![]()
เซลล์ส่วนใหญ่ในสิ่งมีชีวิต (ยกเว้นเซลล์เริ่มต้นที่จะแบ่งไปเป็นเซลล์สืบพันธุ์
ซึ่งในเซลล์สัตว์เพศผู้เรียกว่า primary spermatocyte ส่วนในเซลล์สัตว์เพศเมียเรียกว่า primary oocyte) จะแบ่งตัวแบบไมโทซีส เมื่อเซลล์แบ่งตัวแบบไมโทซีสจะแยกออกเป็นเซลล์ใหม่ 2 เซลล์ที่มีจำนวนชุดโครโมโซมหรือชุดสารพันธุกรรมตลอดจนข้อมูลทางพันธุกรรมเหมือนกับเซลล์เริ่มต้นและเหมือนกันเองทุกประการ
การแบ่งเซลล์แบบไมโทซีสเกิดขึ้นตั้งแต่ระยะไซโกตและเกิดอย่างต่อเนื่องตลอดชีวิตเมื่อสิ่งมีชีวิตเจริญเติบโตจะต้องสร้างเซลล์ใหม่เพื่อเพิ่มจำนวนเซลล์เพื่อการรักษาบาดแผล การงอกของเมล็ด หรือเพื่อทดแทนเซลล์ที่ตายไป นอกจากนี้การแบ่งแบบไมโทซีสยังเกิดขึ้นในการสืบพันธุ์แบบไม่อาศัยเพศได้ด้วยโดยเซลล์พ่อแม่ของสัตว์หรือพืชจะแบ่งตัวให้เซลล์ใหม่ซึ่งจะกลายเป็นต้นพืชต้นใหม่หรือสัตว์ตัวใหม่
เป็นที่ทราบกันดีว่าในการแบ่งเซลล์ หรือการทำอะไรก็ตามจากของหนึ่งสิ่งให้กลายเป็นของสองสิ่งที่สามารถทำงานได้อย่างปกตินั้นไม่ใช่การเอาอะไรมาแบ่งครึ่งเพื่อให้แบ่งเป็นสองสิ่ง (เหมือนการผ่าแตงโม) หากแต่ต้องมีการเพิ่มองค์ประกอบสำคัญบางอย่างภายในของสิ่งนั้น ให้มีจำนวนเพิ่มขึ้นเป็นสองเท่า เพื่อแบ่งให้ของชิ้นใหม่หรือเซลล์ใหม่อย่างละครึ่ง
เช่น การสร้างรถยนต์ให้ทำงานได้ไม่ใช่แบ่งจาก 4 ล้อ ให้ได้คันละ 2 ล้อ หรือจากพวงมาลัย
1 อัน
ให้รถ 2 คันอย่างละครึ่งอัน เราต้องเพิ่มจาก 4 ล้อให้เป็น 8 ล้อ จากพวงมาลัยอันเดียวให้กลายเป็น 2 อัน เช่นเดียวกันการแบ่งเซลล์ก็ต้องเพิ่มปริมาณขององค์ประกอบที่สำคัญของเซลล์ เช่น ดีเอ็นเอ เยื่อหุ้มเซลล์ และเซนโทรโซม (centrosome) ซึ่งทำหน้าที่สร้างเส้นใยสปินเดิล (spindle fibers) ตลอดจนโปรตีนต่างๆ จำนวนมากเป็นต้น
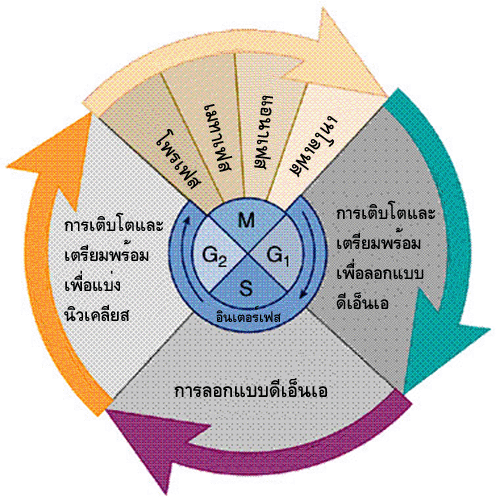
ในหนึ่งรอบของการแบ่งแบบไมโทซีส จะเป็นการเกิดสลับกันระหว่างอินเตอร์เฟส (interphase) และไมโทติกเฟส (mitotic phase) ซึ่งเป็นช่วงของการแบ่งนิวเคลียส(mitosis) และการแบ่งไซโทพลาซึม (cytokinesis) ทั้งนี้เพื่อให้ได้เป็นเซลล์ใหม่
2 เซลล์ ในขั้นตอนของการแบ่งนิวเคลียสยังแบ่งออกเป็นระยะย่อยๆ
ได้อีก 4 ระยะ คือ โพรเฟส(prophase) เมทาเฟส (metaphase) แอนาเฟส (anaphase) และเทโลเฟส (telophase)
![]()
เป็นช่วงของการเตรียมสะสมวัตถุดิบที่จำเป็นสำหรับการสังเคราะห์สารต่างๆ
รวมทั้งการสร้างออร์แกเนลล์เพื่อเตรียมพร้อมสำหรับการแบ่งเซลล์ ระยะนี้แบ่งออกเป็น
3 ระยะย่อยๆได้แก่ ระยะ G1 ระยะ S และระยะ G2 ในทั้ง 3 ระยะนี้เซลล์จะมีการสังเคราะห์โปรตีน คาร์โบไฮเดรต ไขมัน และอื่นๆ เพื่อใช้สร้างองค์ประกอบที่จำเป็นสำหรับเซลล์
เช่น เยื่อหุ้มเซลล์ รวมทั้งออร์แกเนลล์อื่นๆ แต่การลอกแบบดีเอ็นเอ (DNA replication)
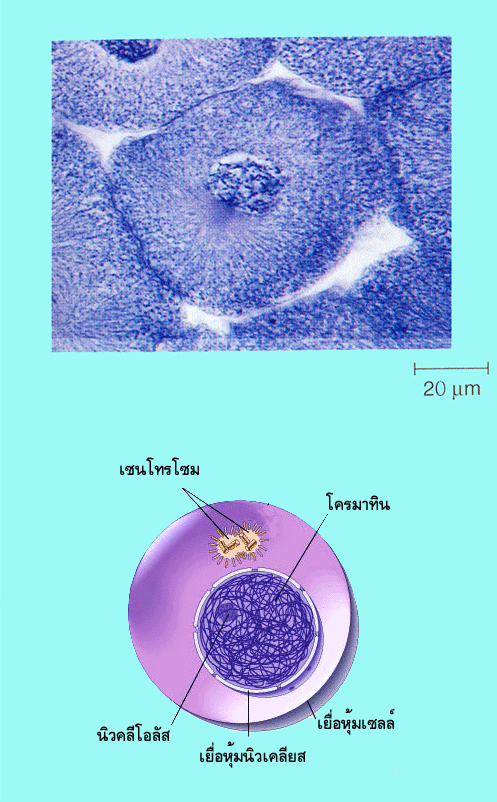
เพื่อเพิ่มปริมาณดีเอ็นเอหรือสารพันธุกรรมนั้นจะเกิดขึ้นในระยะ S เท่านั้น (S ย่อมาจาก synthesisแปลว่าสังเคราะห์) ในระยะอินเตอร์เฟสนี้จะสามารถสังเกตเห็นขอบเขตของนิวเคลียสได้ชัดเจน เนื่องจากเยื่อหุ้มนิวเคลียสยังไม่สลายในไซโทพลาซึม
เซนโทรโซมที่เคยมีหนึ่งอันจะจำลองตัวเองจนมีสองอัน ในเซลล์สัตว์ตรงกลางของเซนโทรโซมจะมีเซนทริโอล (centriole) 2 อันวางตั้งฉากซึ่งกันและกัน ไมโครทูบูล (microtubule) ซึ่งทำหน้าที่เกี่ยวกับการแยกโครโมโซมจะยืดยาวออกมาจากบริเวณเซนโทรโซม ส่วนในเซลล์พืชชั้นสูงจะไม่พบโครงสร้างที่มีลักษณะดังกล่าว ในช่วงท้ายของระยะนี้แม้ว่าการลอกแบบดีเอ็นเอจะเสร็จเรียบร้อยแล้วแต่เราจะไม่สามารถสังเกตเห็นโครโมโซมเป็นแท่งๆ เหมือนเครื่องหมายกากบาท
เนื่องจากดีเอ็นเอยังอยู่ในสภาพโครมาทินที่ยังไม่ขดตัวกันแน่น
ในแต่ละรอบของการแบ่งเซลล์แบบไมโทซีสระยะอินเตอร์เฟสจะใช้เวลายาวนานกว่า
90 เปอร์เซ็นต์ของเวลาทั้งหมดและแม้จะเป็นระยะที่ไม่สามารถสังเกตเห็นการเปลี่ยนแปลงภายใต้กล้องจุลทรรศน์ได้อย่างชัดเจน แต่ก็เป็นระยะที่มีความสำคัญมากเนื่องจากเป็นระยะที่มีการเพิ่มปริมาณดีเอนเอหรือโครมาทินขึ้นเป็นสองเท่า (แต่จำนวนชุดของสารพันธุกรรมยังเท่ากับ 2n เช่นเดิม) เช่น ถ้ากำหนดให้ตัวอักษรภาษาอังกฤษ 1 ตัว แทนโครมาทิน
1 สายในระยะอินเตอร์เฟส สิ่งที่เกิดขึ้นคือ

![]()
มีการเปลี่ยนแปลงปรากฏขึ้นทั้งในนิวเคลียสและไซโทพลาซึม
โดยในนิวเคลียส นิวคลี-โอลัส (nucleolus: เป็นโครงสร้างที่ทำหน้าที่เกี่ยวกับการลอกแบบเพื่อสร้าง
ribosomal RNA และประกอบ ribosomal RNA นี้เข้ากับโปรตีนอีกหลายชนิดเพื่อสร้างเป็นหน่วยย่อย
(subunit)ของไรโบโซม) เริ่มสลายตัวจนไม่ปรากฏให้เห็นเมื่อเซลล์เข้าสู่ช่วงท้ายของระยะนี้
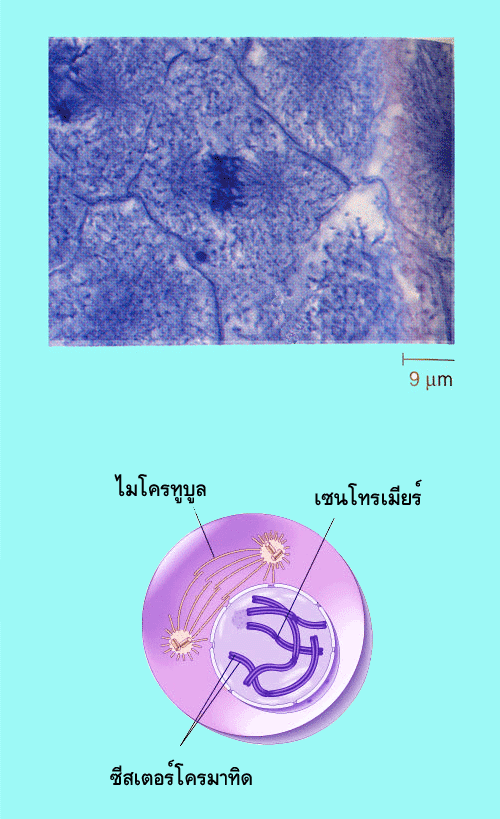
เส้นใยโครมาทินขดตัวมากขึ้นเป็นโครโมโซมแต่ยังไม่ชัดเจนเท่ากับในระยะเมทาเฟส
โครโมโซมแต่ละแท่งประกอบด้วยโครมาทิด 2 เส้น ติดกันอยู่ที่เซนโทรเมียร์ เรียกโครมาทิด
2 เส้นนี้ว่า ซีสเตอร์โครมาทิด (sister chromatids) ในไซโทพลาซึม เซนโทรโซมทั้งสองอันจะเริ่มเคลื่อนไปยังคนละขั้วเซลล์ และในขณะเดียวกันจะมีไมโครทูบูลยืดยาวออกจากบริเวณเซนโทรโซมในช่วงท้ายของระยะนี้เยื่อหุ้มนิวเคลียสเริ่มสลายทำให้ไมโครทูบูลบางส่วน
ยืดเข้ามาในบริเวณนิวเคลียสและจับกับโครโมโซมตรงตำแหน่งไคนีโทคอร์ (kinetochore)ซึ่งเป็นโครงสร้างร่วมกันระหว่างโปรตีนกับดีเอ็นเอบริเวณเซนโทรเมียร์ของซีสเตอร์โครมาทิดแต่ละแท่ง เพื่อความสะดวกและความถูกต้องของการแยกซีสเตอร์โครมาทิดออกจากกันไคนีโทคอร์แต่ละอันจะถูกยึดด้วยไมโครทูบูลที่สร้างจากเซนโทรโซมที่มาจากคนละขั้ว
ของเซลล์ส่วนไมโครทูบูลอีกส่วนหนึ่งจะยึดระหว่างเซนโทร-โซมกับเยื่อหุ้มเซลล์บริเวณใกล้ขั้วเซลล์และไมโครทูบูลอีกส่วนหนึ่งจะยื่นเข้ามาในบริเวณนิวเคลียสแต่ไม่ยึดกับโครโมโซมผลจากการจับระหว่างไมโครทูบูลกับโครโมโซมทำให้เกิดการดึงโครโมโซมไปมา เพื่อให้โครโมโซมเคลื่อนเข้าสู่บริเวณที่เรียกว่าเมทาเฟสเพลท (metaphase plate)
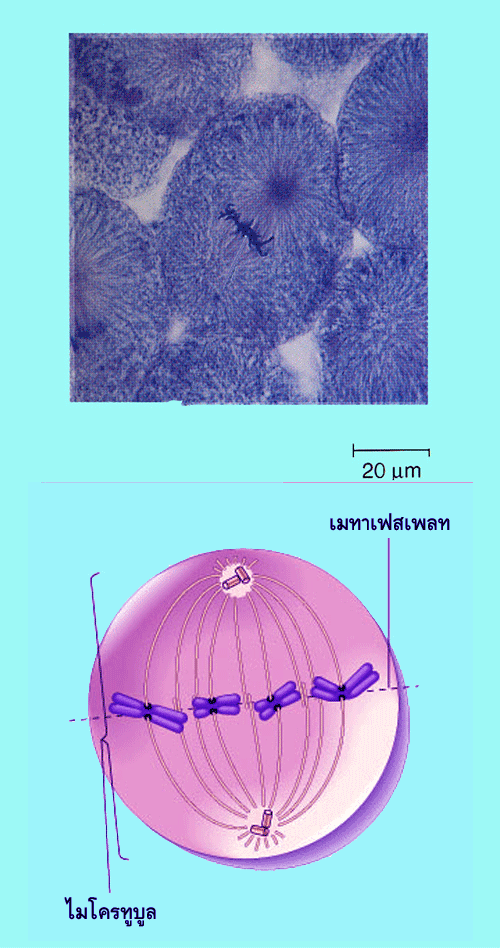
![]()
ในระยะเมทาเฟส เซนโทรโซมทั้งสองอันจะเคลื่อนมาอยู่คนละขั้วของเซลล์ และดึงให้โครโมโซมให้วางตัวในแนวเมทาเฟสเพลทซึ่งอยู่บริเวณกึ่งกลางของเซลล์
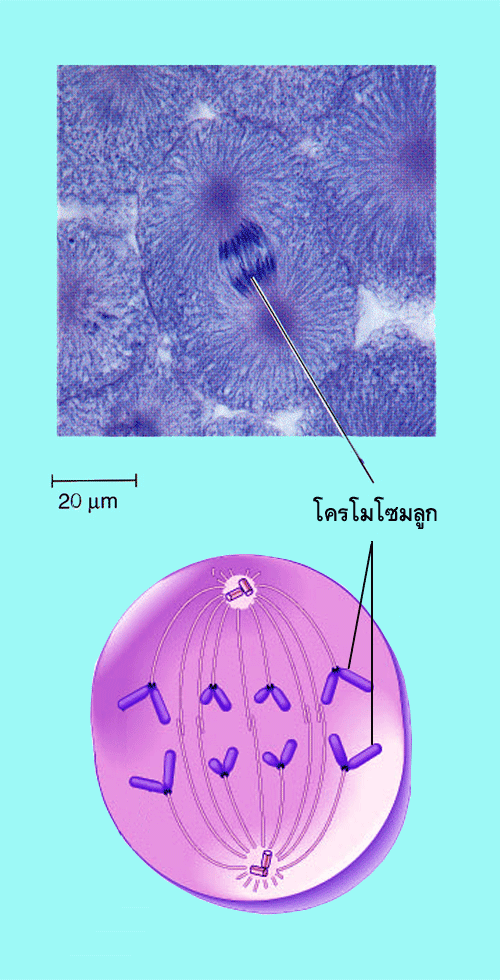
![]()
เป็นระยะที่เริ่มขึ้นเมื่อเซนโทรเมียร์ที่ยึดโครมาทิดสองเส้นให้ติดกันเริ่มแยกออกจากกันอันการแยกจากกันนี้เป็นผลจากการทำงานของไมโครทูบูล การแยกกันของเซนโทรเมียร์ทำให้ซีสเตอร์โครมาทิดแยกออกจากกันเพื่อเคลื่อนไปยังคนละขั้วของเซลล์
![]()
เป็นระยะที่โครโมโซมที่แยกออกจากกันในระยะแอนาเฟส เคลื่อนมาถึงแต่ละขั้วของเซลล์ไมโครทูบูลที่จับอยู่ที่ไคนีโทคอร์สลายตัวปรากฏเยื่อหุ้มนิวเคลียสล้อมรอบกลุ่มของโครมาทิดที่แต่ละขั้วเซลล์จึงเกิดเป็นนิวเคลียสใหม่สองอัน ภายในนิวเคลียสแต่ละอันจะมีโครโมโซม 2 ชุด (2n) โครมาทิดที่เคยขดตัวแน่นตั้งแต่ระยะโพรเฟสเริ่มคลายตัว จนมีลักษณะคล้ายกับโครมาทินในระยะอินเตอร์เฟส นอกจากนี้นิวคลีโอลัสที่ไม่สามารถสังเกตเห็นได้ตั้งแต่ระยะโพรเฟส ก็เริ่มปรากฏให้เห็นอีกครั้ง
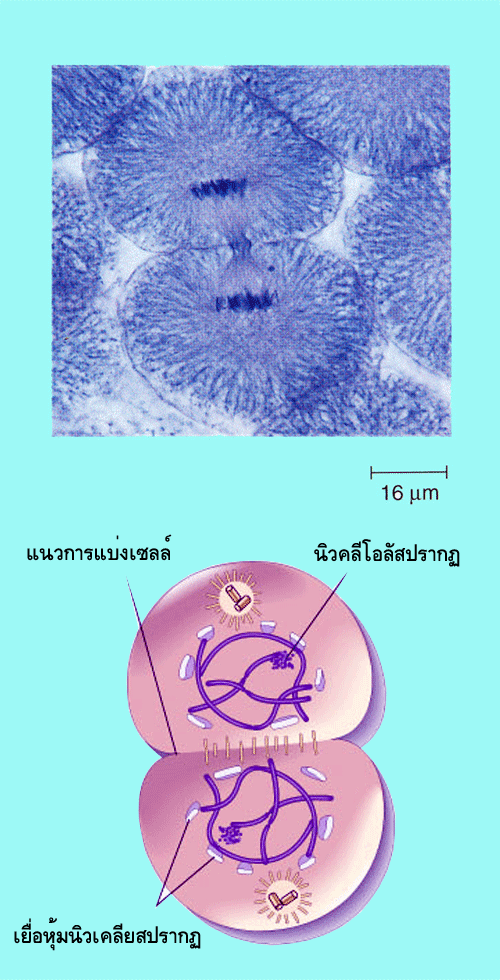
เมื่อสิ้นสุดระยะเทโลเฟสกระบวนการที่เกิดตามมา คือการแบ่งไซโทพลาซึม
ทั้งนี้ก็เพื่อให้เกิดเป็นเซลล์ใหม่ 2เซลล์ซึ่งในนิวเคลียสมีโครโมโซม 2 ชุด (2n) นั่นเอง
ในการแบ่งไซโทพลาซึมนี้เซลล์ใหม่ทั้งสองเซลล์จะได้ปริมาณไซโทพลาซึมเท่ากัน แต่ได้องค์ประกอบในไซโทพลาซึมต่างกัน เนื่องจากความแตกต่างทางโครงสร้างของเซลล์สัตว์และเซลล์พืชทำให้การแบ่งไซโทพลาซึมในเซลล์ทั้ง 2ประเภทนี้มีความแตกต่างกันกล่าวคือ ในเซลล์สัตว์การแบ่งไซโทพลาซึมเกิดจากการคอดเข้าหากันของเยื่อหุ้มเซลล์โดยในช่วงแรกจะสามารถสังเกตได้จากการเกิดร่องหรือรอยย่น(cleavagefurrow)ที่ผิวเซลล์บริเวณแนวที่เคยเป็นเมทาเฟสเพลทการคอดเข้าหากันของเยื่อหุ้มเซลล์เป็นผลจากการหดตัวของโครงสร้างที่เรียกว่าคอนแทรกไทล์ริง(contractilering:เป็นโครงสร้างที่ประกอบด้วยเส้นใยโปรตีนชนิดactinและmyosinII)การหดตัวของโครงสร้างคอนแทรกไทล์ริงเกิดจากการหดตัวของเส้นใยactin(ทำให้มีเส้นผ่าศูนย์กลางลดลง)การหดตัวนี้เป็นผลจากการการทำงานของmyosinIIที่เคลื่อนที่อยู่บนเส้นใยactinนั่นเอง(ถ้าเปรียบเส้นใยactin เป็นเชือกที่มีความยาวเท่ากับเส้นรอบวงของเซลล์ และ myosin II เป็นมด
2 ตัวที่มีปลายของส่วนท้องติดกัน การแบ่งเซลล์จะเกิดขึ้นโดย เชือกที่พันเป็นวงกลมรอบเซลล์จะมีปลาย2 ด้านมาบรรจบกัน ณ จุดหนึ่ง วางมดที่ตัวติดกันไว้จุดนั้นโดยเอาปลายเชือกด้านหนึ่งผูกไว้กับมดตัวที่หนึ่ง ส่วนปลายเชือกอีกด้านหนึ่งผูกไว้กับมดตัวที่สอง เมื่อให้มดที่ตัวติดกันเดินไป
ในทิศตรงกันข้ามกัน (เดินตามเชือก) จะเหมือนเป็นการดึงปลายเชือกให้เหลื่อมกันมากขึ้นซึ่งจะทำให้เส้นรอบวงของเชือกลดลงเรื่อยๆ การลดลงของเส้นรอบวงของเชือกนี้จะรัดให้
เส้นรอบวงของเซลล์ลดลงคือคอดเข้าเรื่อยๆ เช่นกันจนขาดออกจากกันในที่สุด
)
สำหรับเซลล์พืช จะไม่สามารถแบ่งไซโทพลาซึมด้วยวิธีการคอดเข้าหากันของเยื่อหุ้มเซลล์เช่นเดียวกับในเซลล์สัตว์ ดังนั้นจึงไม่เกิดร่องหรือรอยย่น
ในทางกลับกันเซลล์พืชจะแบ่งไซโทพลาซึมโดยการสร้างผนังเซลล์และเยื่อหุ้มเซลล์ขึ้นมาใหม่
โดยตรงบริเวณที่เคยเป็นเมทาเฟสเพลทจะมีการโยงใยของเส้นใยโครงร่างค้ำจุนเซลล์เกิดเป็นโครงร่างคล้ายใยแมงมุม จากนั้นจะมีการนำวัตถุดิบที่ใช้ในการสร้างเยื่อหุ้มเซลล์และผนังเซลล์ใหม่ซึ่งเป็นถุงขนาดเล็กที่ได้มาจาก golgi apparatus ให้ถูกลำเลียงมาตลอดความยาวของไมโครทูบูลจนมาถึงบริเวณของเมทาเฟสเพลท แล้วถุงเล็กๆเหล่านั้นจะรวมกันเป็นแผ่นที่มีขนาดใหญ่ขึ้นเรียกว่า แผ่นกั้นเซลล์ (cell plate)
เมื่อถุงเหล่านั้นรวมตัวกันมากขึ้นจะทำให้แผ่นกั้นเซลล์มีพื้นที่เพิ่มขึ้นเรื่อยๆ จนกั้นเซลล์เดิมให้เกิดเป็นเซลล์ใหม่ 2 เซลล์ในที่สุด