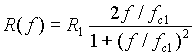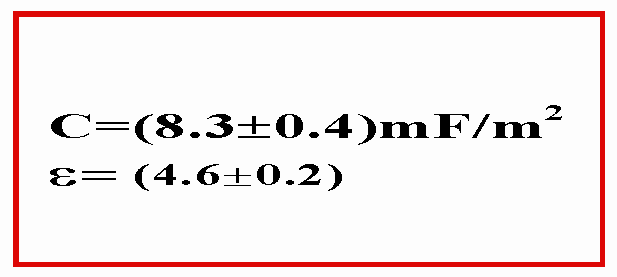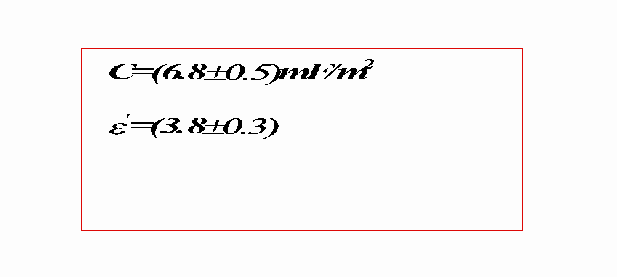
ELETTROROTAZIONE
Introduzione
La tecnica dell’elettrorotazione consente essenzialmente di studiare le proprietà dielettriche delle membrane cellulari ( capacità, permittività, conducibilità ecc. ). Scopo della nostra esperienza è quello di misurare la capacità e la permittività di membrana di cellule 3T6 (fibroblasti di topo); si è inoltre proceduti ad una stima del raggio di tali cellule e si è apportata una significativa evoluzione all’apparato sperimentale inizialmente presente. Introduciamo l’argomento in esame facendo delle considerazioni fisiche introduttive.
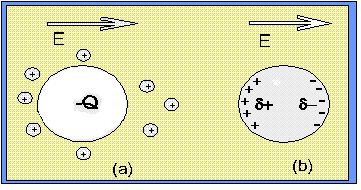
Quando andiamo a sospendere in un elettrolita una particella (o una cellula) trasportante una carica elettrica, essa attrarrà attorno a sé ioni di carica elettrica opposta, formando un doppio strato elettrico. Nella figura di sinistra (a) seguente viene mostrato che la simmetria del doppio strato può essere alterata dalla presenza di un campo elettrico esterno applicato. Di solito le proprietà dielettriche della particella (cioè la conducibilità e la costante dielettrica ) differiscono da quelle della sospensione: come risultato di ciò l’azione del campo elettrico applicato sarà quella di indurre delle cariche elettriche alla superficie di separazione tra particella e sospensione (mezzo esterno). Quest’ultimo è quello che viene detto effetto Maxwell-Wagner ed è evidenziato nella seguente figura di destra (b).
La distorsione
del doppio strato elettrico e la creazione di cariche interfacciali per
effetto Maxwell-Wagner si prestano alla creazione di un momento di dipolo
indotto della particella. Le ampiezze di questi dipoli indotti sono molto più
grandi rispetto a quelle associate alle specie molecolari perché, sebbene
la quantità di carica coinvolta non risulta essere molto grande, la distanza
tra due cariche di segno opposto è considerevole. Come mostrato nella
figura (b) di cui sopra il momento di dipolo indotto originato da due cariche
indotte ![]() e
e ![]() localizzate a distanza rispettivamente r+ e r- è
dato da :
localizzate a distanza rispettivamente r+ e r- è
dato da :
dove r è il raggio delle particelle.
La teoria
Se il campo elettrico applicato è un campo alternato (a.c.), la quantità di carica indotta su ogni interfaccia dipende dalla frequenza di tale campo. Le forze coulombiane indurranno sulle cellule polarizzate una traslazione (dielettroforesi : qui la velocità traslazionale delle cellule dipende dalla polarizzazione elettrica e soprattutto dal grado di disomogeneità del campo elettrico a.c. generato da coppie di elettrodi diversi) od una rotazione (elettrorotazione E.R.).
Per indurre la rotazione delle cellule si espongono le medesime ad un campo elettrico ruotante ottenuto pilotando una cella (a 4 elettrodi vedi dopo) con 4 segnali sfasati di 900. Le cellule sottoposte a tale campo iniziano a ruotare con una frequenza molto più bassa di quella del campo applicato.
Il momento
di dipolo indotto della particella ruota con la stessa frequenza del campo elettrico
ma sfasato rispetto ad esso. Questo sfasamento è identico all’angolo
di fase ![]() tra
tra ![]() e
e ![]() .
.
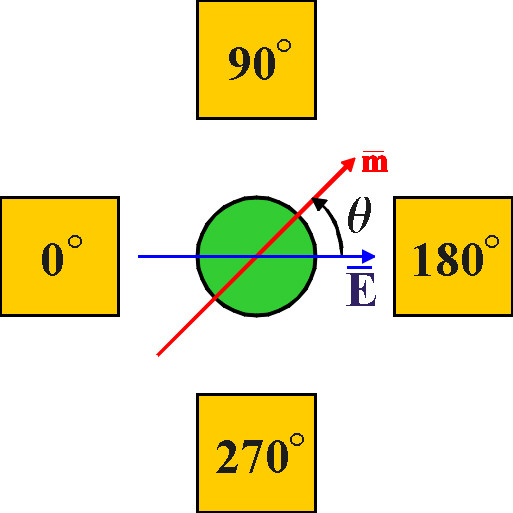
Fig. c
Il momento
di dipolo ![]() di una sfera
dielettrica omogenea immersa in un mezzo dielettrico esterno può scriversi
come
di una sfera
dielettrica omogenea immersa in un mezzo dielettrico esterno può scriversi
come
![]() 0000000(1)
0000000(1)
dove:
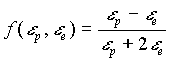 0000000(2)
0000000(2)
è detto fattore di Clausius-Mossotti
![]() e
e ![]() sono rispettivamente le costanti dielettriche complesse (dipendenti dalla frequenza
sono rispettivamente le costanti dielettriche complesse (dipendenti dalla frequenza
![]() )
della cellula e del mezzo esterno (
)
della cellula e del mezzo esterno (![]() ).
).
Quando![]() >
> ![]() allora
allora ![]() >0 e il momento di dipolo è allineato
al campo
>0 e il momento di dipolo è allineato
al campo ![]() e viceversa
se
e viceversa
se ![]() <
< ![]() .
.
Notare che f(ep ,ee) è limitata cioè
1![]()
![]()
![]() -1/2 (
-1/2 (![]() =1
per
=1
per ![]() =0
,
=0
, ![]() =-1/2 per
=-1/2 per ![]() =0),
=0),
cosicchè
l’ampiezza di ![]() ,dipendente
dal volume della particella, dall’ampiezza del campo elettrico applicato e dalla
differenza delle polarizzabilità della cellula e del mezzo esterno, è
anch’essa limitata.
,dipendente
dal volume della particella, dall’ampiezza del campo elettrico applicato e dalla
differenza delle polarizzabilità della cellula e del mezzo esterno, è
anch’essa limitata.
L’interazione
tra ![]() e
e ![]() ,
per ogni data frequenza , genera un momento meccanico
,
per ogni data frequenza , genera un momento meccanico ![]() che induce le cellule a ruotare (seppure come detto a frequenza più lenta
di quella del campo). Nella determinazione delle proprietà dielettriche
la dipendenza dalla frequenza della direzione e della velocità di rotazione
può esprimersi mediante un modello lineare (vero per rotazioni lente)
che consente di esprimere
che induce le cellule a ruotare (seppure come detto a frequenza più lenta
di quella del campo). Nella determinazione delle proprietà dielettriche
la dipendenza dalla frequenza della direzione e della velocità di rotazione
può esprimersi mediante un modello lineare (vero per rotazioni lente)
che consente di esprimere ![]() come
come
![]() 0000000(3)
0000000(3)
Nell’elettrorotazione
si può misurare la dipendenza dalla frequenza della parte immaginaria
di ![]() . Poiché
. Poiché ![]()
![]()
![]() allora
allora ![]()
![]() E2 e risulta anche che
E2 e risulta anche che
![]() 0000000(4)
0000000(4)
per cui il momento meccanico medio di una cellula sferica di raggio r può scriversi
![]() 0000000(5)
0000000(5)
Il simbolo
Im( ) indica che solo la parte immaginaria del momento di dipolo da contributo
al momento meccanico : ciò significa che tale componente immaginaria
determina l’ampiezza e il verso della rotazione delle cellule. Allora se Im(![]() )
è positiva il momento meccanico sarà negativo causando la rotazione
delle cellule nel verso opposto a quello del campo rotante.
)
è positiva il momento meccanico sarà negativo causando la rotazione
delle cellule nel verso opposto a quello del campo rotante.
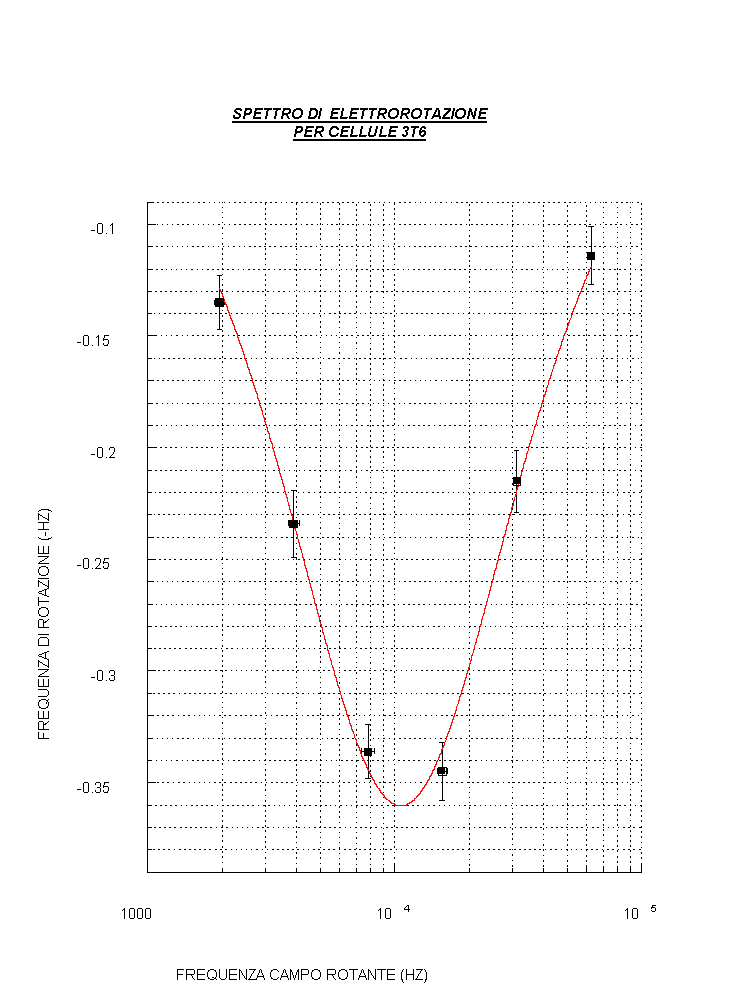
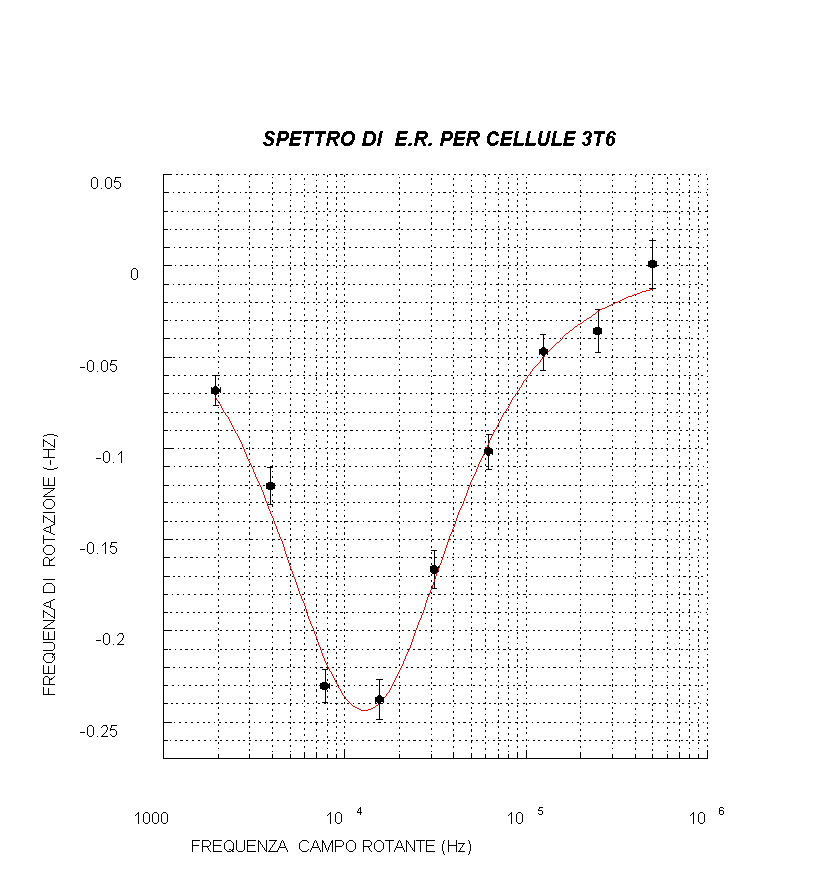
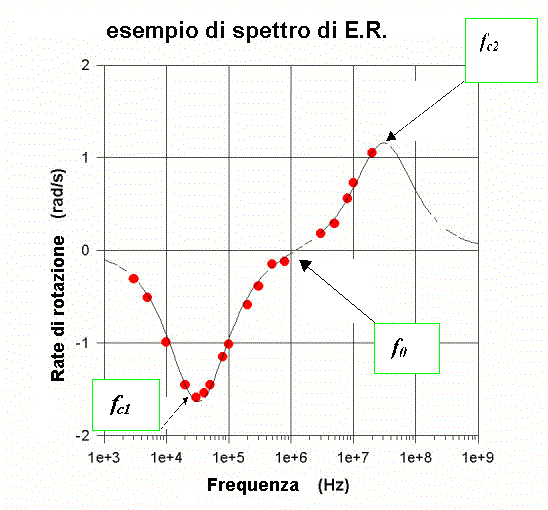
Lo spettro di rotazione e le due dispersioni caratteristiche dell’E.R.
Sperimentalmente uno spettro di elettrorotazione, ottenuto rappresentando la frequenza di rotazione delle cellule (eventualmente normalizzata al quadrato dell'ampiezza del campo) in funzione della frequenza del campo esterno applicato (vedi fig. d), presenta 2 dispersioni caratterizzate da altrettanti picchi (in corrispondenza dei massimi della frequenza di rotazione) che evidenziano: il primo, una rotazione delle cellule in verso opposto al campo (e ciò si vedrà avvenire a bassa frequenza), il secondo (ad alta frequenza) una rotazione concorde a quella del campo.
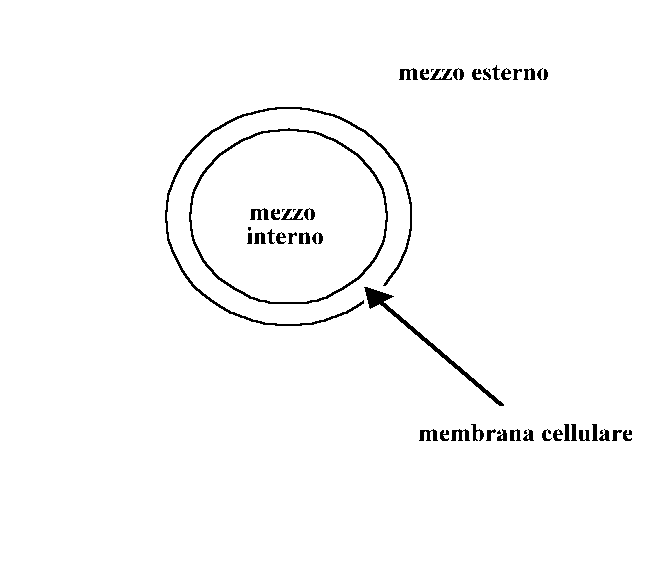
Per interpretare lo spettro di elettrorotazione schematizziamo le cellule come nel modello a single shell , cioè come sfere il cui interno può essere rappresentato da un elettrolita più o meno omogeneo (mezzo interno) racchiuso da una membrana (strato isolante) e il tutto immerso in un mezzo esterno (soluzione tampone) come in fig. g .
Risolvendo l’equazione di Laplace con le condizioni al contorno valide per tale modello a tre dielettrici, si possono esprimere due frequenze caratteristiche in funzione dei parametri dielettrici: la frequenza del primo picco (fc1) , determinata dalle proprietà della membrana ( in particolare fc1 permette, una volta stimata, la determinazione della permittività della membrana plasmatica che è una espressione strutturale cioè dipende dalla disposizione delle proteine intramembranose nel doppio strato lipidico) e la frequenza del secondo picco (fc2) , legata alle proprietà del mezzo interno (vedi fig. d).
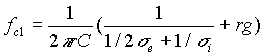 0000000(6)
0000000(6)
![]() 00000 (7)
00000 (7)
dove C
e g indicano rispettivamente la capacità specifica e la conducibilità
di membrana, mentre![]() e
e ![]() sono le conducibilità rispettivamente del mezzo interno e del mezzo esterno.
sono le conducibilità rispettivamente del mezzo interno e del mezzo esterno.
E’ possibile esprimere in base a quanto suddetto anche una terza frequenza f0 in cui si ha l’inversione del verso di rotazione della cellula (vedi fig. d).
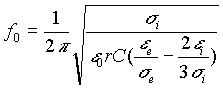
In prima
approssimazione nella (6) si può trascurare il prodotto rg ed
il termine 1/ ![]() rispetto a 1/
rispetto a 1/ ![]() in quanto
in quanto ![]() >>
>> ![]() per cui la (4) diventa
per cui la (4) diventa
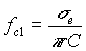 000000(8)
000000(8)
Dalla (8)
misurando![]() si ricava la capacità per unità
di superficie C (una volta stimato il raggio cellulare)
si ricava la capacità per unità
di superficie C (una volta stimato il raggio cellulare)
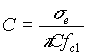 000000(9)
000000(9)
e infine
nota quest’ultima quantità, e noto lo spessore di membrana d =50*10
–10 m (come da letteratura) è possibile stimare la permittività
di membrana ![]()
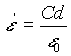 000000(10)
000000(10)
Abbiamo accennato che applicando un campo elettrico rotante su una sospensione cellulare, si induce all’interfaccia membrana-tampone un momento di dipolo per polarizzazione interfacciale (in quanto c’è una forte differenza di conducibilità e costante dielettrica tra tampone e membrana).
A frequenza molto bassa il momento di dipolo viene istantaneamente indotto, in fase con il campo elettrico, sulla cellula (il campo elettrico gira e man mano che ruota, sulla cellula si instaura il suddetto momento di dipolo sempre allineato con il campo). Ricordiamo infatti che un dipolo allineato con il campo non subisce alcun momento meccanico. A frequenza molto alta non si induce momento di dipolo (e dunque anche qui non succede niente). Tra questi due estremi c’è una situazione in cui il momento di dipolo viene indotto però sfasato rispetto al campo elettrico rotante: in ogni istante abbiamo che il campo elettrico e il momento di dipolo indotto non sono allineati per via del fenomeno del rilassamento.
Analizziamo più in dettaglio quanto detto.
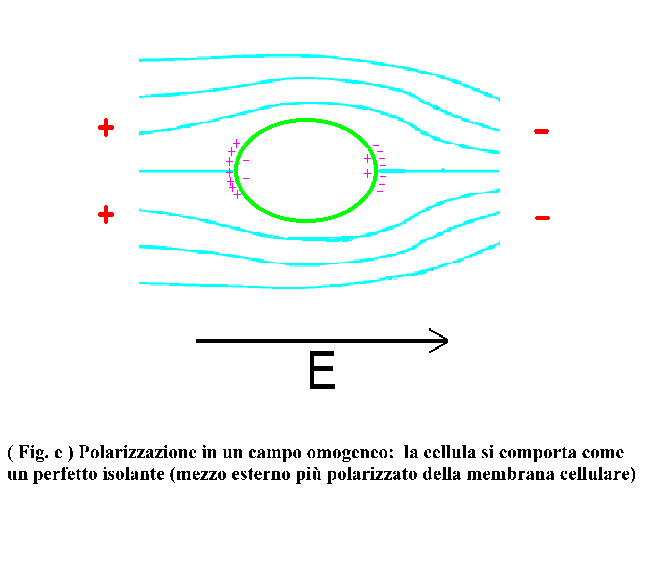
I) La prima dispersione (in genere per frequenze < 1 MHZ)
Consideriamo un campo alternato a frequenza sufficientemente bassa, in modo tale che l’impedenza di membrana sia molto più grande della resistenza del mezzo interno e del mezzo esterno. In tali condizioni la corrente associata a tale campo elettrico è forzata a scorrere intorno alla cella, provocando un accumulo di cariche affacciate agli elettrodi (vedi fig. e)
Avendo
il citoplasma una grossa conducibilità il campo elettrico all’interno
della cellula è praticamente nullo: la differenza di potenziale è
localizzata sullo strato della membrana. Il momento di dipolo indotto è
come detto, dovuto alle cariche poste sull’interfaccia tra la membrana e il
mezzo esterno e la sua orientazione è parallela al campo esterno. Dato
che il processo di carica della membrana ha una sua costante di tempo, dipendente
dalle proprietà dielettriche della cellula e del mezzo esterno, si può
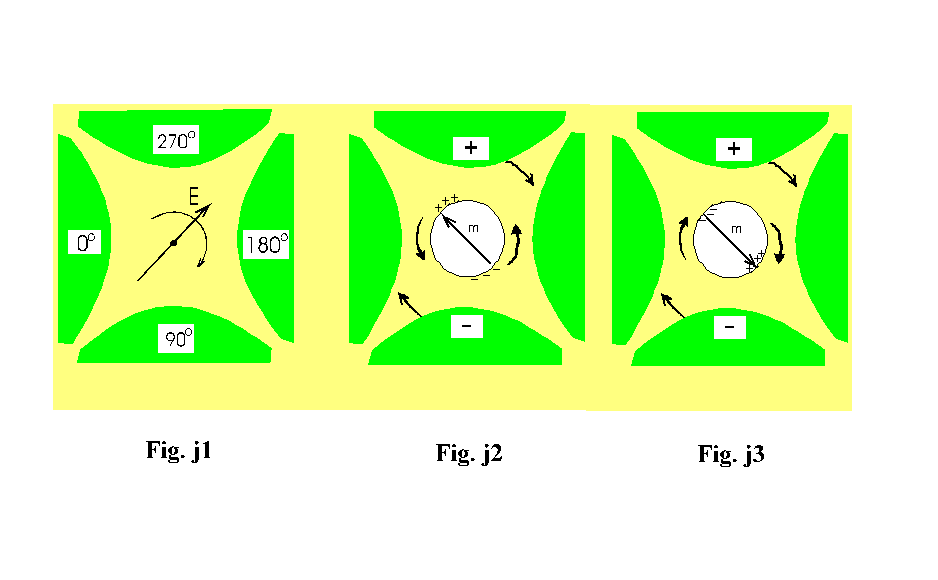
avere uno sfasamento tra il campo applicato e il momento di dipolo indotto (Fig.
j1 j2). Tale sfasamento, in un campo rotante, coincide con l’angolo di fase
spaziale ![]() (
(![]() ruoterà con la
stessa frequenza di
ruoterà con la
stessa frequenza di![]() e
con una certa differenza di fase che corrisponde all’angolo tra la direzione
dei due vettori sopra menzionati).
e
con una certa differenza di fase che corrisponde all’angolo tra la direzione
dei due vettori sopra menzionati).
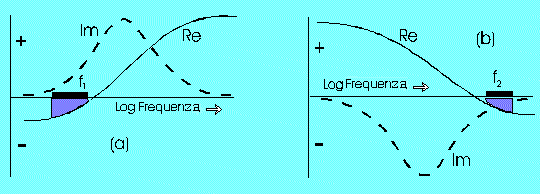
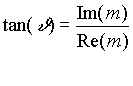
In Fig.
f (a) si vede che la parte immaginaria del momento di dipolo indotto è
positiva per angoli compresi tra 0 e 180 gradi in modo che per la (4) le cellule
ruotano in verso contrario al campo. Quindi ricordando che un momento di dipolo
![]() non allineato con il
campo elettrico E genera un momento meccanico m
nel nostro caso avremo che quest’ultimo, per l’orientazione
del dipolo, spingerà la cellula a ruotare (lentamente) , come detto in
direzione opposta al campo (questo fenomeno si verifica sperimentalmente per
frequenze < 1MHz). La velocità di rotazione dipende dalla frequenza
del campo ed è massima quando la frequenza del campo è uguale
alla frequenza di rilassamento della polarizzazione interfacciale.
non allineato con il
campo elettrico E genera un momento meccanico m
nel nostro caso avremo che quest’ultimo, per l’orientazione
del dipolo, spingerà la cellula a ruotare (lentamente) , come detto in
direzione opposta al campo (questo fenomeno si verifica sperimentalmente per
frequenze < 1MHz). La velocità di rotazione dipende dalla frequenza
del campo ed è massima quando la frequenza del campo è uguale
alla frequenza di rilassamento della polarizzazione interfacciale.
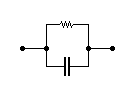
II) La seconda dispersione (> 1 MHz)
Schematizzando la membrana cellulare come un circuito RC parallelo
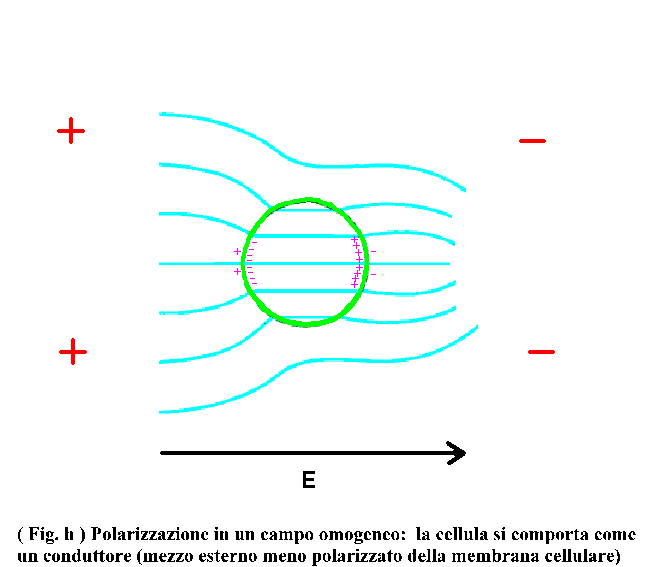
A frequenze sufficientemente elevate (ma non troppo in quanto le cariche devono riuscire a seguire le oscillazioni del campo anche se sfasate) l’isolamento della membrana è bypassato per via capacitiva (le linee di forza del campo riescono ad attraversare la membrana e vanno ad interessare il mezzo interno caratterizzato da una elevata conducibilità). Così le cariche positive sono accumulate dentro il citoplasma di fronte all’elettrodo negativo (nel caso precedente, invece esse si accumulavano sulla superficie esterna della membrana di fronte all’elettrodo positivo); viceversa si comportano gli ioni negativi. Tutto ciò porta ad un orientazione del momento di dipolo indotto antiparallela al campo esterno (vedi Fig. h)
In
Fig. f (b) si vede che la parte immaginaria del momento di dipolo indotto è
negativa per angoli compresi tra 0 e 180 gradi in modo che per la (4) le cellule
ruotano in verso concorde al campo. Quindi ricordando che un momento di dipolo
![]() non allineato con il
campo elettrico E genera un momento meccanico m
: nel nostro caso avremo che quest’ultimo, per
l’orientazione del dipolo, spingerà la cellula a ruotare (lentamente)
, come detto nella stessa direzione del campo (passando dalla membrana al mezzo
interno la polarizzazione è in verso contrario alla precedente dispersione)
come in Fig. j3.
non allineato con il
campo elettrico E genera un momento meccanico m
: nel nostro caso avremo che quest’ultimo, per
l’orientazione del dipolo, spingerà la cellula a ruotare (lentamente)
, come detto nella stessa direzione del campo (passando dalla membrana al mezzo
interno la polarizzazione è in verso contrario alla precedente dispersione)
come in Fig. j3.
Strumentazione a disposizione
Qui sotto (Fig.1) si può notare l'apparato sperimentale usato per effettuare le misure di elettrorotazione. Si possono individuare:
- microscopio Zeiss
- cella porta campione
- generatore di campo rotante
- una telecamera
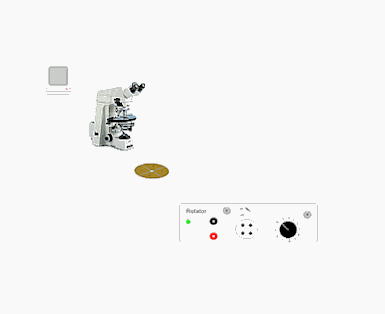
Fig 1
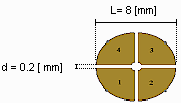
La cella di misura è costituita da quattro elettrodi elettricamente isolati l’uno dall’altro e montati su di
un vetrino da microscopio nel cui centro è presente una cavità di diametro pari a 2.5 mm.
Fig 2 (cella di misura )
In questa cavità viene posto il campione: una goccia della sospensione di cellule ( fibroblasti di topo 3T6) di volume pari a circa 10-20 ml; il tutto verrà successivamente ricoperto da un vetrino posto con lo scopo di favorire una migliore visualizzazione da parte dell’operatore.
Il circuito è alimentato con un generatore di campo rotante che invia ai quattro elettrodi in sequenza, dei segnali ad onda quadra sfasati di 90°. Grazie alla presenza di un divisore, la frequenza del segnale può essere variata tra 240 Hz e 500 KHz in 12 step.
Fig 3 (generatore di campo rotante )
Di seguito vengono riportate delle immagini a fluorescenza riguardanti la distribuzione del campo nella zona centrale della cella di misura.
Preliminari di misura
Le misure vengono effettuate dalla rotazione di cellule 3T6 sospese in una soluzione di saccarosio (300 mM). La soluzione presenta una conducibiltà misurata di
= ( 2.36 ± 0.01) 10-3 S/m
Dal punto di vista sperimentale si è individuata la più appropriata tecnica di rilevamento del periodo di rotazione delle cellule: la più indicata era quella di ottenerlo come risultato di una media su due rotazioni complete, osservate tramite telecamera CCD posta sulla sommità del microscopio; le misure venivano condotte con obbiettivi da 16 e 33 ingrandimenti ( il 33 quando disponibile ).
Il secondo problema era relativo alle dimensioni particolarmente estese della celletta rispetto a quelle della coltura: in prossimità degli elettrodi, il movimento predominante era quello traslatorio mentre nella parte più interna del pozzetto si assisteva ad una effettiva rotazione: questo perché il campo non era uniforme su tutto il volume ma solamente in una piccola porzione di spazio nel centro.
In virtù di quanto detto sopra, le cellule, durante la misura, non rimangono sempre su di uno stesso piano focale, quindi oltre che regolare la messa a fuoco, bisogna anche inseguirle per tutto il pozzetto (escludendo dalla misura quelle più profonde a diretto contatto con il vetrino).
La misura è ulteriormente complicata dalla eventuale presenza nella soluzione di aggregati cellulari,
(cellule addensate tra di loro): campioni di questo genere non sono utilizzabili in quanto hanno una consistente inerzia al moto. È quindi necessario poter visualizzare una cellula isolata seguendone gli spostamenti ( rotazioni ), quando essa è sottoposta al campo esterno.
Dal punto di vista elettrico abbiamo verificato, come al variare della frequenza l’ampiezza del segnale in uscita ai quattro elettrodi non variasse: in virtù di ciò si è fissata una tensione di riferimento intorno ai 12 V ( questo anche allo scopo di salvaguardare la componentistica elettronica ).
Abbiamo verificato, grazie all’ausilio di un’oscilloscopio, i valori dei periodi rilevati alle varie (12) posizioni della manopola sul generatore di campo rotante ( ROTATOR ) tensione qui di seguito riportate:
|
Posizione Manopola |
Periodo T± DT [s] |
Frequenza n ± D n [Hz] |
|
1 |
( 4.2 ± 0.1 ) 10-3 |
240 ± 5 |
|
2 |
( 2.08 ± 0.02 ) 10-3 |
480 ± 5 |
|
3 |
( 1.03 ± 0.01 ) 10-3 |
970 ± 10 |
|
4 |
( 5.1 ± 0.1 ) 10-4 |
1950 ± 50 |
|
5 |
( 2.56 ± 0.05 ) 10-4 |
3900 ± 100 |
|
6 |
( 1.29 ± 0.02 ) 10-4 |
7810 ± 120 |
|
7 |
( 6.4 ± 0.1 ) 10-5 |
15600 ± 240 |
|
8 |
( 3.21 ± 0.05 ) 10-5 |
31200 ± 500 |
|
9 |
( 1.60 ± 0.02 ) 10-5 |
62400 ± 800 |
|
10 |
( 8.0 ± 0.2 ) 10-6 |
125000 ± 2200 |
|
11 |
( 4.0 ± 0.1 ) 10-6 |
250000 ± 3100 |
|
12 |
( 2.00 ± 0.02 ) 10-6 |
500000 ± 5000 |
|
- |
( 1.82 ± 0.05 ) 10-4 |
5500 ± 150 |
|
- |
( 1.00 ± 0.02 ) 10-4 |
10000 ± 200 |
|
- |
( 8.3 ± 0.2 ) 10-5 |
12000 ± 300 |
|
- |
( 5.0 ± 0.1 ) 10-5 |
20000 ± 400 |
|
- |
( 4.00 ± 0.05 ) 10-5 |
25000 ± 300 |
|
- |
( 2.50 ± 0.05 ) 10-5 |
40000 ± 800 |
|
- |
( 2.00 ± 0.02 ) 10-5 |
50000 ± 1300 |
|
- |
( 1.34 ± 0.02 ) 10-5 |
75000 ± 1100 |
|
- |
( 1.00 ± 0.02 ) 10-5 |
100000 ± 2000 |
|
- |
( 6.7 ± 0.1 ) 10-6 |
150000 ± 2200 |
(per calcolare il D n si è fatto uso della propagazione degli errori statistici ).
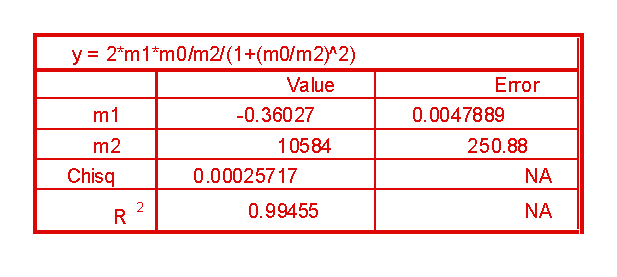
Per il fit dei dati si è usata la formula (vera in prima approssimazione per il modello a single shell ):
dove f è la frequenza del campo applicato, R(f) la frequenza di rotazione cellulare, R1 il picco di R(f) e fc1 la relativa frequenza del campo in corrispondenza di R1
La nostra misura (prova1)
|
Frequenza Campo (Hz) |
Errore associato (Hz) |
|
1950 |
100 |
|
3900 |
200 |
|
7810 |
400 |
|
15600 |
800 |
|
31200 |
1600 |
|
62400 |
3100 |
|
Misura 1 (s) |
Misura 1 ( Hz ) |
|
8.33 |
- 0.120 |
|
8.40 |
- 0.119 |
|
3.15 |
- 0.317 |
|
3.03 |
- 0.330 |
|
5.05 |
- 0.198 |
|
11.1 |
- 0.090 |
|
Misura 2 (s) |
Misura 2 ( Hz ) |
|
7.04 |
- 0.142 |
|
4.15 |
- 0.241 |
|
2.92 |
- 0.343 |
|
2.86 |
- 0.350 |
|
4.50 |
- 0.222 |
|
8.26 |
- 0.121 |
|
Misura 3 (s) |
Misura 3 ( Hz ) |
|
8.85 |
- 0.113 |
|
4.61 |
- 0.217 |
|
3.14 |
- 0.318 |
|
3.09 |
- 0.324 |
|
5.26 |
- 0.190 |
|
11.1 |
- 0.090 |
|
Misura 4 (s) |
Misura 4 ( Hz ) |
|
6.58 |
- 0.152 |
|
3.95 |
- 0.253 |
|
2.86 |
- 0.350 |
|
2.74 |
- 0.365 |
|
4.35 |
- 0.230 |
|
7.46 |
- 0.134 |
|
Misura 5 (s) |
Misura 5 ( Hz ) |
|
7.46 |
- 0.134 |
|
4.24 |
- 0.236 |
|
2.99 |
- 0.335 |
|
3.00 |
- 0.333 |
|
4.57 |
- 0.219 |
|
9.62 |
- 0.104 |
|
Misura 6 (s) |
Misura 6 ( Hz ) |
|
7.94 |
- 0.126 |
|
4.33 |
- 0.231 |
|
3.00 |
- 0.333 |
|
2.90 |
- 0.345 |
|
5.03 |
- 0.199 |
|
9.01 |
- 0.111 |
|
Misura 7 (s) |
Misura 7 ( Hz ) |
|
7.09 |
- 0.141 |
|
4.61 |
- 0.217 |
|
3.09 |
- 0.324 |
|
2.87 |
- 0.348 |
|
4.65 |
- 0.215 |
|
8.26 |
- 0.121 |
|
Misura 8 (s) |
Misura 8 ( Hz ) |
|
7.25 |
- 0.138 |
|
4.50 |
- 0.222 |
|
2.85 |
- 0.351 |
|
2.91 |
- 0.344 |
|
4.33 |
- 0.231 |
|
9.26 |
- 0.108 |
|
Misura 9 (s) |
Misura 9 ( Hz ) |
|
6.34 |
- 0.144 |
|
4.13 |
- 0.242 |
|
2.98 |
- 0.336 |
|
2.84 |
- 0.352 |
|
4.46 |
- 0.224 |
|
7.94 |
- 0.126 |
|
Misura 10 (s) |
Misura 10 (Hz) |
|
7.09 |
- 0.141 |
|
3.88 |
- 0.258 |
|
2.87 |
- 0.348 |
|
2.77 |
- 0.361 |
|
4.46 |
- 0.224 |
|
8.13 |
- 0.123 |
|
Media frequenza di rotazione (Hz) |
Deviazione Standard (Hz) |
|
- 0.135 |
0.012 |
|
- 0.234 |
0.015 |
|
- 0.336 |
0.012 |
|
- 0.345 |
0.013 |
|
- 0.215 |
0.014 |
|
- 0.114 |
0.013 |
Si noti che:
La nostra misura (prova2) (verifica della riproducibilità della misura fisica)
|
Frequenza Campo (Hz) |
Errore associato (Hz) |
|
1950 |
100 |
|
3900 |
200 |
|
7810 |
400 |
|
15600 |
800 |
|
31200 |
1600 |
|
62400 |
3100 |
|
125000 |
6200 |
|
250000 |
12400 |
|
500000 |
24800 |
|
Misura 1 (s) |
Misura 1 ( Hz ) |
|
13.51 |
-0.074 |
|
9.03 |
-0.111 |
|
4.54 |
-0.220 |
|
4.31 |
-0.232 |
|
6.33 |
-0.158 |
|
8.13 |
-0.123 |
|
17.01 |
-0.059 |
|
78.74 |
-0.013 |
|
77.50 |
+0.013 |
|
Misura 2 (s) |
Misura 2 ( Hz ) |
|
13.72 |
-0.073 |
|
9.14 |
-0.109 |
|
4.31 |
-0.232 |
|
4.34 |
-0.230 |
|
5.97 |
-0.168 |
|
10.16 |
-0.098 |
|
25.97 |
-0.038 |
|
23.15 |
-0.043 |
|
93.46 |
-0.011 |
|
Misura 3 (s) |
Misura 3 ( Hz ) |
|
14.74 |
-0.068 |
|
8.79 |
-0.114 |
|
4.08 |
-0.245 |
|
3.84 |
-0.260 |
|
5.81 |
-0.172 |
|
10.02 |
-0.100 |
|
35.59 |
-0.028 |
|
27.84 |
-0.036 |
|
65.40 |
+0.015 |
|
Misura 4 (s) |
Misura 4 ( Hz ) |
|
19.92 |
-0.050 |
|
8.62 |
-0.116 |
|
4.33 |
-0.231 |
|
4.39 |
-0.228 |
|
5.44 |
-0.184 |
|
9.80 |
-0.102 |
|
20.44 |
-0.049 |
|
33.47 |
-0.030 |
|
73.53 |
-0.014 |
|
Misura 5 (s) |
Misura 5 ( Hz ) |
|
13.15 |
-0.076 |
|
8.09 |
-0.124 |
|
4.59 |
-0.218 |
|
4.11 |
-0.243 |
|
5.90 |
-0.169 |
|
9.75 |
-0.103 |
|
17.18 |
-0.058 |
|
26.01 |
-0.038 |
|
106.38 |
-0.009 |
|
Misura 6 (s) |
Misura 6 ( Hz ) |
|
13.59 |
-0.074 |
|
7.19 |
-0.139 |
|
4.12 |
-0.243 |
|
4.18 |
-0.239 |
|
6.00 |
-0.167 |
|
9.40 |
-0.106 |
|
17.58 |
-0.057 |
|
25.94 |
-0.039 |
|
94.34 |
-0.011 |
|
Misura 7 (s) |
Misura 7 ( Hz ) |
|
14.02 |
-0.071 |
|
8.64 |
-0.116 |
|
4.51 |
-0.222 |
|
4.22 |
-0.237 |
|
6.90 |
-0.145 |
|
11.91 |
-0.084 |
|
20.69 |
-0.048 |
|
17.24 |
-0.058 |
|
72.50 |
+0.014 |
|
Misura 8 (s) |
Misura 8 ( Hz ) |
|
14.92 |
-0.067 |
|
7.35 |
-0.136 |
|
4.31 |
-0.231 |
|
4.40 |
-0.227 |
|
5.71 |
-0.175 |
|
9.87 |
-0.101 |
|
20.55 |
-0.049 |
|
27.55 |
-0.036 |
|
80.60 |
+0.012 |
|
Misura 9 (s) |
Misura 9 ( Hz ) |
|
17.07 |
-0.059 |
|
8.00 |
-0.125 |
|
4.42 |
-0.226 |
|
4.34 |
-0.230 |
|
6.20 |
-0.161 |
|
10.06 |
-0.099 |
|
25.78 |
-0.039 |
|
39.37 |
-0.025 |
|
78.13 |
-0.013 |
|
Misura 10 (s) |
Misura 10 (Hz) |
|
14.16 |
-0.071 |
|
8.54 |
-0.117 |
|
4.25 |
-0.235 |
|
3.98 |
-0.251 |
|
6.06 |
-0.165 |
|
9.66 |
-0.104 |
|
20.78 |
-0.048 |
|
25.63 |
-0.039 |
|
84.70 |
+0.012 |
|
Media frequenza di rotazione (Hz) |
Deviazione standard ( Hz ) |
|
-0.068 |
0.008 |
|
-0.121 |
0.010 |
|
-0.230 |
0.009 |
|
-0.238 |
0.011 |
|
-0.166 |
0.010 |
|
-0.102 |
0.009 |
|
-0.047 |
0.010 |
|
-0.036 |
0.012 |
|
+0.001 |
0.013 |
Osservare che nelle tabelle relative alla prova 2 per la frequenza 500000 Hz si è ottenuta talvolta una rotazione delle cellule in verso orario cioè conforme al campo e la si è indicata con un segno positivo.
Gli errori sulle misure
Mediante propagazione degli errori si ha (viste le (8) , (9), (10) e il valore del raggio r stimato (vedi prossimo paragrafo):
0000000(11)
0000000(12)
0000000(13)
0000000(14)
Nel caso della prova1 si ha:
(11)=0.4 mF/m2
(12)=0.4% + 2.3% + 2.4% = 5.1%
(13)=0.2
(14)=5.1%
Nel caso della prova 2 si ha:
(11)=0.5 mF/m2
(12)=0.4% + 2.3% + 4.5% = 7.2%
(13)=0.3
(14)=7.2%
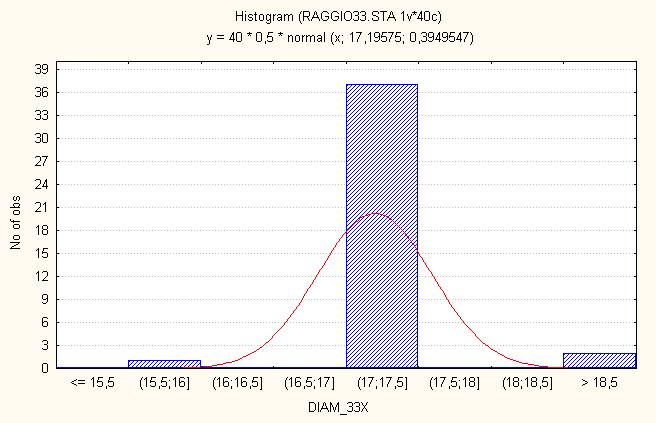
La stima del raggio
Mediante microscopio ottico (obiettivo 33X) si è stimato il diametro delle cellule 3T6. Si è usata come misura di taratura l’area di una "finestra" di un reticolo di diffrazione: Sapendo che tale area misurava 1/400 mm2 e che il lato di una tale "finestra" era formato da 35 divisioni si ricava l’unità di misura delle divisioni che per il nostro obiettivo risultava essere di 1.43
. Dai seguenti dati si ricavava una stima del diametro e quindi del raggio con un errore relativo molto contenuto.
|
MISURA (casi) |
DIAMETRO ( |
|
1 |
17.16 |
|
2 |
17.16 |
|
3 |
17.16 |
|
4 |
17.16 |
|
5 |
17.16 |
|
6 |
17.16 |
|
7 |
17.16 |
|
8 |
17.16 |
|
9 |
18.59 |
|
10 |
17.16 |
|
11 |
17.16 |
|
12 |
17.16 |
|
13 |
17.16 |
|
14 |
17.16 |
|
15 |
17.16 |
|
16 |
17.16 |
|
17 |
17.16 |
|
18 |
17.16 |
|
19 |
17.16 |
|
20 |
17.16 |
|
21 |
17.16 |
|
22 |
17.16 |
|
23 |
18.59 |
|
24 |
17.16 |
|
25 |
17.16 |
|
26 |
17.16 |
|
27 |
17.16 |
|
28 |
15.73 |
|
29 |
17.16 |
|
30 |
17.16 |
|
31 |
17.16 |
|
32 |
17.16 |
|
33 |
18.59 |
|
34 |
17.16 |
|
35 |
17.16 |
|
36 |
17.16 |
|
37 |
17.16 |
|
38 |
17.16 |
|
39 |
17.16 |
|
40 |
17.16 |
|
MEDIA diametro |
17.20 |
|
DEV.ST. diametro |
0.40 |
|
RAGGIO |
8..60 |
|
DEV. ST. |
0.20 |