INTRODUÇÃO
Para
se reproduzirem os organismos devem transmitir aos seus descendentes uma informação
genética semelhante àquela que possuem. Na verdade, a informação transmitida
não é igual pois uma das características da reprodução sexuada é assegurar
a variabilidade genética a qual, associada às mutações, é extremamente
importante em termos evolutivos.
O
ciclo de reprodução sexuada implica uma alternância entre a produção de células
haplóides (gâmetas), em que uma célula contém apenas um grupo de
cromossomas, e de células diplóides. A mistura de genomas
é assegurada pela fusão de duas células haplóides que originam uma célula
diplóide.
O mecanismo que, em muitos organismos, assegura a reprodução sexuada é
a meiose. Através da meiose, uma célula
diplóide origina quatro células células
haplóides evitando, deste modo, que aquando da fusão entre os gâmetas
o número de cromossomas sofra uma duplicação.
A
meiose consiste em duas divisões sucessivas do núcleo mas apenas uma divisão
de cromossomas. A primeira divisão é chamada reducional pelo facto do número
de cromossomas homólogos se reduzir a metade. Segue-se a segunda divisão,
chamada equacional, que mantém o número haplóide de cromossomas.
À semelhança do que acontece na mitose, em cada uma das duas divisões
da meiose podem distinguir-se quatro fases: profase, metafase, anafase e
telofase.
A primeira divisão meiótica começa por uma profase I muito longa e
complexa em que se podem considerar cinco sub-fases.
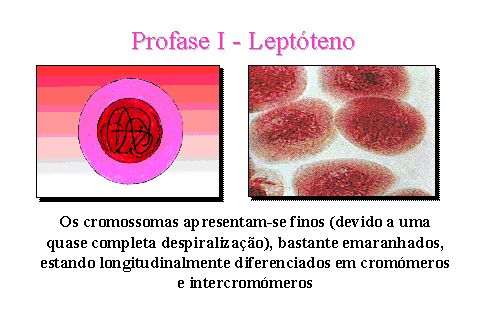
Leptóteno.
Os cromossomas apresentam-se muito finos devido a uma quase completa despiralização,
bastante emaranhados e longitudinalmente diferenciados em cromómeros e
intercromómeros. Embora a síntese de DNA já tenha ocorrido não é possível
observar os dois cromatídeos que constituem cada cromossoma.
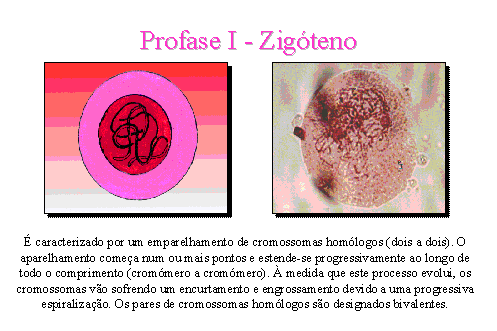
Zigóteno.
É caracterizado por um emparelhamento de cromossomas homólogos. O
emparelhamento começa num ou mais pontos do cromossoma e estende-se,
progressivamente, ao longo de todo o comprimento (cromómero a cromómero). Este
processo de emparelhamento é denominado sinapse. À medida que este
processo evolui, os cromossomas vão sofrendo um encurtamento e engrossamento
devido a uma progressiva espiralização. Os pares de cromossomas homólogos são
designados bivalentes.
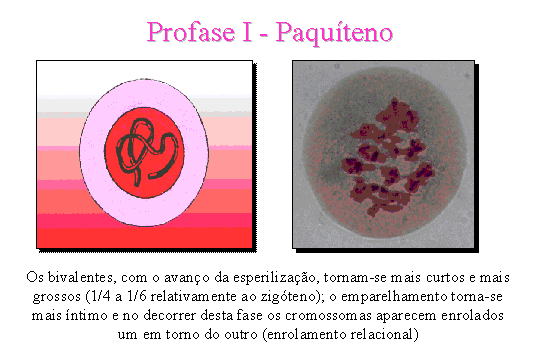
Paquíteno.
Os bivalentes, com o processo de espiralização, tornam-se mais curtos e mais
grossos. O emparelhamento torna-se mais acentuado e no decorrer desta fase os
cromossomas aparecem enrolados um em torno do outro.
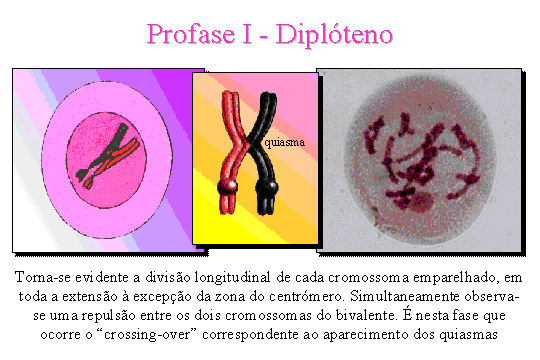
Diplóteno. Torna-se
evidente a divisão longitudinal de cada cromossoma emparelhado em toda a extensão à excepção da
zona do centrómero. Simultaneamente observa-se uma repulsão entre os dois
cromossomas constituintes do bivalente. É
nesta fase que tem lugar o mecanismo do crossing-over que consiste
numa troca de cromatídeos entre
cromossomas homólogos.
O
fenómeno do crossing-over implica, como é evidente, quebra e reunião
de cromatídeos em pontos estritamente homólogos. O equivalente citológico do
crossing-over é o aparecimento de quiasmas (pl. chiasmata). Um quiasma
resulta do facto dos cromatídeos se cruzarem nos pontos onde houve quebra e
reunião. O número de chiasmata é variável, podendo existir apenas um
ou vários.
Como
nesta fase cada bivalente é constituído por dois pares de cromatídeos recebe
o nome de tétrada cromossómica.
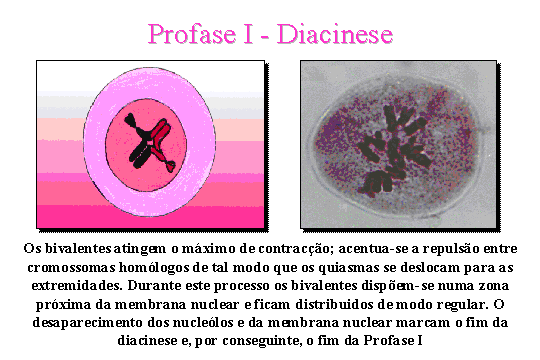
Diacinese. A distinção
entre o diplóteno e a diacinese não é muito clara, embora a diacinese seja
caracterizada por uma maior contracção dos cromossomas bivalentes. Além disso
verifica-se uma maior dispersão dos bivalentes na célula (fácil determinação
do número cromossómico) em virtude do desaparecimento da membrana nuclear.
Ocorre também o desaparecimento do nucléolo. Acentua-se a repulsão entre os
cromossomas homólogos de tal modo que os chiasmata se deslocam para as
extremidades (terminalização dos chiasmata).
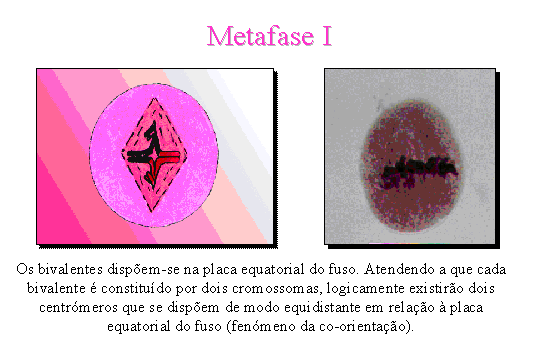
Metafase
I
.
Os bivalentes dispõem-se na placa equatorial da célula. Atendendo a que cada
bivalente é constituído por dois cromossomas, logicamente existirão dois
centrómeros que se dispõem de modo equidistante em relação à placa
equatorial.
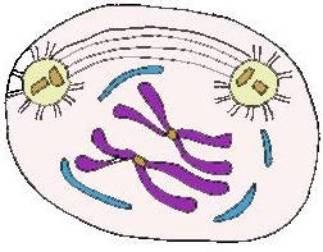
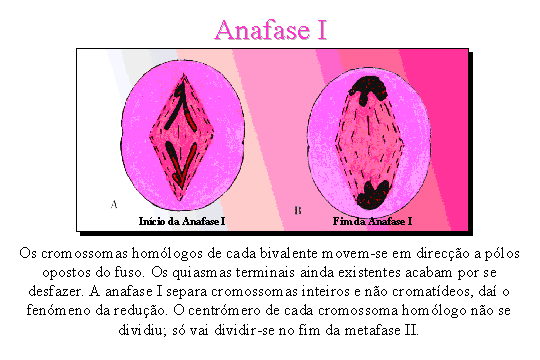
Anafase I . Os
cromossomas homólogos separam-se e cada um vai para um dos pólos da célula. O
movimento dos cromossomas é feito ao acaso, ou seja, cada cromossoma pode
deslocar-se para qualquer dos pólos.
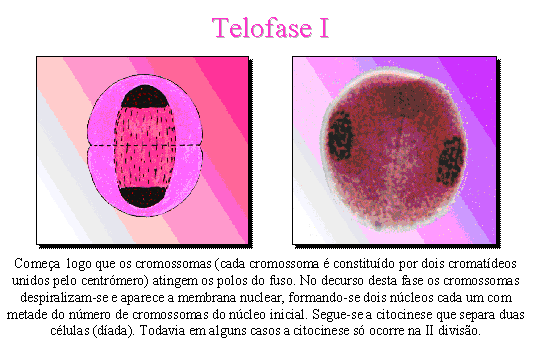
Telofase I .
Começa logo que os cromossomas (2 cromatídeos) atingem os pólos do fuso. No
decurso desta fase os cromossomas despiralizam-se e aparece a membrana nuclear,
formando-se dois núcleos cada um com metade do número de cromossomas do núcleo
inicial. Segue-se uma citocinese que individualiza duas células. Todavia, em
algumas espécies a citocinese ocorre apenas após a telofase II.
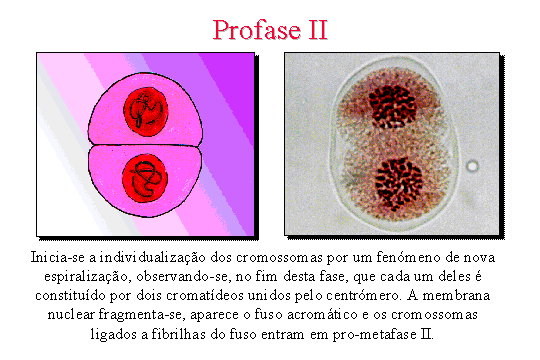
Profase II . Dá-se
a individualização dos cromossomas por um fenómeno de nova espiralização,
observando-se no fim desta fase que cada um dos cromossomas é
constituído por dois cromatídeos unidos pelo centrómero. A membrana
nuclear fragmenta-se, aparece o fuso acromático e os cromossomas ligados pelas
fibrilhas do fuso, entram em metafase.
Metafase II . Os
cromossomas dispõem-se pelos centrómeros na região equatorial do fuso, tal
como numa mitose vulgar.

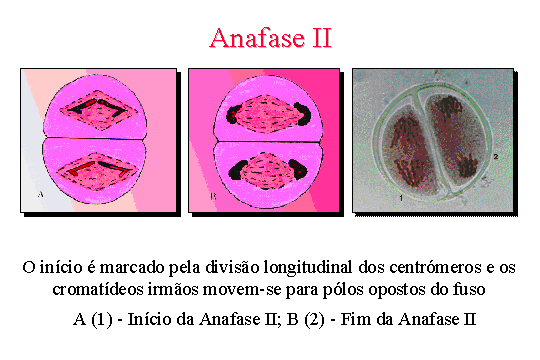
Anafase II . Dividem-se
os centrómeros e os cromatídeos irmãos movem-se para pólos opostos do fuso.
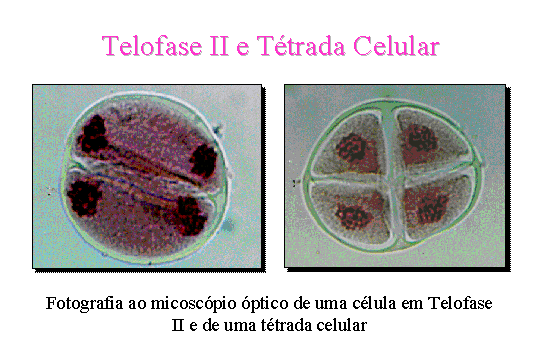
Telofase II . Os cromatídeos
despiralizam-se e aparece a membrana nuclear
e os núcleolos. Segue-se a citocinese obtendo-se assim quatro células
com metade do número de cromossomas da célula inicial.

O
processo da meiose pode ser estudado, com relativa facilidade na
formação dos grãos de pólen.
1)
Observação
de preparações definitivas de meiose obtidas pela técnica do carmim-acético.
2) Técnica
do carmim-acético para coloração de cromossomas nas células mães dos grãos
de pólen.
a) Fixar
as anteras em álcool-acético ( 3:1 ) durante 12 - 24h. O material pode ser
conservado durante meses em álcool a 70% a O-4ºC.
b) Transferir
uma antera para uma lâmina.
c) Adicionar
2 - 3 gotas de carmim acético.
d) Esmagar
a antera com uma agulha.
e) Esperar
cerca de 5 min. Retirar os detritos.
f) Aquecer
ligeiramente à chama e colocar uma lamela sobre o material.
g) Absorver
o excesso de corante com papel de filtro
e observar em imersão.
3) Observação
de cortes transversais de anteras e de ovários em preparações definitivas.
4) Observação
de tubos polínicos pela técnica do carmim-acético.
a) Numa
caixa de Petri contendo um
meio de cultura apropriado ( 3% sacarose, 1% agar ) colocar grãos de pólen
maduros.
b) Após
3 - 4h de cultura retirar um pedaço de meio ( 1 cm2 ) e colocar sobre uma lâmina
de microscópio.
c) Adicionar
2 - 3 gotas de carmim acético.
d) Aquecer
até o agar derreter.
e) Colocar
uma lamela e observar ao microscópio.
BIBLIOGRAFIA:
n
Amabis, J. M. e Martho, G. R., 1985 - Curso Básico de Biologia vol. 1,
Editora Moderna Ltd, cap. 9.
n
Darlington, C. D. e La Cour, L. F., 1969 - The Handling of Chromosomes.
George Allen & Unwin, 5ª ed.
n
Novikoff,
A. B. e Holtzman, E., 1977 - Células e estrutura Celular. Interamericana,
2ª ed. P, 278 - 289.
n
Raven, P. H., Evert, R. F. e Curtis, H., 1981 - Biology of Plants. Worth Publishers, Inc., 3ª ed., p 134 - 139.
n
Stahl, F., 1987 - Pourquoi nous différons de nos parents? Pour la
Science, 147:18-31.
n
Strasburguer, E. et al, 1981 - Tratado de Botânica. Editorial Marin, 6ª
ed., p, 32-39.
n
Strickberger, M. W., 1974 - Genética. Ediciones Omega, S.A..
Texto baseado na videocassete didáctica Estudo da Meiose
Uma Edição da
