about the effects of grazing on soil N and invasion of ryegrass microsites by clover. The 15N dilution experiment (Section 4) showed that yields of soil derived N in a ryegrass monoculture over an entire growing season could be as high as 17 g N m-2. The soil chemistry experiment (Section 5) showed differences in soil N between microsites containing only ryegrass and those containing clover, which either indicates elevation of soil N by clover, depletion of soil N by grass or both. It is likely therefore that an area of sward containing no clover, that was grazed for a growing season would be depleted in mineral N sufficiently to affect the invasion of clover. This is confirmed by the study of Korseath et al. (2003) which observed net removal of N from cut grass monocultures, and depletion of soil organic matter, even with fertiliser or manure applications.
Ryegrass showed very little tendency to invade microsites under any circumstances in the invasion experiment. There was very little evidence therefore that ryegrass is capable of changing its distribution significantly in response to dung and urine deposition within the space of one or two growing seasons. This contrasts with the findings of Barthram et al. (2002), that perennial ryegrass could spread by 2 cm in a single year. Thorhallsdottir (1990b) grew grass and clover species in an experimental sward similar to the invasion experiments and observed that after 27 months of growth (from May 1978 to May 1980) more than 80% of the perennial ryegrass present was in the microsites in which it had originally been sown. In contrast, less than 25% of the clover was in the microsites in which it had originally been sown. In invasion experiment B, only 5% of the ryegrass present at the end of the experiment had invaded other cells, but 53% of the clover sown in experiment A had spread to neighbouring microsites. In the Tulloch leys, ryegrass and timothy were present almost everywhere except for the densest clover patches. It is likely, therefore, that changes in soil N could cause clover to become extinct in a microsite, and replaced by grass even if there was no actual invasion of the microsite by grass stolons. Invasion of microsites by ryegrass in the field may be due to seed germination as much as vegetative spread. Milbau et al. (2003) found that Lolium perenne was highly invasive compared to other common grass species studied (Arrhenatherum elatius, Agrostis tenuis, Dactylis glomerata, Festuca rubra, Holcus lanatus and Poa trivialis). This contradicts the earlier finding of Barthram et al. (2002), that L. perenne was less invasive than F.rubra and as invasive as H.lanatus. The reason for the high invasiveness of L.perenne seemed to be that it had a high seed germination rate, especially when soil nitrate was high, and it grew rapidly in favourable conditions, suppressing and shading out other species (Milbau et al. 2003)
The evidence from invasion experiment C showed that in some cases, when clover was forced to grow in the same microsite for two seasons, while being periodically harvested, its yield was reduced, even though there was no invasion by grass. It is possible that this effect was caused by depletion of nutrients such as P or K by the clover.
Clover was more likely to invade microsites containing ryegrass than microsites with no vegetation. Ryegrass was more likely to invade microsites containing clover than microsites with no vegetation. This disproves hypothesis 8. This also contradicts much of the literature, for example Markuvitz and Turkington (2000), Wilman and Fisher (1996) and Thompson (1993) who all found that clover stolons branched more when they were exposed to direct sunlight than when they were shaded by grass. It is possible that this may have been an experimental artefact: the bare patches were created at the start of the experiment when the clover plants were at the seedling stage, and therefore unable to spread by stolons. In the field most bare patches would be adjacent to well developed grass and clover plants. This may have caused the soil to develop a crust, which could have deterred invasion by clover. Alternatively, nutrients may have leached out of the bare soil, preventing the successful invasion of the microsite by clover. However, poor establishment of clover in freshly sown pastures is quite common (Harris, 1987), but there is no evidence in the literature that the resulting bare patches persist over subsequent growing seasons.
These experiments do not entirely support cyclical replacement as it was presented in Section 2.1.1. It is clear from these experiments and others (e.g. Thorhallsdottir 1990a and b) that clover changes its spatial distribution in the sward very rapidly. The distribution of clover in the sward is influenced by urine patches, and there was some evidence that clover elevated soil mineral N by root exudation. In the field, grass yield and N concentration were positively correlated with soil nitrate-N, and pNdfa was negatively correlated with soil nitrate-N. This evidence supports cyclical hypothesis.
However, although it is well established that high soil N benefits grass at the expense of clover (Hogh-Jensen and Schjoerring, 1997, Ledgard et al. 1996, Ledgard et al. 2001), no correlation was found between soil N and clover yield in this study. This contrasts with the findings of Elgersma et al. (2000), that repeated N applications, intended to simulate natural accumulation of fixed N, had an immediate effect on clover yield. In addition, clover did not show any tendency to invade bare patches in an experimental sward, although it was more likely to invade grass microsites with low grass yields than those with high yields, and this may have been related to the availability of nitrogen in these microsites.
Elevation of soil N by clover root exudation or depletion of N by removal of grass may affect the species composition of a microsite, but this was not observed under the conditions of the invasion experiment, and so no firm conclusions can be drawn about this.
Clover yield was positively correlated with soil P and pH, and clover microsites were observed to have lower levels of extractable K than grass microsites. It is possible, therefore, that cyclical replacement in this system is occuring, but driven not by N, but by P, K and/or pH. Legumes are known to have a high requirement for P and K, and this can influence their distribution (Hogh-Jensen et al. 2002, Riffkin et al. 1999, Bailey and Laidlaw, 1999). Legumes also do not fix N effectively in acid soils (Brauer et al. 2002), and their distribution is influenced by local variations in soil pH (Gibberd and Cocks, 1997). In addition, white clover may actually acidify soil in the vicinity of its roots (Raven and Smith, 1976, Tang, C., 1998, Monaghan et al. 1998). Clover microsites in older leys also appeared to contain more moss than ryegrass, and this may be a response to increased acidification, although this observation was not tested.
The average rate of fertiliser N applied to leys (defined as grassland less than 5 years old) in North East Scotland in 2000 was 126 kg N ha-1. The equivalent figures for oats and root crops were 103 and 56 kg N ha-1 (Defra, 2001). Conventional rotations with the same sequence of ley and arable crops as the Tulloch 50% ley and 66% ley rotations would therefore use a total of 640 and 710 kg of fertiliser N per hectare, requiring an additional 22.6 and 25.1 GJ ha-1 over the 6 years of the rotation (Hullsbergen et al. 2002).
Total N fixation in 2000, including below ground fixation, fixation in early spring and N transfer at Tulloch was estimated at 214, 164, 146 and 109 kg N ha-1 for 1-, 2-, 3- and 4 year-old leys, respectively. This gives totals of 633 and 524 kg N ha-1 for the ley phases of the 66% and 50% ley rotations, respectively. The arable crops in the rotation receive manure, but as the N in this manure ultimately originates from fixed N in the leys, it has not been included in the calculation. It is not clear how much N is likely to be fixed in conventional grassland systems by white clover. Clover is present in 70% of grass seed mixtures sown in the UK, but clover only thrives in 20% of sown grassland (HERO, 2003). The average content of clover in UK grasslands is 13.2% (Firbank et al, 2000). This is less than half as much clover as was present at Tulloch during the period of the experiment, which was typically over 30% (Figure 4.6.2).
This suggests that N fixation by white clover in many conventional swards would be low, and in some cases zero. Symbiotic N fixation at Tulloch was therefore providing a similar amount of N to the system as would be applied as fertiliser in a conventional system. Yields of oats between 1997 and 2000 were on average 4.1 t ha-1 and yields of swedes were 48.1 t ha-1 (SAC data). The average yield for oats in Scotland in 2000 was 5.22 t ha-1 (Defra, 2002, SEERAD, 2001). This confirms the findings of Nieberg and Schulze Pals (1996), that yields from organic arable crops in Europe are generally about 20-40% lower than yields from equivalent conventional farms. Assuming that machinery use in conventional and organic oat production is similar to that in conventional wheat production, then conventional oat production in a ley arable rotation would require 5.76 GJ ha-1 for machinery, 3.64 GJ ha-1 for N fertiliser and 0.86 GJ ha-1 for herbicides and fungicides (Hullsbergen et al., 2002). This gives a total of 10.26 GJ ha-1. Oat production at Tulloch required an average of 6.25 tonnes of manure between 1997 and 2000. Spreading one tonne of manure requires 0.06 GJ ha-1 (Pimentel, 1983). This makes the energy input for organic oats at Tulloch 6.154 GJ ha-1. This means that the energy input per ton of oats is 1.5 GJ as opposed to 1.97 GJ t-1 in an equivalent conventional rotation.Yield data for swedes in Scotland is not available, but in England and Wales in 2003, the average yield was 30.9 tonnes per hectare (Defra, 2003). Yields from the experimental rotations at Tulloch are therefore similar to or slightly lower than yields from conventional farms in Scotland, and energy efficiency of cereal production is better than in an equivalent conventional rotation.
In all years except 1999, soil derived N in herbage exceeded N fixation in roots and stubble, and N transfer. Return of N, either by grazing animals or as composted manure in the cut swards was therefore essential for the leys to accumulate fixed N.
Improved grassland makes up 13% of the land area of Scotland (10.5 x 105 ha, Defra, 2000) and receives on average 96-127 kg N ha-1. Mixed farms in north-east Scotland typically apply 105 kg N ha-1 to temporary leys (Defra, 2001). There is therefore considerable scope for farms in this area to reduce their use of N fertilisers in favour of clover with little or no loss of yield, as many of them are already practising mixed farming, and many are only applying moderate N inputs. However, because of the losses inherent in the ploughing of grass-clover leys, economic benefits to cereal growers from a legume based system may be less clear. Organic farms are on the whole limited by the supply of N, and it is not always possible to supply N precisely when and where it is needed in organic farms (Berry et al., 2002).
Reductions in subsidies and greater liberalisation of world trade, which are forecast for the future should act to reduce the intensity of world farming. These changes, along with the increasing demand for organic produce are likely to favour legume based production systems (Ledgard, 2001). Retailers believe that the market for organic food could grow by 40% annually over the next three years (Soil Association, 2000b). In the event of a total abolition of price support subsidies to farmers, organic farmers would suffer less than conventional farmers, mainly due to the organic premium price (O’Riordan and Cobb, 2001). In addition, farm conversions to organic methods would have benefits to society as a whole. Environmental problems carry costs which can be measured. For instance, removing pesticides from drinking water costs £150 million annually (House of Lords, 1996).
Organic farms have greater diversity of grasses, spiders and birds, and more butterflies than conventional farms. They produce lower greenhouse gas emissions. Overall, the external cost of organic farmland to society as a whole can be estimated at £10-15 per hectare, as opposed to £ 25-40 per hectare for conventionally managed land (O’Riordan and Cobb, 2001). Measuring N fixationN fixation (including transfer), calculated by the N difference method in plots 3, 10, 13 and 19 in 2000 gave values of 81.2, 87.33, 83.2 and 66.9 kg N ha-1, for 1-, 2-, 3- and 4-year-old leys, respectively. N transfer values by this method were 23.5, 4, -8 and 27.7 kg N ha-1 for 1-, 2-, 3- and 4-year-old leys. N fixation excluding transfer was therefore 57.7, 83.3, 83.2 and 39.2 for 1-, 2-, 3- and 4-year-old leys. The equivalent values estimated by 15N dilution were 53.1, 75.6, 76.1 and 31.4. The results from the two methods were therefore very closely correlated, confirming the findings of Høgh-Jensen and Kristensen (1995). The N-difference method gave consistently higher values, but the difference was not significant. N transfer estimated by the N difference method was not correlated with isotope method a) and only weakly correlated with isotope method b). None of these methods of estimating N transfer provided reliable results unless the grass monoculture plots were in close proximity to the grass-clover mixtures.
Values for N fixation at Tulloch and Woodside are relatively low compared to estimates from other sites (Boller and Nosberger, 1987, Høgh-Jensen & Schjoerring, 1997). This may have been caused by the relatively short growing season at Tulloch and Woodside.
pNdfa was higher than many estimates in the literature for leys with little of no applied N (Høgh-Jensen & Schjoerring, 1997, Boller and Nosberger, 1987, McNeill & Wood, 1990, Ledgard et al., 1990, Ledgard et al., 1996, Goodman, 1988, Heichel & Henjum, 1991, Jorgensen et al. 1999, Mallarino et al. 1990, Ledgard et al. 1987, Wheeler et al. 1997).
In this study, N fixation in clover herbage has been measured directly. This value is closely related to the yield of clover and grass obtained from monthly harvests, and cutting frequency can affect yield and N fixation (Pedersen and Moller, 1976). The experimental plots were cut once or twice a year, and were treated with manure, unlike the experimental subplots. This is one possible source of error in the calculations. Monthly cuts are also unlikely to reflect the yield of herbage produced by a grazed ley. Grazing animals trample grass and clover, and graze selectively. They also transfer fixed N from clover to grass in dung and urine. None of these factors affect the experimental subplots. The amount of grass and clover ingested by grazing animals is difficult to measure or estimate accurately. The N fixation values obtained from the experimental subplots can only be taken as an estimate of N fixation in the surrounding plots.
Time constraints prevented direct measurement of fixed N in roots, so this has been estimated, assuming a constant root/stolon:shoot ratio in clover. In reality the ratio of root/stolon:shoot can vary because of factors such as soil nutrients (Jorgensen and Ledgard, 1997, Ryle et al. 1981a). N transfer has been calculated by several methods, all of which are subject to large errors. N transfer is a measure of the amount of N transferred from clover to grass during the growing season. A number of mechanisms have been proposed for this, including mycorrhizal networks (Haystead et al., 1988), but the simplest explanation is that senescing leaves, roots and nodules decay and release N which is taken up by the grass. If this is the case then it is unlikely that all of the N present in the senescing leaves, roots and nodules will be taken up by the grass over the course of a single growing season. Therefore, in addition to fixed N in clover herbage and roots, and fixed N transferred from clover to grass; N fixation estimates must also include fixed N transferred to soil, known as rhizodeposition (Hogh-Jensen and Kristensen, 1995). Høgh-Jensen and Schjoerring (2001), estimate that rhizodeposited N from legumes may be as high as 50% of the fixed N in herbage. This figure was obtained by leaf labelling of legumes with 15N. Evidence from the preliminary root exudation experiment (in appendix) suggests that leaf labelling may dramatically increase release of N from plant roots, so these figures should be taken with caution. As there is considerable death and decay of clover plants and plant parts over winter (Wachendorf et al., 2001). Rhizodeposition has not been included in estimates of N fixation in this study.
In order to measure the total N fixation at Tulloch, including N transfer, it would be necessary to maintain a clover free control plot throughout the entire ley phase of the rotation, and compare the yields of grass to those in mixed clover grass plots. This would create practical problems on an organic farm, as the plot would very likely be invaded by clover, and this could only be controlled by hand weeding in an organic system.
During the growing season, the quantity of mineral N (ammonium-N + nitrate-N) in the top 150 mm of soil was generally similar to the quantity of soil derived N in herbage. Levels of mineral N were relatively low throughout the growing season. This confirms the observation of Granstedt (1992), that organic leys are efficient at utilising available mineral N. In the 2-year-old leys grass yield and N concentration were very low suggesting that the grass plants were deficient in N. However, this situation rapidly changed, and by the following year (3-year-old leys), grass N yields and N concentrations were at their highest.
Soil nitrate-N was increased by grazing, confirming the findings of Ledgard (2001). Variation of nitrate-N between plots was greater in grazed plots than in cut plots (see appendix). This could be a result of the rotational grazing process, which insured that on any sampling date, some of the plots would have been grazed more recently than others. Antil et al. (2001) found that mineral N underneath dung patches in grass-clover swards one month after deposition was on average 25 kg N ha-1, and in non-dunged areas mineral N was generally less than 10 kg N ha-1. This is similar to the range of values seen in this study. However the study of Antil et al. (2001) did not observe increases in mineral N in autumn on the scale seen in this study. Soil nitrate-N at Tulloch was highest in the autumn and winter in 3-year old leys, and this may have been related to the high yields of clover in the preceding growing season. Chalmers et al. (2001) and Eriksen et al. (1999) also observed enhanced mineralisation in grass-clover leys in Autumn, This was observed even in the absence of any manure or fertiliser applications, strongly suggesting that the enhanced mineralisation was due to clover rhizodeposition.
Elevated soil N under clover patches was observed at Tulloch. This could have been due to lower uptake of N by clover than grass (known as the N sparing effect, Senaratne and Hardarson, 1988), release of nitrogen from clover roots and leaf litter or enhanced mineralisation of soil organic matter by clover. The N sparing hypothesis assumes that clover suppresses grass, and prevents it from taking up nitrogen. The 15N dilution experiment showed that this was not the case: clover enhanced the yield of grass growing in close proximity to it, therefore clover patches would have more grass than grass only patches with similar soil nutrient levels.
The effects of cutting and grazing on N fixation could not be measured in this study, with any certainty, because all of the experimental subplots received the same management: monthly cutting, with no manure applied. However, there were some indications that grazing of leys early in the year increased grass yield and N concentration and reduced pNdfa, confirming the findings of Vinther (1998) and Ledgard (2001).
It was expected that the 66% ley rotation would have higher soil fertility, because it had a longer fertility building ley phase. Higher ammonium and K was observed, but there were no differences in the total soluble N levels or in soil organic matter between the two rotations. The higher ammonium levels may have been responsible for the slightly higher grass yields and SDN observed in the 66% ley rotation.
The assumption that an extra year of ley will increase soil fertility may not be correct. In 2000, the ratio of grass:clover in 4-year-old leys was very high, and consequently, cutting of the ley for silage would remove more N from the soil than the ley fixed. Much of the N present in the silage would ultimately be returned to the soil in the form of manure, but composting of manure can result in considerable losses of N (Granstedt 1995, Sommer, 2001). Granstedt (1992) observed a rapid decline in N fixation with age of ley, and noted that the final year of ley had a high grass:clover ratio and would therefore deplete soil N. Removal of N in silage and the potential losses of N from manure must be balanced against the reduced leaching losses of N resulting from ploughing the ley when it has a low clover content: mineralisation of N is likely to be slower following the ploughing of a grass rich ley than following a clover rich ley. N leaching losses following ploughing of a grass-clover ley may be almost twice as high (80 kg N ha-1 year-1) as leaching losses from a grass-only ley (42 kg N ha-1 year-1, Goulding, 2000). In 2000, 4-year-old leys had less than half as much clover N yield as 3-year-old leys. Grass yields were similar in 3 and 4-year-old leys, but grass N concentration was lower in 4-year-old leys. This suggests that leaching losses would be around 50% lower following ploughing of 4-year-old leys than after ploughing of 3-year-old leys. This could mean that losses of N would be reduced following ploughing by approximately 20 kg N ha-1, if 4-year-old rather than 3-year-old leys were ploughed. The benefits of a fourth year of ley to subsequent crops may not be very great. The decision to include a fourth year of ley may depend on the relative values of arable crops and silage in the system.
Frame and Newbould (1986) state that "once a threshold quantity of organic matter has been built up which mineralises sufficient nitrogen to support moderate levels of grass growth, it is extremely difficult to sustain the presence of white clover…..The farmer should then concentrate on the use of high yielding grasses using additional fertiliser N, while this is available and its use is economically justified". The results of this study do not bear this out. Tulloch has a high fertility soil, rich in organic matter, but clover does not, on the whole seem to be suffering from the effects of excess soil N, as pNdfa is generally high, as is the clover content of the swards.
Soil organic matter in leys at Tulloch was slightly higher in 2000 (9.81%) than in 1992 (8.2%, SAC data), but there is no clear evidence of a long-term trend. Depletion of soil N and other nutrients would be indicated by a decline in soil N (Granstedt, 1992, Korsaeth et al., 2003).
The role of SON in the system was not clear. SON was correlated with grass N concentration, but had no discernible effect on clover. SON did not appear to follow any clear seasonal pattern, although there was variation between sampling dates. Some of this SON may result from rhizodeposition by clover, as SON was higher under clover microsites on some sampling dates. The ability of clover and grass to use SON is not well understood.
The ratio of dry weight to fresh weight was slightly higher in 1-year-old leys than in older leys in 2001. The reasons for this are not clear, and it has not been observed in any other studies in the literature. Flowering rate declined with age of ley, as did tap root numbers. Both flowering rate and tap roots have been linked to clover persistence (Williams, 1987). Tap-roots may senesce earlier when P and K are deficient (Westbrook and Tesar, 1955). There is ample evidence that white clover stolon growth and branching are suppressed by shading from grass (De Kroons and Hutchings, 1995, Heraut-Bron et al. 2001, Marriott et al. 1997a, Faurie et al. 1996, Thompson, 1993).
Elevated soil N was associated with elevated clover N concentration in these experiments. Most studies of this type have found no effect of soil N on N concentration of clover (Herrmann et al., 2001, Ledgard et al., 2001, Høgh-Jensen and Schjoerring, 1994) and in one instance, elevated soil N has been observed to reduce clover N concentration (Elgersma et al. 2000). The correlation of clover N concentration with soil N observed in this study could not simply have been due to uptake of soil N, because there was not a close, linear correlation between clover N concentration and pNdfa (Figure 5.6.2). In the years and ages of ley studied, high clover N concentration and low pNdfa seemed to be an indicator of low N fixation in the following year (Figure 8.12.1). Clover N concentration was negatively correlated with soil P and pNdfa was negatively correlated with soil N.
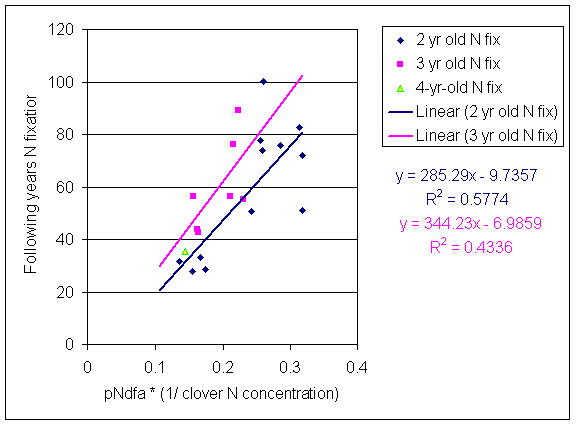
Figure .1 Correlation between average annual pNdfa x (1/average annual clover N concentration) and total N fixation in the following year. Each point is the mean of three subplots in a plot.
It can be hypothesised from the available evidence that shading by grass, causes clover to divert energy into leaf and petiole production, at the expense of stolons, in order to successfully compete with grass for light (De Kroons and Hutchings, 1995, Faurie et al. 1996, Marriott et al. 1997a, Markuvitz and Turkington, 2000). As leaves and petioles have higher N concentration than stolons (Warembourg et al. 1997, Høgh-Jensen et al. 2001), this could be partly responsible for the changes in clover N concentration observed between different ages of ley and year. This would explain why high clover N concentration often precedes low clover yields (Figure 8.12.1). It is not clear why other studies have not observed changes in clover N concentration in response to N fertilisation.
It is also possible that clover changes its reproductive strategy in response to environmental conditions. White clover can persist in one location as a tap-rooted perennial, lose its tap-root and colonise new microsites as a clonal plant using its stolons, and also vary the type and quantity of seed produced. Hard seed is produced in response to drought (Thomas, 1987), but it could also be a response to a number of unfavourable environmental conditions.