| There are probably less than 700 gorillas of the subspecies G. g. beringei in existence. As conservation plans depend heavily on obtaining as much knowledge as possible about the endangered species of interest, here I have taken a look at the works of some leading researchers in gorilla conservation and presented the research as a comparison between the two known remaining popultions of mountain gorillas left. |
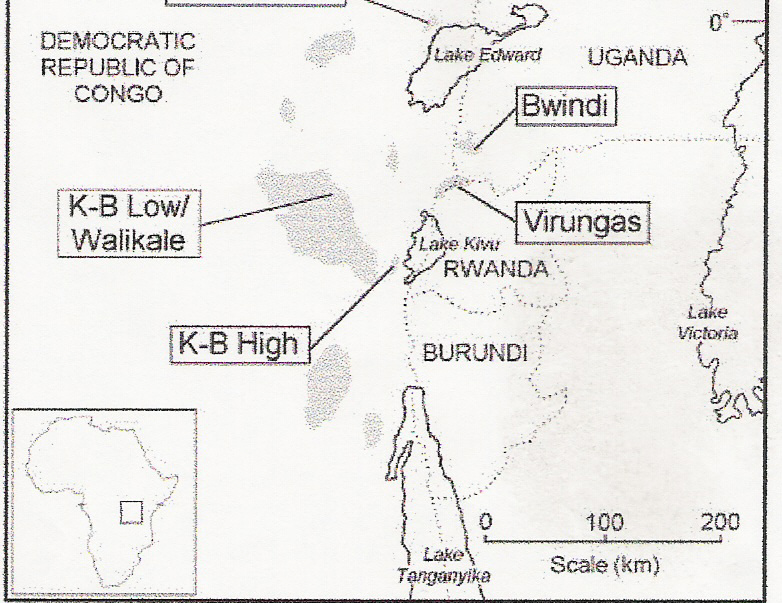
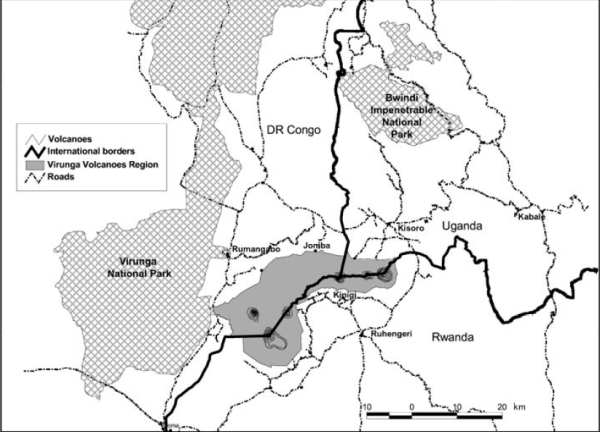
Largest of all extant primates, the gorilla (Gorilla gorilla) can be found living in the tropical forests of Africa. This species, one of the great apes, can be divided into three subspecies across two areas of Africa. The western lowland subspecies, G. g. gorilla, are distributed in western equatorial Africa in the lowlands around the Cameroon coast (Csomos 2001). About a thousand miles to the east (Jensen-Seaman & Kidd 2001) in central Africa, we find the eastern gorillas, which are divided into the subspecies of eastern lowland gorillas, G. g. graueri, and the mountain gorillas, G. g. beringei. The mountain gorillas are the least numerous of all three subspecies; the subspecies inhabits two sites with approximately 300 individual gorillas in each site (Jensen-Seaman & Kidd 2001). The respective sites for G. g. beringei include the Bwindi Impenetrable National Park in Uganda and the Virunga volcanoes region separating Uganda from Zaire and Rwanda (Lindsley & Sorin 2001). The two sites have markedly different climates that we can expect to have an impact on the mountain gorillas in each area. By comparing the two sites where populations of G. g. beringei reside, we can expect significant morphological, behavioral, and genetic differences between the two groups of gorillas. Before looking at the specific morphological and behavioral differences between mountain gorillas, it would be beneficial to first consider the habitats in which these groups reside. The Virunga region of Africa is much higher in elevation than the Bwindi forests (Sarmiento et. al. 1996). As a result of this higher elevation, the climate of Virunga volcaones is colder than the climate at Bwindi. The vegetation is effected in the expected ways, but it is of particular importance in our comparison to note that the cooler Virunga region promotes the growth of large areas of bamboo as well as several types of herbaceous foods (Sarmiento et. al. 1996). Trees are not as common in the Virunga region, but fruiting trees are a significant part of the ecosystem in the Bwindi forest. Figure 1 (Jensen-Seaman & Kidd 2001) provides us with a look at where the Bwindi and Virunga regions are located in Africa; the shaded regions indicate the range of gorillas. Ranges to the west of the Bwindi and Virunga regions are areas occupied by the eastern lowland gorilla, G. g. graueri. A closer, and more detailed, image of the Bwindi and Virunga regions can be seen in Figure 2 (Kalpers et. al. 2003).
The Bwindi gorilla census of 1997 can offer us more specific information on the Bwinidi region. The Bwindi Impenetrable National Park occupies an area of southwestern Uganda (McNeilage et. al. 1998). The region stands between 1160 and 2607 meters in altitude (McNewilage et. al. 1998); the forested areas of Bwindi was originally set aside as a reserve in 1932, and the region became a national park in 1991 in effort to protect the large amount of biodiversity that exists in the area (McNewilage et. al. 1998). The area of the park includes 331 square kilometers of a very rugged terrain surrounded by a large and growing human population (McNewilage et. al. 1998). Although the destruction of the forest has been banned, several areas in Bwindi are used for the purposes of growing medicinal plants and bee-keeping (McNewilage); tourists are also allowed to visit the area to observe the gorillas. All of these practices may impact the forest area and the gorilla population at Bwindi.
For a more detailed look at the Virunga volcanoes region, we can look online to the Biodiversity Support Program. The stretch of Virunga volcanoes is located mainly in the former Zaire in what is now the Democratic Republic of Congo (Kalpers 2001). The Virunga area consists of one uninterrupted region of forest that covers about 450 square kilometers. Its climate is both cool and humid, and the region is elevated from 2,300 to 4,511 meters high (Kalpers 2001), and holds an extremely high degree of biodiversity. Unfortunately, the area of the Virungas has a history of intense human conflict, which has done damage to the ecosystem as a whole. The region was set aside as park land to because of the mountain gorillas, but, due to the of the amount of biodiversity in the area, several species are being protected. After looking at the two sites that mountain gorillas inhabit, we can examine the many morphological differences between the two species. Being in a cooler climate with more herbaceous vegetation, we can hypothesize that gorillas at the Virunga site will have morphological adaptions for greater insulation and the processing of more leafy foods. In contrast, the Bwindi gorillas will likely have characteristics that are beneficial in warmer, more arboreal climates, which contain fruit producing trees. These hypotheses can be examined in great detail thanks to the morphological research conducted by Esteban E. Sarmiento, Thomas M. Butynski, and Jan Kalina, who compared physical characteristics in gorillas from both the Virunga and Bwindi sites. The intent of these researchers was to “quantify the skeletal, dental, and cranial features distinguishing Bwindi from Virunga gorillas, and explore the association of these features with their observed differences in habitat and behavior” (Sarmiento et. al. 1996). The skeletons used for the comparative anatomy came from a variety of institutes; specimens from gorillas at both sites and of both sexes were present, and there were two juvenile skeletons used as well (Sarmiento et. al. 1996). Several bones from the right side of the gorilla's body were measured for length with an osteometric board; these bones included the clavicle, os coxa, humerus, radius, femur, and tibia (Sarmiento et. al. 1996). Unfortunately, the number of specimens from the Bwindi site was very low, which made accurate estimates difficult. As is shown in Table 1, however, the small number of specimens available seem to show little difference in size between males at both sites. Sarmiento et. al. suggests that there may not be as great of a size difference for males between the populations as there is for females (1996). The skeletal length of the females show a significant difference in length. One interpretation of these measurements might be that large size in males is due to intrasexual selection; females, in contrast, may exhibit a size that reflects the result of more ecological forces. Further comparison of skeletal features suggest a considerable difference between female size in both sites. The lumbar vertebra volume in particular is noted by Sarmiento et. al. as an indicator of this difference (1996), and these measurements tend to be supported by visual observations in the field.
In addition to skeletal features, external features were also compared by Esteban Sarmiento and his colleagues. The features these researchers found to be different in the two populations include the body and facial hair, the adult male saddle, the facial skin, the nose, and the sole of the foot (Sarmiento 1996). The body hair, to start with, was found to be shorter in gorillas of the Bwindi region with a brown tint to the color; this is in contrast to the long and shaggy black hair of the Virunga gorillas. Likewise in the Bwindi gorillas, the facial hair observed tended to be shorter and did not hide the ears of the gorillas; some adult males were also found with white hair on the face. In the Virunga gorillas, the facial hair, like the bodily hair, was longer and managed to hide the ears; hair on the face of the Virunga gorillas also formed a whiskers or a beard, which was not present in gorillas of the Bwindi forests. The skin of the face of Bwindi gorillas was not wrinkled as heavily as the skin of Virunga gorillas. The structure of the nose between gorillas at the two sites differs greatly; Bwindi gorillas completely lack a dorsonasal cleft or overhanging lip above the septum, but both of these structures are present in gorillas of the Virunga population. Also, Virunga gorillas have larger nostrils compared to individuals of the Bwindi forest. The saddle of male silverbacks in the Bwindi region included white hairs from the rump to the front of the thighs, whereas the white hair color of silverbacks in the Virunga region takes on a more saddle-like appearance with white hairs occurring between the shoulders and the rump area. Finally, the soles of the foot in both sites of gorillas extend “up to the midshaft of the middle phalanges between toes II-V” (Sarmiento et. al. 1996). The sole of gorillas in the Bwindi forest differ in that the big toe cleft goes as far as the first metatarsal head; in the Virunga gorillas, the cleft is distal to the base of the first proximal phalanx. Several of these characteristics, including the long hair and large nasal structures, can be predicted by considering the cooler climate of the Virunga volcanoes. Sarmiento and his colleagues also examed how these morphological differences may be related to the gorilla's environment, and the researchers took an especially close look at the dietary habits of both groups of gorillas. They note that the size differences between gorillas in the Bwindi and the Virunga sites reflect what we might expect from the differences that are observed in the gorilla's diets. The diet of Virunga gorillas is composed of almost 55% bamboo (Sarmiento et. al. 1996), which is strikingly different from the diet of gorillas in the Bwindi because gorillas of the Bwindi rarely encounter areas that have bamboo growing. Instead, the Bwindi gorillas focus much more on consuming fruit for their dietary needs (Sarmiento et. al. 1996). It should be noted that this relationship of size and diet is exactly what we expect in species of mammals where a larger body is needed to process the less nutritious cellulose found in herbaceous foods. Personal observations have also been made by Sarmiento that larger males within each site consume “more roughage and less fruits than the smaller females or juveniles” (Sarmiento et. al. 1996). To what extent, if at all, large body size between the sites is driven evolutionarily by the availability of these specific foods is, of course, unknown. At this point we should also consider the work of Amy L. Vedder, who studied the movement patterns of mountain gorillas in relation to food availability. By studying the work of Vedder, we can get a better understanding of the dietary habits of Virunga gorillas, as this was the site that Vedder focused on. Vedder expected that, as a large herbivore, the gorillas at Virunga would require little dietary expansion and a lot of time traveling in search of high-quality foods, with the gorillas spending an especially extended period of time in the areas where these foods are located (Vedder 1984). Vedder divided the Virunga region, and examined food distribution throughout the area of study. The frequency, quality, and temporal availability of foods were considered, and a group of gorillas was followed for 311 days to record the group's ranging patterns (Vedder 1984). Zones from the Virunga region were separated into seven categories. These regions included the Bamboo region, which was most prominent in the lowest zone of the study area. The largest vegetation zone was the Hagenia-Hypericum woodland area between two mountains where a herbaceous undergrowth provides food for gorilla populations (Vedder 1984). A bit higher up, the Herbaceous slope exists with similar undergrowth as the Hagenia-Hypericum area, and patches of shrubs with various densities make up the Vernonia Zone. The Hypericum woodland occupies a higher altitude that is covered with woodland and blackberries; the Giant senecio is the second highest region in altitude; this area holds trees approximately three meters tall, and there is little undergrowth (Vedder 1984). Short grasses cover the Alpine region, which is the highest region Vedder describes in the study; the region reaches an elevation of 3,710 meters (1984). Vedder noted all of the flora that gorillas would use as food then calculated the biomass of food in plots within the regions. Vedder also looked at the temporal availability of the flora and found that bamboo was actively preferred when it was available in from October to December (Vedder 1984). Availability of the bamboo was taken each month be recording individual shoots in different size classes. In the months when bamboo was not available to the gorillas, the primary food consumed came from herbaceous, with a small amount of the diet coming from woody growth of immature plants (Vedder 1984). As was predicted, gorillas of the Virunga do spend more time in areas of richer food, and the hypothesis that ranging patterns of gorillas at the Virunga site are determined by food availability appears well supported. Furthermore, Vedder found that during times of less rainfall when food was more scarce, gorillas increased their home range size, but whether or not the diet was expanded as well was not analyzed (1984). The folivorous diet of gorillas in the Virunga regions differs from the diet of those in the Bwindi forests in many ways. For an analysis of home range and feeding patterns of Bwindi gorillas, we can turn to a study done by Martha M. Robbins and Alastair McNeilage. Robbins and McNeilage noted earlier studies that observed larger home ranges and a more frugivorous diet in the Bwindi gorillas (2003). The central goals behind their study included looking at the occurrence of fruit eating in gorillas of the Bwindi, measuring home range sizes, looking for correlations between fruit eating and home range sizes as well as social factors and their influence on home range sizes. Regions were not broken down in the same way as they were in Vedder's study, but Robbins and McNeilage made note that the altitude range of Bwindi is equal to the Virunga site in about 33% of the Virunga area (Robbins & McNeilage 2003). Home range sizes were determined with the help of GPS readings, and, almost daily, field assistants tracked the gorillas of interest to observe their feeding habits (Robbins & McNeilage 2003). The diet of the Bwindi gorillas included 16 species of fruit; several of these species were also included in the diet of Grauer's gorilla, G. g. graueri (Robbins & McNeilage 2003). Some of these fruits, as Robbins and McNeilage pointed out, were also species that were observed to be eaten by Virunga gorillas in the lower altitudes of the Virunga site. Over three years of study, researchers were able to calculate fruit eating occuring on 27% of the days observed; this figure puts fruit consumption of Bwindi gorillas quite higher than consumption in gorillas of the Virunga site, yet still lower than observed in Grauer's gorillas (Robbins & McNeilage 2003). Fruit consumption also varied temporally in Bwindi gorillas; less fruit was consumed between the months of September and December, and their was an increase in frugivory between January and June (Robbins and McNeilage 2003). Interestingly enough, Robbins and McNeilage found no relationship between fruit eating and home range size (2003); this seems strange given Vedder's findings that show support for Virunga gorillas spending greater time in areas of high quality food (1984). Robbins and McNeilage suggest several reasons for their findings; they suggest that Bwindi gorillas may feed only opportunistically on fruit, thus frugivory would not effect range patterns (2003), but this seems inconsistent with their results, which clearly show Bwindi gorillas moving great distances in search of fruit trees. More complicated relationships may be suggested; gorillas may cross the same path twice and increase journey length without increasing home range, or the relationship between frugivory and home range size may be blurred by other factors such as a poorly defined fruiting season (Robbins & McNeilage 2003). Whatever the case, it was easy to conclude that the home range size of mountain gorillas at the Bwindi site was certainly larger than home ranges found in Virunga gorillas. Robbins and McNeilage suggest that this fact might have something to do with the high density of herbaceous foods that are found in the diet of Virunga gorillas (2003). The authors go on to discuss several important conservation suggestions, which are beyond the scope of this paper. In addition to looking at the morphological differences and the respective feeding habits and range patterns between mountain gorillas at the Virunga and Bwindi sites, interesting comparisons can be made on the genetic level between these two different groups. Studies in population genetics relating to mountain gorillas can be obtained from a small handful of sources. Several genetics studies in these populations focus heavily on mitochondrial DNA variability of populations. M. I. Jensen-Seaman and K. K. Kidd conducted a field study in 2001 by looking at the DNA displacement loop (D-loop); the researchers were looking at the “patterns of D-loop DNA sequence diversity” over eastern gorillas in general. Part of the reason the researchers chose the “D-loop” is because the site is very rapidly evolving and polymorphic; it should also be noted that, because this site is in the mitochondrial genome, no recombination takes place here. Researchers took hair samples from nests of gorillas and zoos with gorillas of known origin (Jensen-Seaman & Kidd 2001). They found that 27.5% of the genetic variation that lies within mountain gorillas as a whole is partitioned among populations; the remainder of the variation is found within the populations (Jensen-Seaman & Kidd 2001). In other words, there is considerably more genetic variation inside individual populations of gorillas than there is from one population to another. Jensen-Seaman and Kidd also note that gorillas at the Bwindi and Virunga sites share a very close genetic relationship, which is closer than expected given the morphological differences between the gorillas at the two sites (2001). This suggests that gorillas at these sites were recently in reproductive contact, and that there may have been a rapid morphological evolution (2001); this rapid divergence may have arisen in part from the differences in habitat discussed earlier. Jensen-Seaman and Kidd were unable to find an accurate time since gorillas at these sites diverged from each other. Still, genetic data can provide us with several interesting comparisons between these two groups of gorillas. Bringing all of these studies together, we can get a good picture of the similarities and differences between gorillas at the Bwindi and Virunga sites. The individual sites themselves show a considerable difference in terrain. The Virunga volcanoes region occupies a higher elevation than the Bwindi Impenetrable National Park area. The colder climates of the Virunga site are complemented by gorillas with longer hair and larger female body size than those of the Bwindi forest. Although the longer hair in Virunga gorillas may be an adaption to the colder climate, we should be hesitant to say the same for the larger body size of the females. If an increase in body size were such an adaption, we might also expect a reduction in limb size to reduce heat loss; by looking at Table 1, we can see that this does not appear to be the case. The body size of female mountain gorillas, however, might be influenced by diet in some way. The vegetation of the Virunga site includes different types of flora, such as bamboo and several herbaceous plants that gorillas of the Virunga feed on. In contrast, the Bwindi region has more fruiting trees, and fruit comprises a significantly higher proportion of the diet for gorillas in the area. There are still interesting comparisons left to be made between gorillas at these sites. More detailed genetic studies might help us understand the evolutionary relationship between these groups; such studies might give us an indication of when these groups diverged from each other and whether or not gene flow still occurs between gorillas at the two different sites. Also, while several behavioral studies have been done on gorillas of the Virunga volcanoes, research in this area on gorillas of the Bwindi forest is sparse in comparison. As there are less than a thousand individuals of G. g. beringei remaining in the wild, such studies could go a long way in conservation efforts to preserve the important genetic and cultural diversity that exists in this subspecies of gorilla. Works Cited
Csomos, R. 2001. "Gorilla gorilla" (On-line), Animal Diversity Web. Accessed November 27, 2005 at <http://animaldiversity.ummz.umich.edu/site/accounts/information/Gorilla_gorilla.html>. Jensen-Seaman, M. I. Evolutionary Genetics of Gorillas. Diss. Yale, 2000. Ann Arbor: Proquest, 2005. Jensen-Seaman, M. I. & Kidd, K. K. 2001. Mitochondrial DNA variation and biogeography of eastern gorillas. Molecular Ecology, 10, 2241-2247. Kalpers, J. 2001. Volcanoes under siege: impact of a decade of armed conflict in the virungas. Biodiversity Support Program. 27 Nov. 2005 <http://www.worldwildlife.org/bsp/publications/africa/144/titlepage.htm>. Kalpers, J., Williamson, E. A., Robbins, M. M., McNeilage, A., Nzamurambaho A., Lola, N., Mugiri, G. 2003. Gorillas in crossfire: population dynamics of the Virunga mountain gorillas over the past three decades. Oryx, 3, 326-337. Lindsley, T. and A. Sorin. 2001. Gorilla gorilla beringei (On-line), Animal Diversity Web. Accessed November 27, 2005 at <http://animaldiversity.ummz.umich.edu/site/accounts/information/Gorilla_gorilla_beringei.html>. McNeilage, A., Plumptre, A. J., Brock-Doyle, A., Vedder, A. 1998. Bwindi Impenetrable National Park, Uganda: gorilla census 1997. Oryx, 35, 39-47. Robbins M. M., McNeilage A. 2003. Home range and frugivory patterns of mountain gorillas in Bwindi Inpenetrable National Park, Uganda. International Journal of Primatology, 24, 467-491. Sarmiento E.E., Butysnksi TM, Kalina J. 1996. Gorillas of Bwindi-impenetrable forest and the Virunga volcanoes: taxonomic implications of morphological and ecological differences. American Journal of Primatology, 40, 1-21. Vedder, A. 1984. Movement patterns of a group of free-ranging mountain gorillas (Gorilla gorilla beringei) and their relation to food availability. American Journal of Primatology, 7, 73-88 |