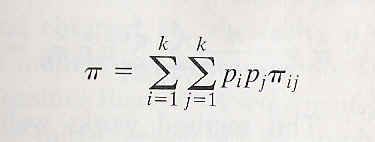|
|
|
Introduction
The importance of population genetics in species conservation is becoming increasingly apparent. Population genetics is defined as an area of study that “considers the factors that determine the evolution of a population, such as natural selection, genetic drift, mutation, recombination, and gene flow” (Halliburton 2004). Although some may view the field as an abstract subject that has little relevance to the real world, population genetics provides us with techniques for assessing relevant information such as genetic diversity and population fitness (Halliburton 2004). More specifically, if we are to ignore the field of population genetics, we may find ourselves adopting recovery strategies that are inadequate for species conservation (Frankham 2005). For this reason, a basic understanding of population genetics should be reviewed for those who are involved in conservation management. The techniques discussed will aid in the conservation biology of the eastern gorillas (Gorilla gorilla graueri and G. g. beringei). Relevant topics of population genetics will include protocols for the analysis of genetic diversity in and among populations, as well as the effect of factors such as genetic drift and migration in gorilla populations of the eastern lowlands. Overall, the incorporation of such issues found in population genetics will give us a better idea of how to make a more effective management program for the species. A Review of Population Genetics Before reviewing the literature that is relevant to gorilla conservation, it is necessary to discuss some of the techniques used to evaluate conservations strategies. To ultimately assess population genetics data, we must first have a way of obtaining DNA samples from the populations of interest. In the case of gorilla studies, it was hair samples that were collected from vacated nests (Jensen-Seaman & Kidd 2001; Saltonstall et. al. 1998). A series of protocols was used to produce a template of DNA, and then a polymerase chain reaction was performed to amplify the DNA. A polymerase chain reaction, or PCR, is a fast and inexpensive way to replicate a nucleotide sequence in large amounts from a DNA sample that has the particular sequence (Alberts et. al. 2004). After running a PCR to amplify the DNA, the DNA may be sequenced; in other words, this technique allows us to view the genetic code of whatever region of DNA we are interested in. Once this code is obtained, we can calculate genetic diversity in and among populations. Genetic diversity can be quantified in several ways, which includes the measurement of the amount of heterozygosity in a population and the amount of polymorphic loci in a population. A particularly strong measurement of genetic diversity that will be used to assess the gorilla populations of interest is the value of nucleotide diversity. Nucleotide diversity is the average pairwise difference between any two randomly selected copies of nucleotides (Duvernell 29 Aug. 2005). In other words, nucleotide diversity measures the probability that any two randomly selected nucleotides are different. Nucleotide diversity is most often symbolized by pi (Halliburton 2004). We can calculate pi with the following formula:
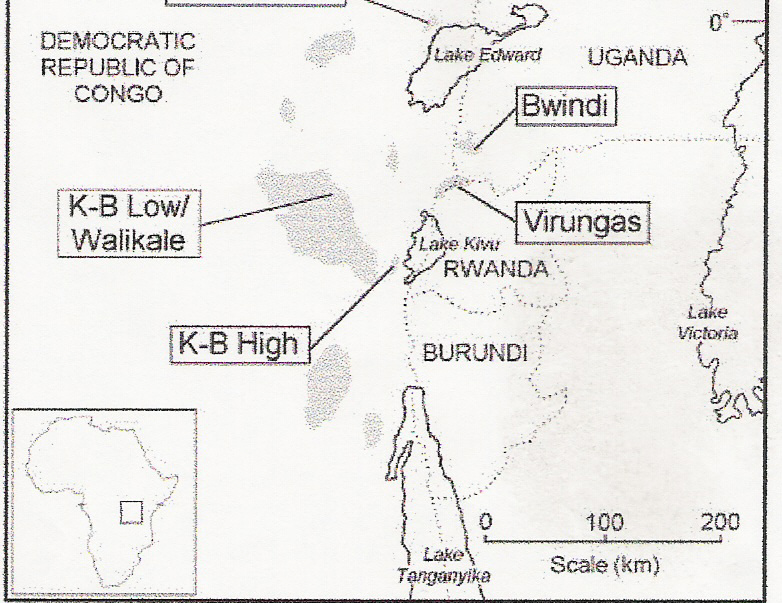
In this formula, piij is the proportion of nucleotide differences between the ith and jth haplotypes where k is equal to the number of different haplotypes that exist in the population; the symbols pi and pj are the frequencies of the two haplotypes of interest (Halliburton 2004). The n/n-1 is used to correct a sampling bias. The reader may note that the strength of quantifying genetic diversity in this technique comes from the technique's measurement of each nucleotide's locus in relation to every other locus in the sample; generally this will give us a better estimate of genetic diversity than the proportion of heterozygotes or the proportion of polymorphic nucleotide sites. Two additional topics to touch on that are relevant in species conservation are the concepts of genetic drift and migration, which are actually observable as two counteracting forces. Genetic drift is defined as a “random fluctuation of allele frequencies due to random sampling of gametes and other chance events that occur in each generation” (Halliburton 2004). In the long run, genetic drift will lead to an overall loss of diversity as some alleles are lost from a population. Since genetic drift is more effective in eliminating genetic variation in small populations, endangered species run the risk of losing a great deal of genetic diversity; such populations often become inbred, which contributes to an overall reduction in fitness in the species. Migration, or gene flow, which is characterized by an event when individuals move from one area to another, sometimes into a new population, counteracts drift by introducing new alleles into a population and increasing genetic variation. In this way, migration is equivalent to mutation, which is the ultimate source of all genetic variation. A brief explanation should be given for the concept of inbreeding depression because inbreeding depression is of special concern in small population as seen eastern gorillas. Inbreeding depression is caused primarily by the pairing of recessive genes that are identical by decent; genes that are identical by decent are genes that share a common ancestry (Wallis 1994). The pairing of recessive alleles that are deleterious results in a reduction of mean fitness in a population (Duvernell 31 Oct. 2005). Mitochondrial DNA Studies of Eastern Gorillas of Interest for Conservation Armed with a knowledge of how to assess genetic variation in populations, we can apply our understanding of such techniques to look at several studies of gorilla populations, and how genetic data can be used to evaluate different conservation strategies. Eastern gorillas are found in equatorial Africa, and are separated from their western counterparts by approximately 1,000 km (Jensen-Seaman & Kidd 2001). The two extant subspecies of eastern gorillas include the eastern lowland gorilla, G. g. graueri, and the mountain gorilla, G. g. beringei. G. g. graueri numbers 8,150 individuals in size, compared to the size of the two populations of G. g. beringei, which contain about 300 individuals in each population (Jensen-Seaman & Kidd 2001; Saltonstall et. al. 1998). The small number of eastern gorillas left in existence and the continued destruction of the tropical forests, habitat loss, and poaching are all valid reasons for the World Conservation Union to declare eastern gorillas as endangered (Yu 2004). The especially small amount of mountain gorillas left in the areas around the Virunga Volcanoes and the Bwindi-Impenetrable Forest makes the subspecies G. g. beringei extremely vulnerable to extinction (Sarmiento et. al. 1996). Although G. g. graueri is also endangered, the population has been doing fairly well and has even been suggested to be increasing in numbers (“Gorilla” 2005). Although efforts have had a positive impact on the survival of the species, a look from the field of population genetics will help us construct the best possible conservation plan for eastern gorillas. Studies in population genetics relating to eastern gorillas can be obtained from a handful of primary sources. Several studies in these populations focus heavily on mitochondrial DNA variability of populations. M. I. Jensen-Seaman and K. K. Kidd conducted a field study in 2001 by looking at the DNA displacement loop (D-loop); the researchers were looking at the patterns of D-loop DNA sequence diversity over eastern gorillas in general. The reason the researchers chose the “D-loop” is because the site is very rapidly evolving and polymorphic; the D-loop is found on the mitochondrial genome, where no recombination takes place. In the study, hair samples were collected from various locations and in both subspecies of eastern gorillas; fecal samples were also collected. A PCR was run to amplify the DNA, and the DNA was sequenced so that the genetic code could be observed. The results suggested clades of what we might expect from looking at the morphological differences between the beringei and graueri populations. There were not any haplotypes found in both graueri and beringei, and there were no haplotypes found that were seen previously in western lowland gorillas (Jensen-Seaman & Kidd 2001). In other words, graueri and beringei populations diverged at some earlier date; the authors put the separation somewhere around 400,000 years ago with an unknown, but probably large confidence interval (Jensen-Seaman & Kidd 2001). These two populations of eastern gorillas are more closely related to each other than western gorillas, however, with the nucleotide diversity values being an entire order of magnitude smaller than in western gorillas; the estimated nucleotide diversity for all eastern gorillas is 0.660 (Jensen-Seaman & Kidd 2001). Surprisingly, these results show a higher degree of genetic similarity than what might be selected by anatomical comparisons. This suggests that the two subspecies have been in recent reproductive contact (Jensen-Seaman & Kidd 2001). The researchers also found that, within graueri and beringei subspecies, all populations shared haplotypes with others; this indicates recent gene flow between different populations of both of the individual subspecies. Figure 1 below, used in Jensen-Seaman and Kidd's article, shows the location of these populations (Jensen-Seaman & Kidd 2001). Mountain populations of the subspecies beringei include the Bwindi and Virungas sites; graueri populations occupy the remaining sites shaded in the figure.
An earlier study also involving mitochondrial DNA can aid in our decisions for future plans involving the conservation of the eastern gorilla. In the study, K. Saltonstall, G. Amato, and J. Powell, analyzed gene flow between populations of the subspecies graueri was studied, as information such as this is necessary to establish conservation priorities for eastern gorillas (1998). As in the study of Jensen-Seaman and Kidd, hair samples were collected from populations of gorillas. Samples from G. g. graueri individuals were collected and compared within graueri populations and between other subspecies. PCR was run to amplify the DNA samples, which were sequenced to quantify the information that was obtained. The mitochondrial gene that was used in this study was coded for cytochrome oxidase subunit II (COII) and a 277 base pair hypervariable portion of the mitochondrial DNA D-loop (Saltonstall et. al. 1998). A total of 39 gorillas were used for sequencing. In these 39 individuals, all of the sequences for the COII gene were identical (Saltonstall et. al. 1998). The COII gene sequence differed from the subspecies G. g. beringei at two positions; this supports the conclusions of Jensen-Seaman and Kidd that the two subspecies have been reproductively isolated (Saltonstall et. al. 1998). Due to DNA degradation, only 15 individuals were used for examining D-loop sequences; six individual haplotypes were found differing by 0.4 to 1.8% (Saltonstall et. al. 1998). Haplotypes from previous studies by K. J. Garner and O. A. Ryder were also used for comparison. Conclusions Regarding Conservation The previously described studies, including the methods of population genetics, are important in planning conservation strategies for eastern gorillas. The studies have shown that despite a large amount of morphological difference between the subspecies G. g. graueri and G. g. beringei, the two groups are closely linked genetically. This indicates that the two subspecies may be in reproductive contact, but it is unlikely that the levels of gene flow are significant (Saltonstall et. al. 1998). Unfortunately, individual populations are often separated by land inhabited by human populations. Isolated populations that occur in eastern gorillas may suffer problems associated with inbreeding depression, which, as noted earlier, is a concern with small isolated populations. To avoid the union of alleles that are identical by descent and thus the negative effect on fitness caused by inbreeding depression, eastern gorillas should be allowed as much freedom as possible to migrate between populations. Such a conservation plan would include not only protecting the forest areas within populations, but also forested land between different populations of eastern gorillas. As discussed, there is clearly a significant amount of genetic diversity between populations of eastern gorillas; isolating this diversity could be detrimental to both subspecies of gorilla. K. Saltonstall, G. Amato, and J. Powell call for similar measures, particularly between two graueri populations east of lake Kivu that appear to be diverging due to an evolutionarily significant level of gene flow (1998). The authors believe that every effort should be made to restore and maintain the corridor that connects the two sections of the park; this would allow gorillas to continue traveling between the two habitats (Saltonstall et. al. 1998). Measures such as these applied to other populations of eastern gorillas would increase or at least maintain genetic diversity in subspecies of gorilla by allowing gene flow to occur between the populations. This would be beneficial in reducing the negative effects of inbreeding depression, and would ultimately result in maintaining stable population sizes of eastern gorillas. Works Cited
Alberts, Bruce, Dennis Bray, Karen Hopkin, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walter. Essential Cell Biology. 2nd ed. New York: Garland Science, 2004. 347-351. Duvernell, David D. Lecture. Southern Illinois University Edwardsville, Illinois. 29 Aug. 2005. Duvernell, David D. Lecture. Southern Illinois University Edwardsville, Illinois. 31 Oct. 2005. Frankham, Richard. "Genetics and extinction." Biological Conservation 126 (2005): 131-140. "Gorilla Group May Be Growing." Wildlife Conservation 2005: 12. Halliburton, Richard. Introduction to Population Genetics. Upper Saddle River, NJ: Pearson Education Inc., 2004. Jensen-Seaman, M I., and K K. Kidd. "Mitochondrial DNA Variation and Biogeography of Eastern Gorillas." Molecular Ecology 10 (2001): 2241-2247. Sarmiento, Esteban E., Thomas M. Butynski, and Jan Kalina. "Gorillas of Bwindi-Impenetrable Forest and the Virunga Volcanoes: Taxonomic Implications of Morphological and Ecological Differences." American Journal of Primatology 40 (1996): 1-21. Saltonstall, K, G Amato, and J Powell. "Mitochondrial DNA Variability in Grauer's Gorillas of Kahuzi-Biega National Park." Journal of Heredity 89 (1998): 129-135. Wallis, Graham P. "Population and Conservation Genetics in New Zealand: A Hierchical Synthesis and Recommendations for the 1990s." Journal of the Royal Society of New Zealand 24 (1994): 143-160. 6 Nov. 2005 <http://www.rsnz.org/publish/jrsnz/1994/9.pdf>. Yu, Ning, Michael I. Jensen-Seaman, Leona Chemnick, Oliver Ryder, and Wen-Hsiung Li. "Nucleotide Diversity in Gorillas." Genetics 166 (2004): 1375-1383. |