![]()
Of all the pathogens that plague life on this planet, viruses are among the most puzzling. We can understand cellular forms attacking other cellular forms, because they are kin to us. That is, a lion making its kill and tearing off chunks of raw flesh from its still-living victim, is not all that different from us masticating a steak that only yesterday was a harmless cow placidly munching its cud, among its dewy-eyed companions. But there seems to be something ALIEN about viruses (and a couple of other nasty "things" out there that should scare you) because they seem so different from us and their form of disease production so loathsome. However, if you take the broader view that you've been exposed to in this educational pilgrimage you will understand that viruses have the same goal in life of every other living "creature" on this planet.
Viruses are basically a way a form of genetic information insures its continued survival. This is another UNIVERSAL PRINCIPLE OF ALL LIFE, but it is difficult concept to "peddle" and to accept. I can't speak for you, but I feel I'm more than a bloody bag for hauling around DNA for a few years. For a discussion of this perspective I recommend an article "God's Utility Function" in the Nov. 1995 Sci. Am., pg. 80 by R. Dawkins. Many of you will be offended or made uncomfortable by the thesis of this article, but it will give you something to think about and it clearly has relevance to our discussion on the life and times of these small but very repellent creatures. If you can view the following description of viral life style as having the GOAL OF SURVIVAL as its top priority, understanding will inevitably follow. For a database of all the known viruses visit here and view some IMAGES to get a feel of viruses (HIV).
![]()
As the work of Koch, Pasteur and others became KNOWN in the late 1800s, the GERM THEORY OF DISEASE was accepted. With the exception of inherited and environmentally produced diseases, most diseases were recognized as being caused by infectious agents. Microbiologists were discovering new ETIOLOGICAL AGENTS (agents that cause disease) of disease almost daily in those early times. Bacterial, protozoan and fungal pathogens seemed to be everywhere and every new disease that turned up offered the chance for fame if one discovered a new pathogen (plus a chance to get it named after you---history book fame).
|
|
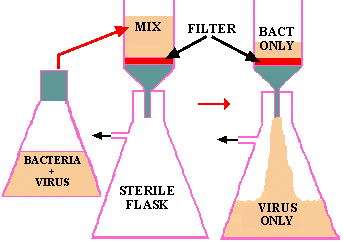
In 1884 C. Chamberland, in Pasteur's lab, discovered that if you passed a liquid containing bacteria through an unglazed PORCELAIN tube, the bacteria were COMPLETELY RETAINED and the solution that passed through (the FILTRATE) was sterile. The advantages of this tool were immediately apparent, for with it one could sterilize solutions containing heat-sensitive components by filtration through sterile porcelain tubes into sterile containers. By carefully controlling the components of the porcelain tubes you could CONTROL THE PORE SIZE and selectively remove larger organisms while letting smaller ones pass through.
This type of filtration immediately became one means of testing the Germ Theory, since if you passed an infected sample through a filter that would hold back all microbes, the filtrate should not induce the disease in a new host if a microbe was responsible. You could then begin to devise ways of growing the suspected pathogen. However, in 1892 D. IWANOWSKI applied this test to a filtrate of plants suffering from TOBACCO MOSAIC DISEASE with shocking results; the filtrate was FULLY CAPABLE of producing the ORIGINAL DISEASE in new hosts. When repeated filtration's produced the same results and nothing could be seen in the filtrates using the most powerful microscopes, nor could anything be cultivated from the filtrates, Iwanowski and associates concluded that they had discovered a new pathogenic life-form which they called by the unimaginative, but functional, name of "FILTERABLE VIRUS". We now know that viruses range in size from 20 nm (10-9 meters) to 250 nm. We also know that some of the smallest bacteria such as the chlamydia and mycoplasma are almost as small as the largest viruses and that they too can pass through filters that retard 99% of the other bacteria.
By the early 1900 diseases like foot-mouth-disease in cattle,
some cancers (in animals) and yellow fever in humans had been demonstrated to be
caused by filterable viruses. The scientific community knew that it had a new
group of dangerous pathogens to contend with. The term "VIRUSES"
became permanently associated with this life form. You have previously seen that
 bacterial
viruses (view the "Single Burst Graph Animation") or bacteriophage
(phage) were discovered in 1915 & 1917. Viruses, however were not
"seen" until the electron microscope was developed in the late 1930s.
This site contains electron micrographs of many bacteriophage; view the
bacterial
viruses (view the "Single Burst Graph Animation") or bacteriophage
(phage) were discovered in 1915 & 1917. Viruses, however were not
"seen" until the electron microscope was developed in the late 1930s.
This site contains electron micrographs of many bacteriophage; view the ![]() P2,
lambda and P4 phage.
P2,
lambda and P4 phage.
|
|
We now know that viruses exist that attack perhaps every form of cellular life on this planet. I haven't seen references to thermophilic phage, but I would be surprised if they didn't exist. We are discovering new viruses all the time and most virologists feel we have only scratched the surface of viral variety. For example, when sea water is concentrated and examined under the electron microscope it teems with VIRUS-LIKE PARTICLES and we have no idea what they are or where they come from or what their host's are?
The nature of viruses became even more confusing when it was observed in 1935 that they could be CRYSTALLIZED like inorganic salts (table salt) and protein molecules. This observation started a spirited, but rather barren, argument as to whether viruses are really "alive" or a "form of life". People have argued that viruses are like salt crystals that grow and reproduce (sort of). In my view this discussion is a waste of time by people who need to "get-a-life". Viruses clearly REPLICATE their genetic material, which like that of all other life forms, is composed of nucleic acid polymers. Viruses have one major characteristic in common: they are OBLIGATE INTRACELLULAR PARASITES. Viruses are UNABLE to grow and reproduce OUTSIDE OF A LIVING CELL. Therefore their survival is absolutely dependent upon the continued survival of their hosts. This poses an interesting dilemma for pathogens that often as not kill their hosts, wouldn't you say?
The intracellular nature of viruses presents a challenge for the investigator who must not only grow the virus but also be able to cultivate the virus' host cell. With plant and bacterial viruses it was possible to extract sufficient virus from an infected host to do analysis on it. These studies showed that viruses were mainly COMPOSED OF PROTEIN AND NUCLEIC ACID. With multicellular eukaryotic viruses the field of virus investigation could only move as rapidly as the advancements in eukaryotic TISSUE CULTURING. The first breakthrough in this problem came with the discovery in 1931 that the fertilized hen's eggs could serve as a "petri dish" for some viruses. This capacity led to the first use of artificially cultivated viruses for vaccine production. Even today many viruses are grown on eggs because they are relatively inexpensive and because the techniques are so well established.
![]()
Have you ever had the doctor ask "Are you allergic to eggs?" before receiving a viral immunization; that means that the virus were grown on eggs and that egg proteins (to which many people are allergic) are present in the vaccine.
![]()
With the discovery of the electron microscope it became possible to study the morphology of viruses. It was quickly realized that the size and shape of an individual virus is a constant and distinguishing characteristic. Therefore a viruses' size and shape is always part of its description. Viruses may consist of circles, ovals, long thick or thin rods, flexible or stiff rods and ones with distinctive heads and tail components. The smallest viruses are around 20 nm in diameter and the largest around 250 nm.
Some electron micrographs and computer renderings of viruses are seen at the following Internet addresses :
These were all functional as of 12/27/00:
Many good EM pictures of virus types
http://www.bocklabs.wisc.edu/virusviz.html Lots of very good virus pictures.
http://www.tulane.edu/~dmsander/garryfavweb.html; huge site for viruses with courses and virus pictures etc.
http://www-micro.msb.le.ac.uk/335/335Structure.html; Excellent discussion of virus structure.
http://www-micro.msb.le.ac.uk/109/structure.html; Many pictures, cartoons and good explanations.
![]()
Viruses are unique from all other life forms in that they can contain ONLY ONE FORM OF NUCLEIC ACID. Some viruses use RNA as their genetic material and other use DNA, but NEVER do they contain both. Further, this nucleic acid polymer may either exist as DOUBLE STRANDED (DS) DNA or RNA or as SINGLE STRANDED (SS) DNA or RNA. Each of these characteristics is a constant for a particular virus and is part of it description. The nucleic acid polymer may contain as few as 4 to 7 genes for very small viruses to 150 to 200 genes for very large viruses. In some viruses the nucleic acid exists in more that one molecule. Some viruses contain a few enzymes and some contain none, but no viruses contain the large numbers of enzymes found even in the smallest bacteria.
|
|
|
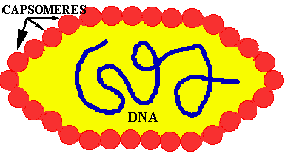
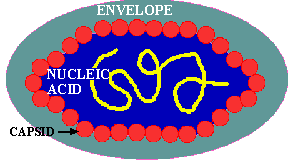
All viruses are covered with a PROTEIN COAT. This protein coat is mainly composed of a FEW TYPES of proteins of which there are many copies per virus; something like the individual threads in a shirt. These identical protein subunits are called CAPSOMERES and they are made so that they spontaneously come together (ASSEMBLE) in a PREDETERMINED way to produce the virus coat which is called the CAPSID.
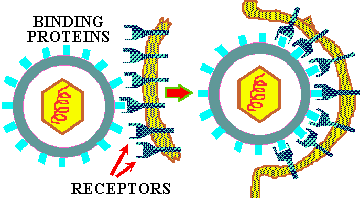
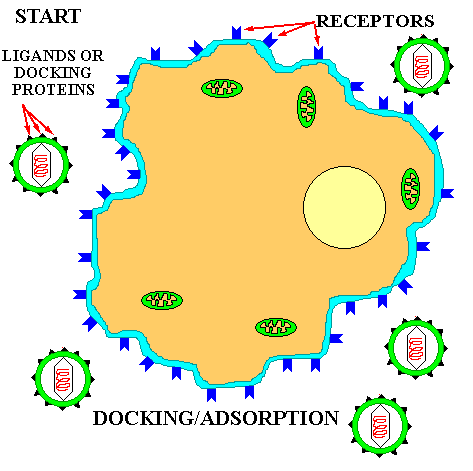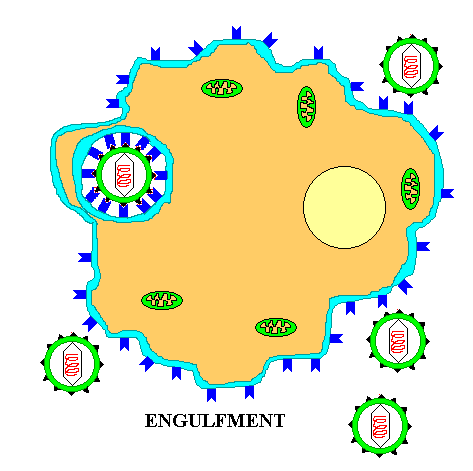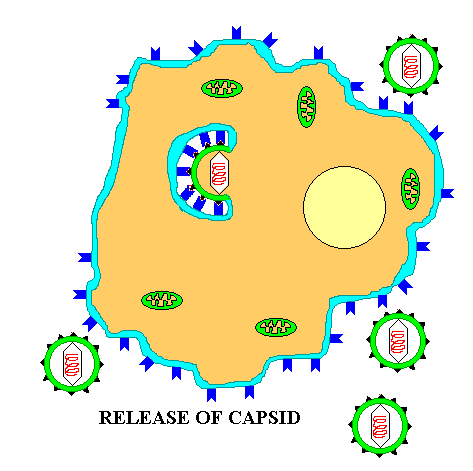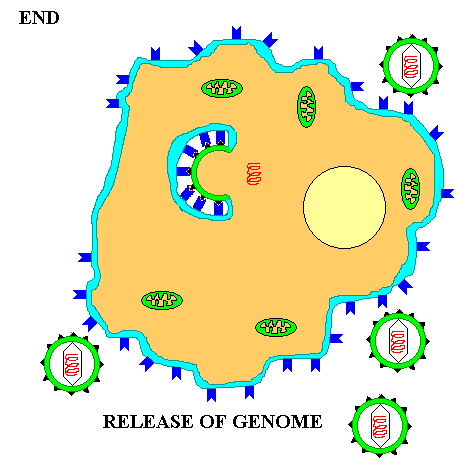
If a virus has ONLY a protein capsid covering it, it is termed a NAKED CAPSID VIRUS or a NAKED VIRUS. However, some viruses pick up a lipid membrane from the host cell when it is released, that surrounds the capsid. The lipid membrane is called an ENVELOPE and such viruses are termed ENVELOPED VIRUSES. All virus contain one more important proteins type; this is ATTACHMENT PROTEIN or docking proteins. The attachment protein is needed by the virus to ATTACH TO ITS TARGET CELL before it can enter that cell. Obviously this attachment protein must lie on the outer surface of the virus so that it is available to contact the appropriate RECEPTOR SITES on the target host cells. These attachment proteins are often called SPIKES because they can extend away from the cell so as to better be able to contact the host receptor (think of the viral AP & the host receptor site as being the pairs of a VELCRO SYSTEM). In addition, virus may contain small quantities of carbohydrate (glycoprotein). Example of these two morphological types are shown in the figures below.
![]()
|
|
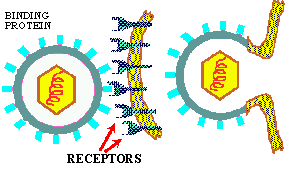
To understand this discussion you may wish to go back over the previous sections on enzyme specificity and substrate/enzyme complex formation that deal with the basic principle of ligand/receptor binding. All viruses have a LIMITED HOST RANGE and even within a host they attach to and invade only those cells with the APPROPRIATE RECEPTOR SITES. This specificity is the result of the same BASIC PRINCIPLE that controls all the other specific mechanisms of life: LIGAND/RECEPTOR BINDING SPECIFICITY; Figure at top of chapter. The following are examples of degrees of specificity:
![]() The smallpox and AIDS viruses only attack man (recently [Fall '95] the first
reports of a chimpanzee being infected with HIV was reported).
The smallpox and AIDS viruses only attack man (recently [Fall '95] the first
reports of a chimpanzee being infected with HIV was reported).
![]() The bacteriophage lambda only attacks cells which contain the receptor for
binding the sugar maltose.
The bacteriophage lambda only attacks cells which contain the receptor for
binding the sugar maltose.
![]() The influenza virus can live in ducks, chickens, wild birds, pigs
and humans.
The influenza virus can live in ducks, chickens, wild birds, pigs
and humans.
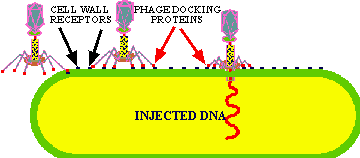
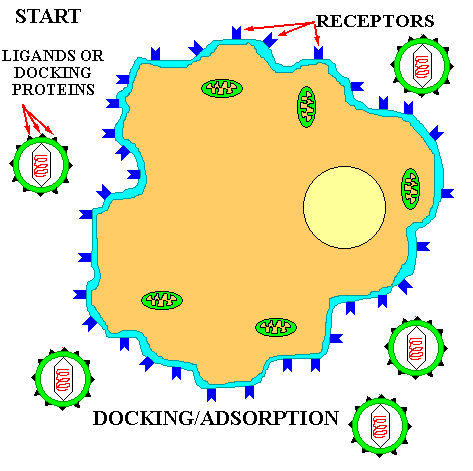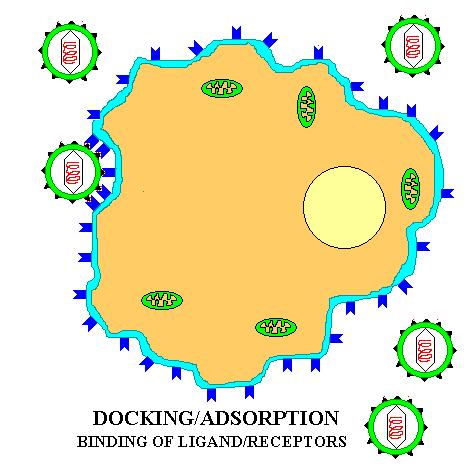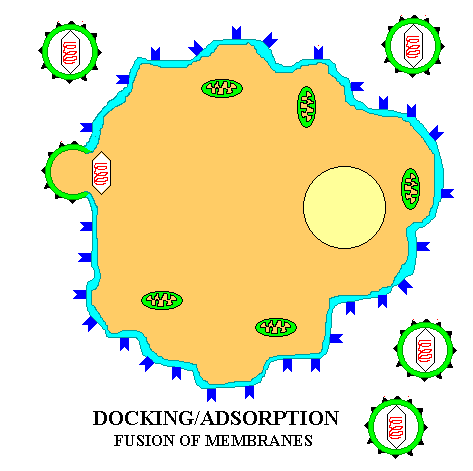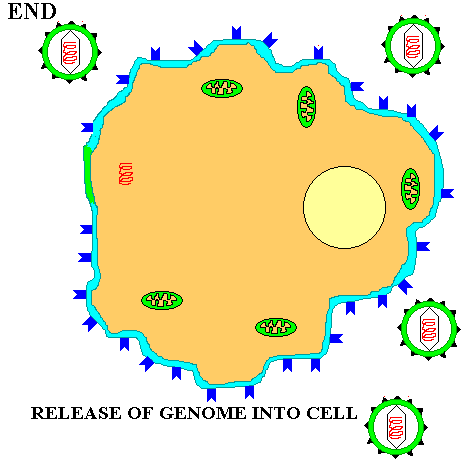
The basic principle of viral infection of a given cell is that if a cell's OUTER SURFACE contains the RECEPTOR to which a viruses' ATTACHMENT PROTEIN (docking proteins) can BIND, the virus will be able to invade and grow in that cell. This only applies within RELATED SPECIES or to different types of cells within a host. For example, the lambda phage will only naturally invade a few species of Gram negative bacteria, but if you put a plasmid in a G- cell that contains the gene for the lambda receptor, then lambda can bind to and inject its DNA into that cell (see below). Another example is the COMMON COLD VIRUSES. These viruses are specific for the human UPPER RESPIRATORY TRACK because they contain the appropriate AP for binding to the receptors on the cell membrane of the specialized cells in the upper respiratory tract. Other viruses that cause intestinal diseases only bind to certain receptors of cells in the intestine; others bind only to liver cell-receptors and cause hepatitis; while others like herpes only bind to receptors on and live in nerve cells.
![]()
The general life cycle (REPLICATION) of all viruses is exemplified by that of the bacteriophage. The following five steps, in order, are involved in phage (virus) replication:
|
|
|
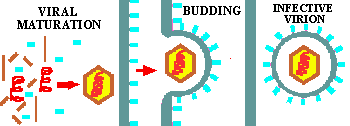
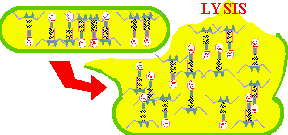 Figure 6C. Phage Maturation and Release. The various components accumulate within the cell and begin to SPONTANEOUSLY ASSEMBLE into new phage. However, the process is an orderly one so that each component is added in the proper sequence. Enzymes are produced that cause the cell to burst or lyse, releasing a crop of new viruses into the environment. In a properly set-up experiment, you can watch the cloudy culture clear up within a few seconds almost as if by magic. This site has another illustration of the lytic growth of phage. |
![]() Adsorption or DOCKING
with the host receptor protein.
Adsorption or DOCKING
with the host receptor protein.
![]() Entry or PENETRATION
of the viral nucleic acid into the host cytoplasm.
Entry or PENETRATION
of the viral nucleic acid into the host cytoplasm.
![]() BIOSYNTHESIS
of the viral components.
BIOSYNTHESIS
of the viral components.
![]() Assembly (MATURATION)
of the viral components into complete viral units.
Assembly (MATURATION)
of the viral components into complete viral units.
![]() RELEASE of
the completed virus from the host cell.
RELEASE of
the completed virus from the host cell.
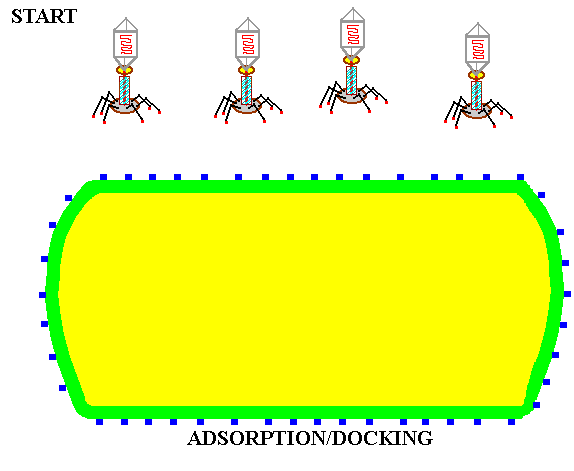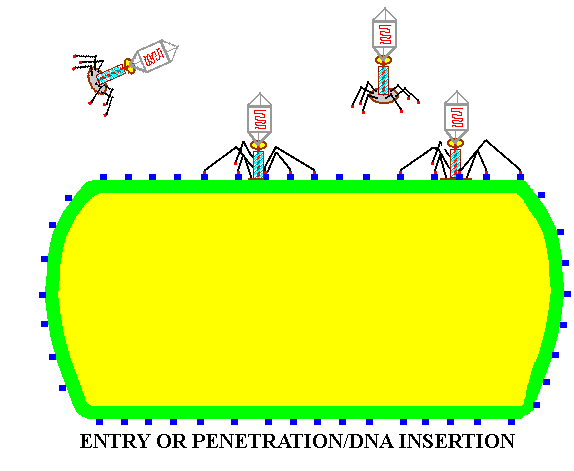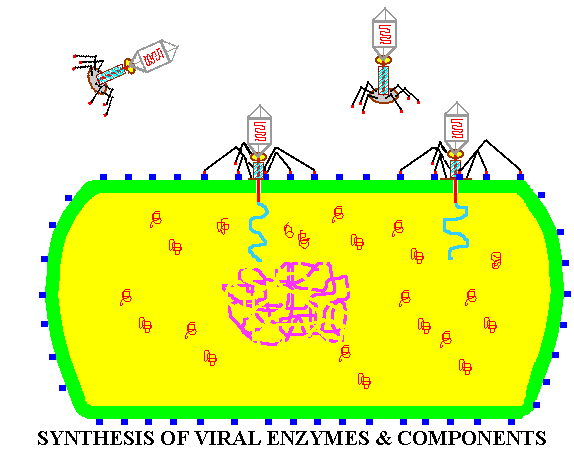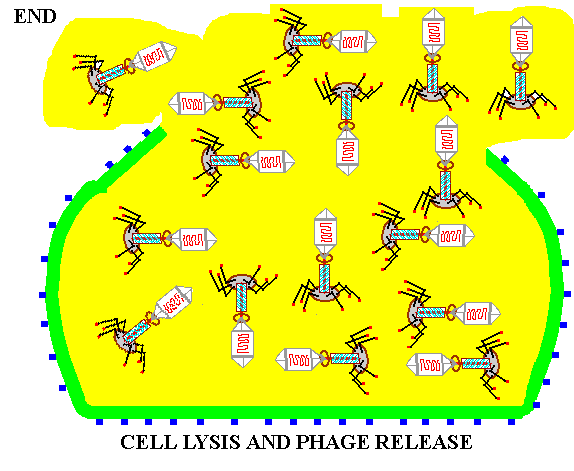
Summary animation of the life cycle of viruses as represented by a bacteriophage.
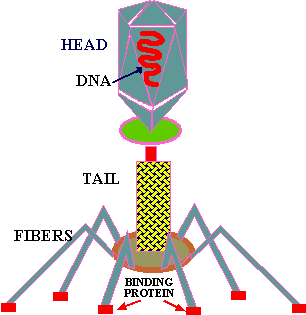
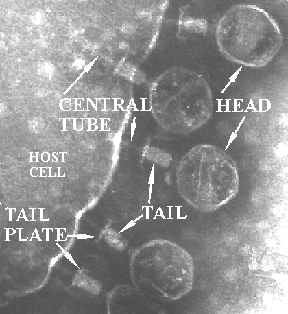
Some bacteriophage are among the larger, more complex viruses. The illustrations above (Fig. 5-8) covers the life cycle of one of these, the T-even phage. These phage have a head, a tail, a tail plate and tail fibers as seen in the Fig. 2
In the absence of a host, the long tail fibers are WOUND AROUND THE TAIL, but when the presence of a suitable host is sensed by the phage the tail fibers UNWIND and extend out from the tail plate, thereby increasing the area of potential contact with the host. The tip of the tail fiber contains the DOCKING PROTEIN (Velcro hooks). When one or more of these adsorption proteins contacts a RECEPTOR PROTEIN (Velcro loops) on the host, it binds tightly to it, bringing the remaining tail fibers quickly into contact with other receptor proteins on the host and firmly ATTACHING THE VIRUS TAIL to the host outer wall.
This attachment sets in motion a series of events that leads to phage lysozyme eating a hole in the cell wall. Then the tail fibers contract forcing the tail through the hole into the cytoplasm. Following this the DNA from the head is injected into the host cytoplasm.
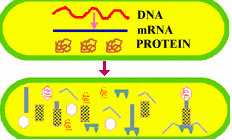
Once in the host's cytoplasm the viral DNA SEIZES CONTROL. It produces a special nuclease that hacks the host's DNA into little, ineffectual fragments (a sort of Freddy Krugar enzyme), thus leaving the cells machinery without a BRAIN. The viral DNA (or brain/CPU), then produces (#transcription & translation) a horde of enzymes and proteins which use the ribosomes, raw materials and energy sources of the BRAIN-DEAD host to make new virus.
These enzymes MANUFACTURE new phage components in large quantities which pile up in the cytoplasm. Subsequently, proteins are formed which assist in the maturation process. Many of the phage components are SELF ASSEMBLING in that they are made so as to SPONTANEOUSLY assemble into certain PREDETERMINED forms or shapes, like the head, tail, tail fibers etc.
Once the MATURE PHAGE are completely assembled and ready to begin another life cycle, the cell is lysed or ruptured and the mature phage are released into the environment; imagine a balloon filled with Jell-O dropping on someone's head!
![]()
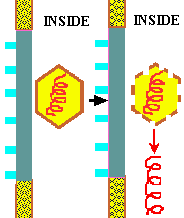
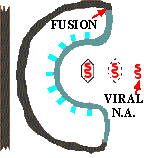
The life cycle of ENVELOPED EUKARYOTIC VIRUSES is basically the same as that of bacteriophage. Adsorption begins with the DOCKING PROTEINS of the virus locking into SPECIFIC RECEPTOR SITES on the surface of the target host cells(See first figure). The details of entry are somewhat more complex in the case of enveloped eukaryotic viruses as two distinct mechanisms are employed for entering the host cell. Rather than having the protein coat of the virus stay outside the cell, as is the case with phage, the CAPSID UNIT enters the cytoplasm by one of TWO BASIC MECHANISMS. In the case of some enveloped eukaryotic viruses the viral membrane FUSES with the cell membrane and in effect opens up so as to allow the virus to enter the cell cytoplasm. In another case the cell ENGULFS the entire enveloped eukaryotic virus, taking it into the cytoplasm. The former case will be described first.
Summary animation of entry of a eukaryotic virus into a host cell by fusion
Once in the cytoplasm the protein capsid is removed and the viral genome is freed into the cytoplasm. Depending on the nature of the virus the viral genome may stay in the cytoplasm or it may enter the nucleus before beginning the next stage of the life cycle.
|
|
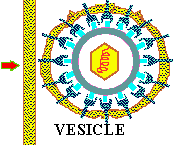
In the case of ENGULFMENT, or ENDOCYTOSIS, the initial adsorption process is the same, but rather than directly fusing with the cell membrane, the cell is stimulated to take-up or engulf the entire eukaryotic virus. This process is illustrated below.
Summary animation of the entry of a eukaryotic virus by engulfment. or endocytosis
Once inside the cytoplasm the enveloped eukaryotic virus is said to be in a VESICLE. Subsequently, the two lipid membrane of the vesicle fuse, releasing the capsid covered virus. The capsid is then removed and the viral genome is freed for the next stage of viral replication.
The actual processes from biosynthesis to maturation and release VARY WIDELY depending on the virus type. However, at the core, the viral genomic material takes over the cellular metabolism for its own selfish purposes. It PRODUCES mRNA which makes VIRAL PROTEINS. The viral proteins produce the various NEW VIRAL COMPONENTS. These components are assembled and the viral genome is PACKAGED in a protein capsid. Finally the mature virus or VIRIONS are released into the environment to begin the cycle again.
|
|
The naked virus replication is less elaborate, but follows the same course. After binding to the host receptors the naked viral capsid penetrates into the host cytoplasm, the genome is released and the steps of viral replication ensue. Figure 11 shows the maturation and release of an enveloped eukaryotic virus. Since the final release of enveloped eukaryotic viruses involves taking a portion of the cell membrane the process is called BUDDING. For example, the HIV virus buds out of the cell. Whether release involves lysis or budding, a cell eventually dies due to the viral infection and destruction of its metabolism.
To be accurate, a viral infection does not always lead to immediate cell death. In some cases a persistent infection may ensue in which viruses are released (leak) from the cell over long periods of time. In other cases a virus may cause the transformation of a cell and then disappear. Finally, some viruses lay DORMANT for a long time only to activated later. Do you know anyone who has had shingles; many of you reading this will have it. It is an example of a dormant virus being activated. Cold sores are another example of a virus that remains latent until something activates it (like the stress of finals).
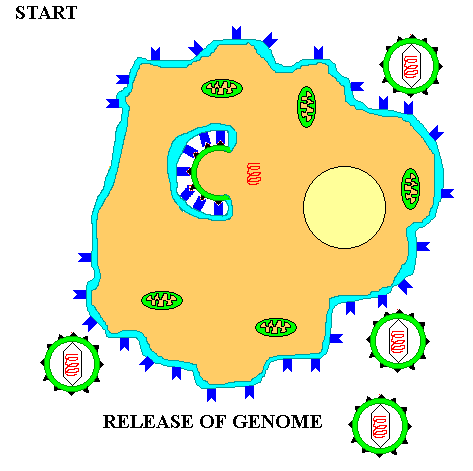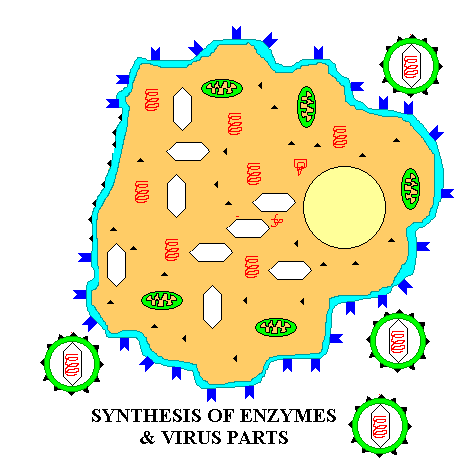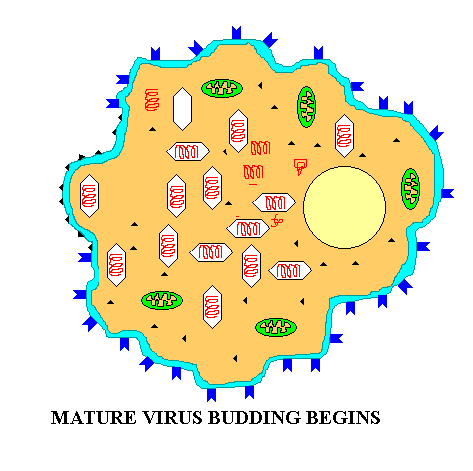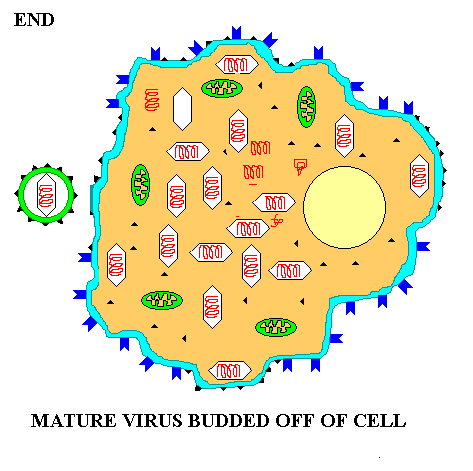
Summary animation of eukaryotic virus budding out of a cell (e.g. HIV)
As you will see viruses that use RNA as their genomic material present a unique problem in replication since the normal process of life is DNA to mRNA to protein. The discovery of how RNA viruses manage to replicate led to a Nobel Prize to the discovers and has given us important knowledge about the etiology of cancer, AIDS and other viral diseases.
For another detailed description of virus replication click here to see some nice, but rather advanced illustrations of what goes on during virus replication.
![]()
All viruses are studied using the general techniques described below.
![]() Find a SUITABLE HOST
cell that supports the growth of the virus under investigation.
Find a SUITABLE HOST
cell that supports the growth of the virus under investigation.
![]() Grow that host as a single layer of cells (MONOLAYER)
on a solid surface, like a bacterial plate.
Grow that host as a single layer of cells (MONOLAYER)
on a solid surface, like a bacterial plate.
![]() Infect that host with the virus of interest; adjusting the concentration of
virus so that most cells are infected by only ONE
VIRION (a
complete, infective virus).
Infect that host with the virus of interest; adjusting the concentration of
virus so that most cells are infected by only ONE
VIRION (a
complete, infective virus).
|
|
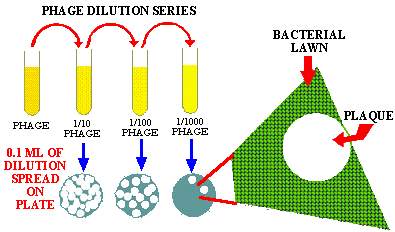
As the viruses replicate and are released, they SPREAD IN A CONCENTRIC CIRCLE and infect the nearby cells. This CIRCLE-OF-DEATH produces a hole or PLAQUE in a LAWN of living cells. These plaques can be easily seen and counted so that the numbers of virus can be quantitated. This is like a person with AIDS infecting everyone they have intercourse with, with the HIV virus, thus producing their own "CIRCLE or HUMAN PLAQUE-of-DEATH".
Studying phage this way is relatively easy and inexpensive, but just the opposite is true for most eukaryotic viruses. In general, eukaryotic cells are DIFFICULT TO GROW. They grow SLOWLY, require special, often EXPENSIVE MEDIUM, and SPECIAL EQUIPMENT. Finally, we still are not able to cultivate many eukaryotic cells or to grow many important viruses.
![]()
|
Major viral disease treatment
factoids: |
The obligate intracellular parasitic nature of viruses makes it very difficult to treat them. Because viral reproduction occurs only inside living cells and mostly uses the host cell’s own metabolic machinery to generate new virions, it is a formidable task to find or develop drugs that are both (a) able to penetrate the cell’s cytoplasmic membrane and (b) to selectively damage ONLY viral components. For example, drugs that disrupt a virus’ protein or nucleic acid synthesis as well as the host’s system are an unacceptable option. How then can viral infections be dealt with?
The classical, and still most effective, weapon in the war against viruses, immunization, is routed in the evolutionary struggle between the host and the viral pathogen. Vaccination, by arousing a host’s evolved immunological defense against foreign antigens (e.g. specific viruses), usually prevents infection. Vaccination has eliminated smallpox as a human disease and we are now attempting to do the same thing to polio and measles. This "extermination strategy" works optimally on viruses that are limited to Homo sapiens.
Many viruses however survive in non-hominid reservoirs (e.g. influenza &, hantavirus). In these cases the strategy depends on the reservoir(s) and mechanism(s) of dissemination to humans. The most common strategies used in this continuous battle are:
Immunization of hosts with dead or attenuated virus (e.g. flu, polio) or
with genetically engineered (G.E.) antigens.
| Immunization of hosts using cloned viral DNA shot directly into host
cells.
|
Immunization of hosts using foods containing cloned viral genes that
produce viral antigen(s).
|
Immunization of humans and alternate hosts (e.g. pets and vets against
rabies).
| Minimize contact between humans and the natural reservoir(s) (e.g.
Hantavirus & mice).
| Minimize vector/human contact (e.g. mosquito/yellow fever). | |
As our knowledge of virus genetics has grown we are developing molecular biological strategies for dealing with viruses. While none of these yet can be said to cure an established viral infection, they can, by minimize the virus load in the infected individual, prevent or decrease the spread of the virus to new hosts (e.g. HIV from mother to fetus) and its damage to an infected host. These approaches include:
The use of analogues that inhibit crucial viral enzymes.
The use of agents that block infection
|
The use of agents that stimulate or enhance the efficacy of the host’s
immune system.
|
|
Physicians are beginning to cautiously talk of a "CURE" for HIV through the use of combinations of the above treatments. The idea being that if the combined attack can lower the virion concentration sufficiently then the body’s natural immunity can "clean up" the remaining virus and render the host virus free. Predictions about such an outcome are chancy, but they are not as totally unreasonable to consider as they were (in 1997).
![]()
Some neurological diseases are caused by protein infectious particles (PRIONS). These include several animal and at least 3 human diseases. One of these diseases, KURU, infects its victims when they eat the brain tissue of their enemies (a questionable activity at best). The best studied of these diseases is scrapies in sheep. The disease entity seems to be composed completely of PROTEIN and to entirely lack any nucleic acid. This poses a major problem given the significant role of DNA and RNA in life. Three theories are currently being considered to explain prions.
The MAD COW disease that was first detected in England and parts of Europe a few years ago is apparently a new prion disease and it has caused the use of beef in Britain to fall precipitously. Thousands of cattle have been slaughtered and their carcasses destroyed to prevent the spread of this disease. At least 3 farm workers have died from a disease with symptoms like those of the Mad Cow disease. If you're interested in learning more about this disease ask me in class click here for URL on the subject; click here for more on Mad Cow Disease. For more information on prions click here.
This is a case of an Emerging Disease about which we understand too little to know whether to be scared-out-of-our-wits or just to be wary and concerned. The Press/TV/Tabloids find prions a good way to sell their services and they tend to hype it up for that purpose. However, there are scientists who are very concerned about the potential dangers of prions. The following are some points of information (tentative) to keep in mind:
![]()
Plants fall victim to agents composed of NAKED RNA that are only 300 to 400 nucleotides long, called VIROIDS. The evidence is conclusive that viroids cause plant diseases, but the mechanism of pathogenicity is not known. So far NO HUMAN viroids have been discovered, but it is considered a real possibility that they exist.
![]()
http://www-micro.msb.le.ac.uk/335/BS335.html; Virology course; advanced for 101, but loads of great information and excellent drawings (I'm envious).
http://www.blc.arizona.edu/INTERACTIVE/virusL2.95/virus.html; Very good pictures
http://www.uct.ac.za/depts/mmi/stannard/virarch.html; Home pg. of virus morphology, explains theory
Copyright © Dr. R. E. Hurlbert, 1999.
This material may be used for educational purposes only and may not be
duplicated for commercial purposes.