TECIDOS E ESTRUTURAS VEGETAIS
Sumário:
Colênquima (Tecidos de Suporte)
Esclerênquima (Tecidos de Suporte)
Tecidos de Revestimento e Protecção
Raiz Secundária de uma Gimnospérmica
Raiz Secundária de uma Dicotiledónea
Caule Primário de uma Monocotiledónea
Caule
Primário de uma Dicotiledónea
Caule Secundário de uma Gimnospérmica
Caule
Secundário de uma Dicotiledónea
Uma vez constituído o ovo ou zigoto,
inicia-se o desenvolvimento embrionário, que inclui os processos de
crescimento, morfogénese e diferenciação celular. As estruturas que resultam
deste desenvolvimento correspondem a um plano estrutural e funcional característico.
As células agrupam-se e associam-se, constituído tecidos; os tecidos formam órgãos,
e estes, sistemas de órgãos. O resultado final de todo o processo é o
organismo.
1) Técnica de Genebrez
Para esta técnica é necessário fazer cortes
muito finos no material vegetal a observar, por exemplo em raízes de Iris
germanica, o lírio.
O material fixado em álcool é colocado num
pedaço de medula de sabugueiro e com o auxílio de uma lâmina de barbear,
fazem-se cortes finos.
Em alternativa pode usar-se um micrótomo mecânico
para executar os cortes.
Os cortes são depois colocados em água e
lavados.
Os cortes são depois introduzidos num
recipiente contendo água de Javel, durante 5 a 10 minutos, para destruir
o conteúdo celular, deixando opacas as paredes.
Em seguida lavam-se em água acética durante
5 minutos, para retirar o excesso de água de Javel.
Os cortes são depois retirados da água acética
e lavados em água durante mais 5 minutos.
A fase seguinte consiste na coloração pelo
corante de Genebrez, e dura cerca de 5 minutos.
Lavam-se em água durante 5 minutos, para
retirar o excesso de corante.
Os cortes são finalmente colocados numa lâmina
de vidro sobre uma gota de água glicerinada.
Com esta técnica as paredes celulósicas
coram de rosa-pálido, as paredes lenhificadas e suberificadas coram desde
amarelo a castanho-alaranjado.
O corante usado nesta técnica é constituído
por 9 partes de "carmim aluminado de Grenacher" e 1 parte de
verde iodo.
A técnica em si é muito semelhante à
descrita anteriormente, residindo a principal diferença no corante usado.
Com esta técnica as paredes celulósicas
coram de vermelho, as paredes lenhificadas coram de verde e as suberificadas
coram de amarelo acastanhado.
Tecidos Meristemáticos
Os primeiros tecidos que aparecem nos vegetais
são os meristemas. Os meristemas também são denominados tecidos de formação,
pois é a partir deles que se originam os tecidos definitivos das plantas.
Os meristemas, como tecidos embrionários que
são, têm como principal característica a capacidade permanente de divisão
das suas células. Estas são, em geral, de pequenas dimensões, isodiamétricas,
cúbicas ou um pouco alongadas, com citoplasma abundante, quase sem vacúolos,
com um núcleo relativamente grande e paredes finas.
Os meristemas podem resultar directamente das células do embrião e
denominam-se meristemas primários ou apicais. Ocupam os ápices vegetativos da raiz
ou do caule e são responsáveis pelo crescimento em comprimento de cada um
daqueles órgãos.
Os meristemas primários originam células que
se especializam em funções específicas e constituem tecidos definitivos primários.
Os meristemas podem também ser constituídos
por células que readquiriram a capacidade de divisão e denominam-se meristemas
secundários ou intercalares.
As células dos meristemas secundários são,
geralmente, mais alongadas que as dos meristemas primários e possuem grandes
vacúolos, o que nunca acontece com as células dos meristemas apicais. São
exemplos de meristemas secundários: o câmbio vascular, meristema secundário
responsável pela formação dos tecidos vasculares secundários (o xilema e o
floema secundários); e o felogénio, responsável pela formação do súber ou
cortiça.
Os meristemas secundários ou intercalares
situam-se entre os tecidos já formados e são responsáveis pelo crescimento em
diâmetro dos órgãos vegetais.
Tecidos Essencialmente
Elaboradores
Os tecidos essencialmente elaboradores são
constituídos, por parênquimas. Estes tecidos são constituídos por células
vivas.
As células do parênquima clorofilino
apresentam paredes finas de natureza celulósica. O seu citoplasma está,
geralmente, concentrado junto da membrana. Para além destas características,
este parênquima apresenta inúmeros cloroplastos.
Distinguem-se dois tipos de parênquima
clorofilino, conforme a forma e a colocação das células. Um, constituído por
células mais ou menos alongadas, dispostas lado a lado, sem espaços entre
elas, que pode ser observado na página superior de folhas de dicotiledóneas.
Este parênquima designa-se parênquima clorofilino em paliçada.
Outro, constituído por células mais ou menos
esféricas, com grandes lacunas entre si, que se pode observar na página
inferior das folhas de dicotiledóneas. Este parênquima designa-se parênquima
clorofilino lacunoso.
As folhas das plantas gimnospérmicas,
apresentam um tipo especial de parênquima clorofilino, pois não se apresenta
diferenciado em parênquima clorofilino lacunoso nem em paliçada. O parênquima
das folhas destas plantas designa-se por parênquima clorofilino encaixado.
O parênquima é o representante principal do
tecido fundamental e encontra-se em quase todos os órgãos da planta, formando
um tecido contínuo, como, por exemplo, no córtex e na medula do caule e da raiz.
As células possuem, em vez de cloroplastos,
leucoplastos, que elaboram produtos de reserva a partir dos alimentos elaborados
em maior quantidade que a necessária à célula.
Parênquima como o da batata, em que as células
armazenam amido em quantidade, denomina-se parênquima amiláceo.

Tecidos Essencialmente Mecânicos
Os tecidos de suporte são constituídos por células
que apresentam as paredes espessas. Encontram-se em várias zonas do organismo
vegetal, às quais conferem grande rigidez, impedindo, assim, o esmagamento das
células de paredes finas, quando ocorrem movimentos provocados pelo vento.
Permitem também, a posição erecta das plantas.
Os tecidos de suporte são de dois tipos: O
Colênquima e o Esclerênquima
O colênquima é um tecido em que as células
são vivas, mesmo na maturidade. Normalmente, as suas células são alongadas e
formam cordões nas regiões periféricas dos caules e pecíolos e rodeiam as
nervuras das folhas das dicotiledóneas.
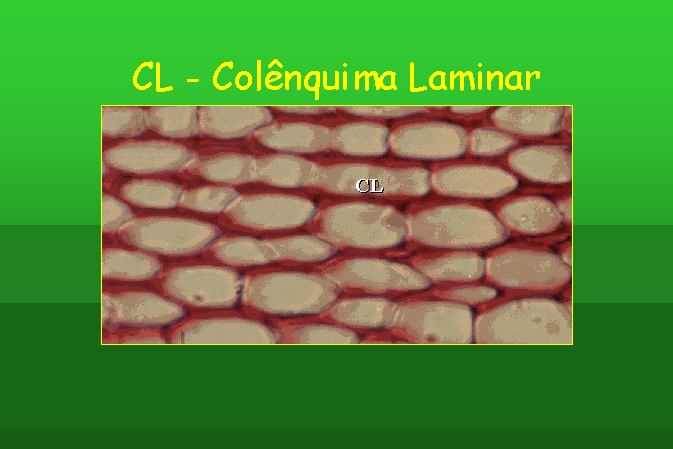
As paredes celulares das suas células são
desigualmente espessadas. Se as células apresentam espessamentos nos ângulos,
o colênquima chama-se colênquima angular ou aristal. Se o espessamento se
encontra nos ângulos, mas apresenta meatos ou lacunas, estamos na presença de
colênquima lacunar. Finalmente, se o espessamento surge nas paredes
tangenciais, o colênquima designa-se colênquima laminar.
O esclerênquima é um tecido constituído por
células desprovidas de conteúdo celular, isto é, mortas, apresentando as
paredes igualmente espessadas e quase sempre lenhificadas. A cavidade celular,
ou lúmen, vai diminuindo à medida que se efectua o espessamento e lenhificação
da parede.
O esclerênquima é um importante elemento de
resistência e suporte nas partes das plantas que terminaram o seu processo de
alongamento.
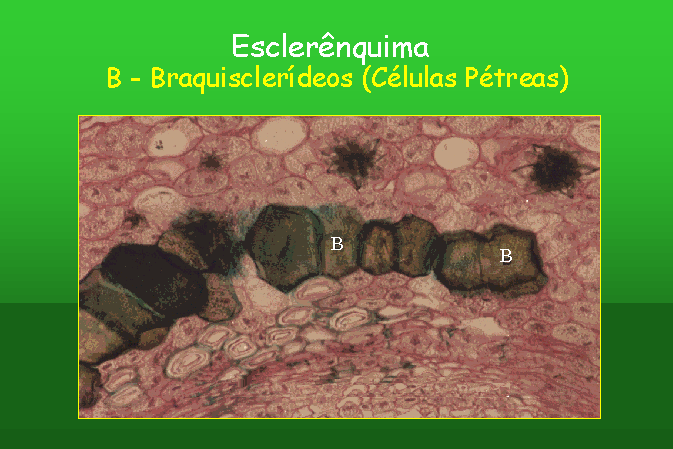
As células de esclerênquima são
habitualmente divididas em duas categorias: os esclerídeos e as fibras. A fibra
é uma célula longa e delgada, ao passo que os esclerídeos variam de uma forma
aproximadamente isodiamétrica, até uma forma consideravelmente isodiamétrica,
sendo alguns deles muito ramificados.
Os esclerídeos têm diferentes denominações, consoante a sua morfologia: Tricosclerídeos; Macrosclerídeos; Osteoclerídeos. Na folha da Oliveira (Olea europea) podem-se observar esclerídeos filiformes; Braquisclerídeos ou células pétreas; Astrosclerídeos; fibras corticais; e finalmente Fibras Perivasculares, que rodeiam o feixes vasculares do caule da planta do milho (Zea mays).
2) Tecidos de Protecção ou Revestimento
As flores, frutos e semente, bem como os
caules e raízes, antes de engrossarem, são revestidos por um tecido de protecção:
a epiderme. A epiderme resulta da actividade da protoderme e as células que a
constituem, sob o ponto de vista funcional e estrutural, podem apresentar grande
variabilidade.
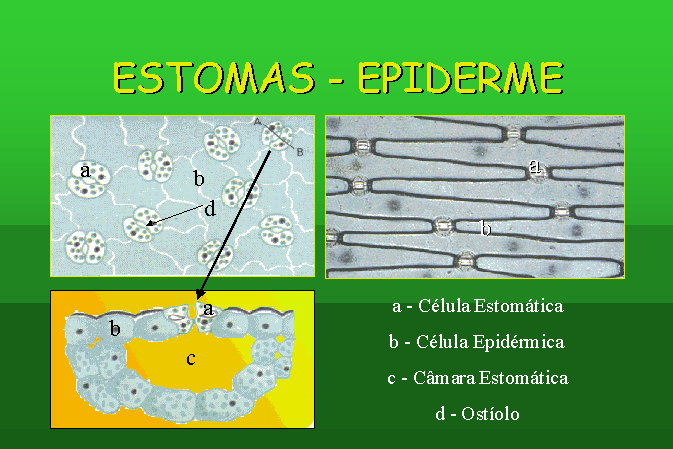
Na epiderme da maioria dos órgãos aéreos,
principalmente nas folhas, existem estomas. os estomas são formações que
permitem a realização fácil da troca de gases entre a planta e a atmosfera.
cada estoma é constituído, essencialmente, por duas células em forma de rim,
sempre com cloroplastos, chamadas células estomáticas, que delimitam uma abertura,
denominada ostíolo. Além das células estomáticas, o estoma possui outras células
anexas, formando, assim o aparelho estomático.
Nos casos típicos, a epiderme é constituída
por uma única camada de células vivas. As células epidérmicas não
apresentam meatos entre si, encontrando-se firmemente unidas, o que confere uma
significativa protecção mecânica ao órgão da planta. Não apresentam
cloroplastos. Nas partes aéreas, a parede externa das células epidérmica
apresenta cutina, constituindo, no seu conjunto, a cutícula, diminuindo assim a
perda de água.
Quando a camada pilosa da raiz deixa de
apresentar actividade, suberificam-se as paredes das células da epiderme e
mesmo das camadas subjacentes. estas células, assim modificadas, protegem com
mais eficácia os tecidos subjacentes. É o que podemos verificar, por exemplo,
nas raízes velhas do lírio. O conjunto destas células suberificadas
constituem a exoderme.
A endoderme é um tecido constituído por células
vivas, que se encontra na generalidade das raízes a separar zonas internas.
As células endodérmicas são prismáticas,
unidas entre si, sem qulquer espaço. Estas células têm espessamentos nas
membranas laterais denominados por pontuações de Caspary ou, em
alternativa, espessamentos em forma de "U".
O súber é um tecido secundário, pois
resulta da actividade de um meristema secundário chamado felogénio ou câmbio
subero-felodérmico. este tecido de protecção forma-se nas partes mais velhas
do vegetal. É um tecido constituído por células mortas, reduzidas à parede
suberificada. O súber ou cortiça pode atingir espessura significativa, como
acontece nos sobreiros. A suberina, que entra, em grande percentagem, na
constituição da parede celular destas células, torna-as impermeáveis aos líquidos
e aos gases. Em consequência da formação contínua de súber, desprovido de
espaços intercelulares, os estomas são substituídos por lentículas. As lentículas
são constituídas por células suberificadas, que deixam espaços entre si,
permitindo a passagem dos gases entre o meio e os órgãos internos.
As plantas vasculares são constantemente percorridas por duas correntes de líquidos. Uma, da raiz para as folhas, constituída por uma solução aquosa de sais minerais extraída do solo. Outra, que circula das folhas para a raiz, constituída por uma solução de matérias orgânicas resultantes das sínteses ocorridas nos tecidos elaboradores, e que fornece às células as substâncias orgânicas de que necessitam. Estas duas correntes ocorrem num sistema de vasos condutores constituído pelo tecido lenhoso e tecido liberino.
O tecido lenhoso, também chamado xilema ou
lenho, é constituído por células mortas, alongadas, dispostas topo a topo em
séries longitudinais, formando colunas contínuas ou tubos, denominados vasos.
A parede celular apresenta zonas mais ou menos lenhificadas, o que confere aos
vasos o aspecto ornamentado característico. O xilema é o principal condutor de
água e sais minerais das plantas vasculares. A circulação realiza-se da raiz
para as folhas.
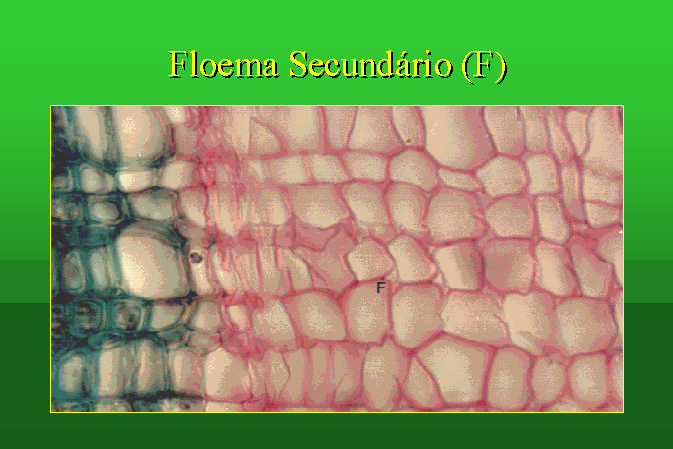
O tecido liberino, também designado floema ou
líber, é o tecido condutor de seiva elaborada. É constituído por células
vivas dispostas topo a topo, ligadas entre si por perfurações ou poros abertos
nas paredes contíguas, formando uma placa crivosa. Os tubos crivosos estão
associadas a células parenquimatosas especializadas, denominadas células
companheiras.
Estrutura da raiz
Na maioria das plantas vasculares, as raízes
constituem a porção subterrânea responsável pela fixação e absorção. Além
destas, desempenham ainda funções de armazenamento e condução de substâncias.
A primeira raiz da planta tem origem no embrião
e denomina-se, geralmente, raiz primária. Nas gimnospérmicas e dicotiledóneas,
esta raiz, também denominada raiz axial, cresce em direcção ao solo,
originando raízes secundárias ou laterais ao longo do seu eixo.
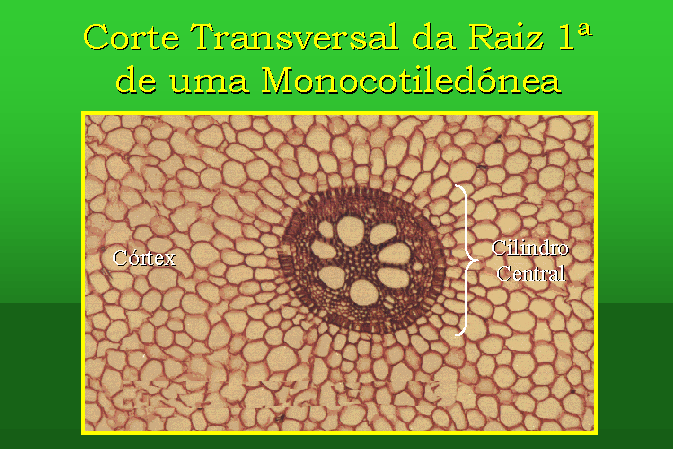
Estrutura Primária da Raiz de uma Monocotiledónea
Um corte transversal, efectuado na zona pilosa
da raiz de Iris germanica (Lírio), mostra-nos a sua estrutura primária,
que, geralmente, compreende as seguintes porções:
- A zona epidérmica, normalmente constituída
pela exoderme, cujas células se encontram suberificadas.
- A zona cortical, ou córtex, é constituída
por parênquima cortical, tecido de reserva que ocupa a maior área da estrutura
primária da raiz.
As células corticais apresentam meatos e,
também, numerosos contactos entre si. Os seus protoplasmas encontram-se ligados
por plasmodemos. Em consequência disso, as substâncias que transitam pelo córtex
podem atravessar as células.
A camada mais interna da zona cortical, a
endoderme, dispõe-se de modo compacto e não apresenta meatos. Estas células
apresentam-se suberificadas lateral e internamente, e em corte transversal
apresentam-se sob a forma de um "U".
Finalmente, o Cilindro Central, está limitado
por uma camada de células parenquimatosas, designada periciclo. Na parte
central encontra-se um tecido constituído por células vivas, denominado parênquima
cortical. Entre o parênquima cortical e o periciclo distribuem-se os tecidos
condutores, que se encontram dispostos radialmente e alternando o floema (corado
de vermelho) e o xilema (corado de verde). Os feixes condutores são simples e
alternos. O xilema primário é constituído na parte periférica por vasos de
pequeno calibre (o protoxilema), e na parte interna por vasos de grande calibre
(o metaxilema). Os feixes de xilema primário apresentam, na raiz,
desenvolvimento centrípeto. Nas monocotiledóneas o número de feixes
vasculares é superior a seis.
Estrutura Primária da Raiz de uma Dicotiledónea
Tal como na raiz primária da Iris
germanica (Lírio), a raiz de Ranunculus apresenta três zonas
distintas:
Uma zona epidérmica, constituída por uma única
camada de células epidérmicas e, por vezes, por uma exoderme, nas zonas mais
velhas da raiz.
A zona cortical é constituída por parênquima
de reserva, sendo delimitada internamente pela endoderme, cujas células
apresentam faixas de Caspary.
O cilindro central é delimitado pelo
periciclo e apresenta no seu interior parênquima medular e feixes alternos de
xilema e floema. O número de feixes nas dicotiledóneas é sempre inferior a
seis
Estrutura Secundária da Raiz de uma Gimnospérmica
Nas gimnospérmicas e dicotiledóneas, a
estrutura primária da raiz pouco tempo se conserva e a raiz engrossa,
aumentando de diâmetro. O engrossamento é assegurado pelo aparecimento de dois
meristemas secundários, cuja actividade origina tecidos definitivos secundários,
que modificam consideravelmente a estrutura da raiz.
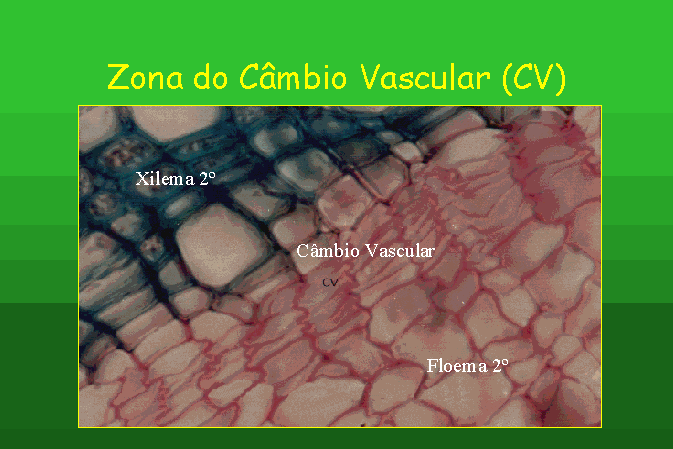
Um dos meristemas secundários é o câmbio
vascular, que origina, para o lado interno, lenho ou xilema secundário e, para
o lado externo, líber ou floema secundário.
O aumento de diâmetro do cilindro central da raiz,
devido à actividade do câmbio, faria rebentar a zona cortical. Contudo, tal não
acontece devido à formação, na zona cortical, de um outro meristema secundário,
o felogénio. O felogénio produz para o exterior o súber e para o interior a
feloderme. O súber constitui uma boa protecção para a raiz e a feloderme funciona
como tecido de reserva.
A raiz secundária de uma gimnospérmica, como
a do Pinus, apresenta raios medulares unisseriados e xilema secundário
homogéneo, ou seja, o tecido xilémico apresenta todas as suas células com
calibres idênticos.

Estrutura Secundária da Raiz de uma Dicotiledónea
A raiz secundária de uma dicotiledónea, como
por exemplo a de Vitis vinifera (Videira), apresenta, também, dois câmbios
secundários. O câmbio vascular que produz xilema e floema secundários e o
felogénio, que produz súber e feloderme.
A raiz secundária de uma dicotiledónea
apresenta raios medulares multisseriados e xilema secundário heterogéneo, ou
seja, o tecido xilémico apresenta células com diferentes calibres.
Estrutura Primária do Caule de uma Monocotiledónea
Se examinarmos um corte transversal do caule
de Zea mays (Milho), podemos distinguir as seguintes regiões, da
periferia para o centro.
A epiderme, camada de células de paredes
espessadas e cutinizadas.
A zona cortical ou córtex, constituído por células
vivas, de paredes finas, situado logo por baixo da epiderme e de espessura variável.
O cilindro central, bastante desenvolvido,
quase não apresentando delimitação relativamente à zona cortical. Os feixes
condutores duplos e colaterais distribuem-se de maneira irregular. Os feixes
libero-lenhosos encontra-se, neste caso, envolvidos por fibras de esclerênquima.
As monocotiledóneas não apresentam,
normalmente, crescimento secundário do caule.
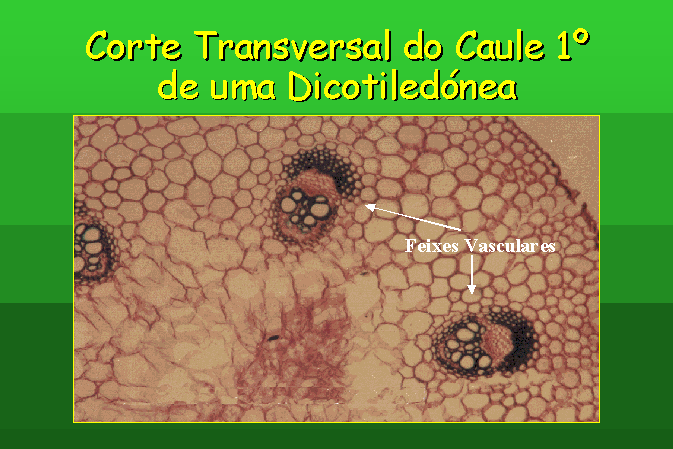
Estrutura Primária do Caule de uma Dicotiledónea
Se examinarmos um corte transversal do caule
jovem de Ranunculus, podemos destinguir as seguintes regiões, da
periferia para o centro:
Uma camada de células epidérmicas.
Uma zona cortical de extensão variável,
constituída por um parênquima cortical, formado por células pequenas de
parede fina e celulósica.
Nas células periféricas deste parênquima
encontram-se cloroplastos.
O cilindro central apresenta um parênquima
medular onde se destinguem feixes condutores duplos e colaterais, que apresentam
procâmbio entre o xilema e o floema.
O floema ocupa a parte externa e o xilema a
parte interna do feixes. No intervalo destes feixes e irradiando da medula ficam
os raios medulares.
O lenho apresenta o protoxilema voltado para
dentro e o metaxilema voltado para fora. Ou seja, ao contrário da raiz, o
xilema tem desenvolvimento centrífugo.
Estrutura Secundária do Caule de uma Gimnospérmica
Da mesma maneira que a raiz, o caule das
gimnospérmicas engrossa com a idade, desenvolvendo uma estrutura secundária,
por acção de meristemas laterais, respectivamente, o câmbio vascular e o
felogénio.
O câmbio vascular produz células que se
diferenciam em xilema secundário, para a parte interna, e floema secundário,
para a parte externa.
O felogénio produz súber ou cortiça para
fora e feloderme para dentro.
O xilema secundário de uma gimnospérmica,
como por exemplo a Ginkgo biloba, diz-se homogéneo, pois apresenta células
com calibres idênticos.
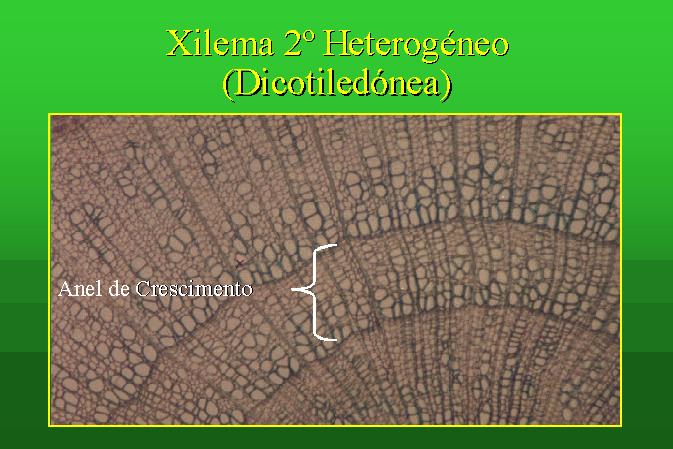
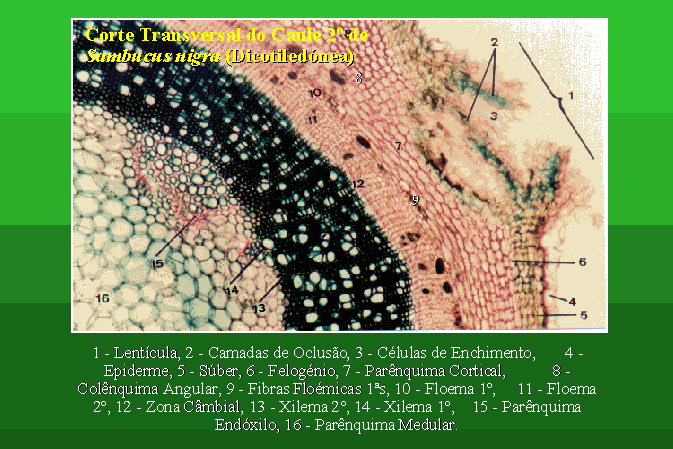
Estrutura Secundária de um Caule de uma Dicotiledónea
A estrutura secundária do caule de uma
dicotiledónea pode ser estudada, por exemplo, no corte transversal do caule de Sambucus
nigra.
Tal como nas gimnospérmicas, existe um câmbio
vascular e um câmbio subero-felodérmico, responsáveis pela formação dos
tecidos secundários.
O xilema secundário apresenta-se aqui, ao contrário do que se passa nas gimnospérmicas, com células de diferentes calibres, ou seja trata-se de um xilema heterogéneo.
Estrutura de uma Folha de Gimnospérmica
A morfologia da folha é muito variável. esta
variabilidade reflecte diferenças genéticas e de habitat, particularmente no
que diz respeito à disponibilidade de água.
A folha acicular do Pinus é um bom
exemplo disso, pois apresenta estruturas que visam diminuir as perdas de água
para o meio exterior.
As folhas aciculares do pinheiro apresentam
uma forte cutinização da epiderme, e os estomas estão localizados
profundamente, sob câmaras pré estomáticas, de forma a se reduzirem as perdas
de água por transpiração. A existência de uma hipoderme, constituída por células
lenhificadas, logo por baixo da epiderme, contribui, de forma significativa,
para evitar a evaporação de água.
As folhas de Pinus (Pinheiro)
apresentam ainda:
Canais resiníferos;
Parênquima clorofilino encaixado;
Endoderme com pontuações de Caspary;
Tecido de transfusão, situado na parte
interior da endoderme;
E feixes vasculares com crescimento secundário.
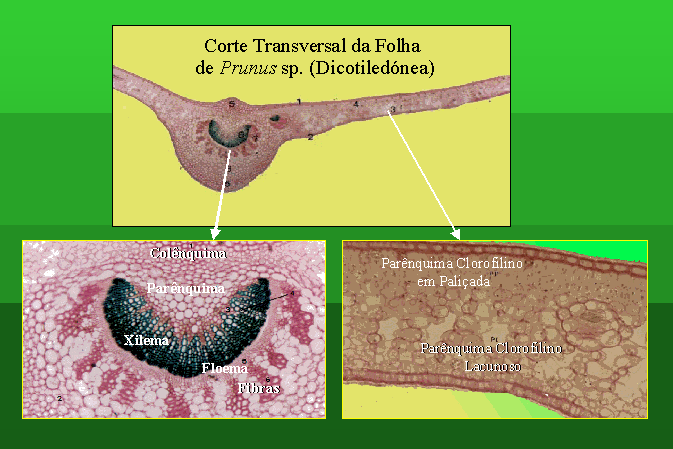
Estrutura da Folha de uma Dicotiledónea
Normalmente as folhas de dicotiledóneas
possuem uma porção expandida, ou limbo, e uma parte que liga o limbo ao caule
denominada pecíolo.
Independentemente da forma da sua forma e
estrutura, as folhas estão envolvidas nos processos da fotossíntese e da
transparição. Em qualquer destes processos os estomas desempenham um papel
muito importante.
Normalmente o número de estomas é maior na página
inferior que na página superior das folhas.
As folhas de Prunus apresentam dois
tipos de parênquima clorofilino. Na página superior surge-nos um parênquima
clorofilino em paliçada. A página inferior da folha apresenta parênquima
clorofilino lacunoso
A folha de uma dicotiledónea apresenta muitos
feixes vasculares, com uma nervura principal com crescimento secundário e inúmeras
nervuras laterais, com crescimento primário.
Estrutura da Folha de uma Monocotiledónea
Observando, ao microscópio, um corte
transversal da folha de Zea mays, encontramos, fazendo parte da sua
estrutura, os seguintes tecidos:
A Epiderme, constituída por uma camada de células
recobertas por cutina e por numerosos estomas dispostos em filas paralelas.
A zona ocupada, pelo parênquima clorofilino,
entre a epiderme das duas páginas, denomina-se mesófilo. Neste caso, e visto
que o parênquima clorofilino é idêntico tanto na página inferior como
superior, o mesófilo diz-se simétrico. O mesófilo da folha é percorrido pelo
floema e pelo xilema, que, conjuntamente com os tecidos de suporte,
fundamentalmente colênquima, constituem as nervuras. As nervação das
monocotiledóneas é paralelinérvea.
A folha da planta do milho (Zea mays),
aqui observada, apresenta também, células buliformes. Estas células motoras
estão envolvidas no mecanismo de enrolamento das folhas.
Texto baseado na videocassete didáctica Tecidos e Estruturas Vegetais
Uma Edição da
