| Osteoporose |
| Fisiologia Having a silent mind is the ultimate and most difficult of life�s goals! |
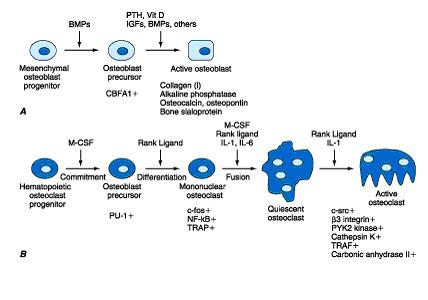
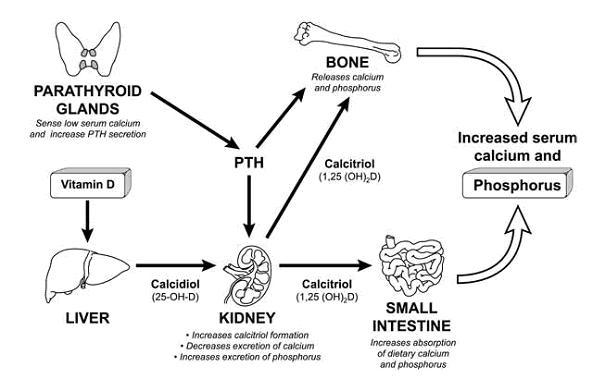
| O esqueleto pode ser dividido em 2 compartimentos principais e tipos de osso: - esqueleto axial - referente � espinha e v�rtebras. Predom�nio de osso trabecular. - esqueleto apendicular - relacionado com os ossos longos dos membros. Nestas �reas o osso � principalmene cortical. a) morfologia H� 2 tipos de osso: trabecular e cortical. A estrutura do osso trabecular � semelhante a um favo de mel, formado por l�minas horizontais e verticais que se interconectam. Isto assegura for�a mec�nica. Eles formam a trama interna dos ossos. A remodela��o �ssea se d� nos envelopes externos e internos de cada trab�cula e ela determina a for�a �ssea. O osso velho � fraco e o novo � mais forte. As trab�culas geralmente n�o apresentam vasos, sendo nutridas a partir da superf�cie. Por isso s�o incapazes de atingir uma espessura maior que 200-300 �m. O osso trabecular predomina na coluna, costelas, cr�nio e esterno, envolto por uma camada cortical. � O osso cortical � a camada compacta que forma a por��o externa dos ossos. Aqui temos os sistemas Haversianos, que s�o orif�cios cil�ndricos de uma profundidade m�dia de 2 a 8 mm e 200 �m de di�metro, em cujo centro h� um canal contendo vasos sangu�neos (Einhorn, 1996). Os ossos corticais e trabeculares contribuem para a for�a �ssea, e cada osso tem sua propor��o normal de componentes corticais e trabeculares. Globalmente, o esqueleto adulto � formado de 80% de osso cortical e 20% trabecular (Crilly et coll, 1981). b) composi��o do osso O osso � constitu�do de: . minerais - em torno de 65%, cristais de hidroxiapatita (Ca10[PO4]6[OH]2) . matriz org�nica - em torno de 35%; sendo 90% col�geno, outras prote�nas e l�pides . c�lulas - sendo osteoblastos, c�lulas de revestimento, oste�citos e osteoclastos . �gua b.1) minerais A estrutura �ssea consiste de uma fase mineral s�lida em �ntima associa��o com a matriz org�nica, onde 90-95% � col�geno tipo I. A fase mineral s�lida � constitu�da de pequenos cristais em forma de placas, em �ntima associa��o com a matriz org�nica. Quimicamente, � uma apatita deficiente em c�lcio, mas contendo outros constituintes como HPO4-, carbonato, citrato, magn�sio, s�dio e fl�or. Existem subst�ncias, tais como as tetraciclinas, polifosfatos e bisfosfonatos que t�m uma afinidade pelo fosfato de c�lcio e portanto pelo osso. Eles s�o depositados preferencialmente nos s�tios de nova forma��o �ssea (Boskey, 1999). A por��o n�o colagenosa cont�m prote�nas derivadas do soro; algumas delas podem atuar na inicia��o da mineraliza��o e na liga��o da por��o mineral com a matriz. O processo de mineraliza��o ocorre rapidamente no in�cio, e nos anos seguintes vai diminuindo a velocidade, um processo denominado mineraliza��o secund�ria. Isto nos explica porque os ossos velhos s�o mais mineralizados e t�m uma maior densidade mineral quando avaliados pela DXA. b.2) matriz org�nica Sua estrutura tridimensional d� ao osso sua for�a tensional. As outras prote�nas n�o-colagenosas, formadas pelos osteoblastos s�o osteonectina, osteocalcina, glicosaminoglicanos, osteopontina, fibronectina, vitronectina, trombospondina e sialoprote�na �ssea (Heinegard et coll, 1999). J� que algumas delas s�o sintetizadas e depositadas em geral apenas no osso, sua excre��o urin�ria e seus n�veis s�ricos s�o usados para se avaliar o turnover �sseo. Na matriz org�nica os osteoblastos tamb�m sintetizam uma grande quantidade de fatores de crescimento, tais como TGFbeta; (transforming growth factor beta;), BFGF (basic fibroblast growth factor), IGF I e II (insulin-like growth factors I and II), e diversas prote�nas morfogen�ticas, os quais provavelmente desempenham importante papel ap�s sua libera��o durante a reabsor��o �ssea na modula��o local da forma��o �ssea. b.3) c�lulas b.3.1) osteoblastos Os osteoblastos, derivados das c�lulas mesenquimais, sintetizam a matriz �ssea. Esta � uma subst�ncia essencial para a posterior mineraliza��o por ader�ncia, de cristais de hidroxiapatita de c�lcio, magn�sio, pot�ssio, s�dio e carbonato a fibrilas de col�geno, individualmente. Eles apresentam um citoplasma fortemente basof�lico, com um proeminente complexo de Golgi, e a membrana plasm�tica possui receptores para o paratorm�nio. Os osteoblastos produzem o col�geno tipo I, inicialmente secretado como um precursor, o qual cont�m extens�es pept�dicas tanto na por��o N-terminal quanto nas termina��es carbox�licas da mol�cula. Quando maduras, as mol�culas de col�geno se re�nem em fibrilas, as quais se inter-conectam pela forma��o de piridinolina e deoxipiridinolina cross-links. Os osteoblastos tamb�m inibem a reabsor��o �ssea (Vitte et coll, 1996) atrav�s da s�ntese de um inibidor da reabsor��o osteocl�stica. Tamb�m produzem grandes quantidades de fosfatase alcalina, expressando a mesma isoforma encontrada no f�gado e rins. Ainda, eles possuem receptores nucleares para estrog�nio (Eriksen et coll, 1988), o que sugere a a��o estrog�nica direta sobre tais c�lulas. Em conseq��ncia do espa�o de tempo entre a forma��o da matriz e sua calcifica��o surge uma camada de matriz oste�ide n�o mineralizada sob os osteoblastos, a qual diminui em espessura quando a taxa de forma��o de matriz �ssea diminui, mas � ampliada quando a mineraliza��o est� atrasada como na osteomal�cia. Desenvolvimento dos osteoblastos e osteoclastos. Horm�nios, citocinas e fatores de crescimento que controlam a prolifera��o celular e a diferencia��o s�o mostrados. Tamb�m fatores de transcri��o e outros marcadores espec�ficos para os v�rios est�gios de desenvolvimento. BMPs, bone morphogenic proteins; PTH, parathyroid hormone; Vit D, vitamin D; IGFs, insulin-like growth factors; CBFA1, core bind-ing factor A1; M-CSF, macrophage colony stimulating factor; PU-1, a monocyte- and B lymphocyte-specific ets family transcription factor; NFkB, nuclear factor kB; TRAF, tumor necrosis factor receptor-associated factors; RANK ligand, receptor activator of NFkB ligand; IL-1, interleukin-1; IL-6, interleukin-6. (Modified from Suda et al.) b.3.2) c�lulas de revestimento Se os osteoblastos n�o est�o formando osso eles se tornam amplos e s�o denominados oste-oblastos em repouso ou c�lulas de revestimento. Os osteoblastos ativos e em repouso formam uma membrana na superf�cie do osso, a qual pode ser importante na constitui��o de uma barreira sangue-osso. b.3.3) oste�citos Em um dado momento alguns osteoblastos param de sintetizar matriz e penetram no osso, quando s�o chamados oste�citos. Eles se localizam nas lacunas e se conectam por processos cito-plasm�ticos entre eles pr�prios e com os osteoblastos e as c�lulas de revestimento. Provavelmente os oste�citos ajudam a regular os minerais no plasma, em especial o c�lcio. Devido � sua localiza��o � poss�vel que participem na resposta ao est�mulo mec�nico transformando-o em mudan�as na forma��o e reabsor��o �ssea. b.3.4) osteoclastos Os osteoclastos s�o derivados de c�lulas mononucleares da medula �ssea (Roodman, 1996) (pr�-osteoclastos), e sua produ��o � estimulada pelas citocinas (Mazlam et Hodgson, 1992) locais e inibida pela calcitonina. Eles s�o geralmente multinucleados e se enfileiram nas superf�cies de tecido �sseo cortical ou trabecular, ou em depress�es chamadas lacunas de Howship, ou dentro do osso cortical. No citoplasma h� grande quantidade de complexos de Golgi em torno de cada n�cleo. O 17 ?-estradiol pode ter um efeito indireto sobre os osteoclastos atrav�s do controle da produ��o de v�rias citocinas nos osteoblastos e nos mon�citos (Horowitz, 1993). Estas citocinas t�m uma fun��o importante na reabsor��o �ssea pelos osteoclastos (Horowitz, 1993). Os osteoclastos s�o localizados na extremidade das unidades de remodela��o, perfurando os canais vasculares onde os novos osteons ser�o formados. Sua principal fun��o � a desmineraliza��o �ssea e a digest�o da matriz do osso. Esta reabsor��o �ssea se d� em um micro-ambiente localizado entre a c�lula e o osso, delimitado por um anel rico em actina que fornece grande ader�ncia entre a membrana celular e a matriz �ssea. Esta regi�o especializada da c�lula � denominada clear zone, e envolve receptores de membrana celular denominados integrinas que reconhecem especificas se-q��ncias de pept�deos na matriz. Cobrindo este micro-ambiente est� outra por��o especializada da membrana celular, a borda dentada, a qual secreta dois diferentes produtos que levam � destrui��o �ssea. O primeiro, s�o os �ons H+, que dissolvem os cristais de hidroxiapatita, sendo originados a partir do H2CO3 em conseq��ncia da a��o da anidrase carb�nica e s�o secretados via bomba de pr�tons. O segundo produto inclui v�rias enzimas lisossomais, como as catepsinas e em especial a catepsina K, e as colagenases, tais como a matrix metalloproteinase 1, que digerem os componentes da matriz. b.3.5) outras c�lulas A medula �ssea cont�m ainda c�lulas hematopoi�ticas, do sistema imune e estromais, as quais s�o importantes na forma��o dos osteoclastos e na interrela��o com as c�lulas �sseas atrav�s de v�rias citocinas, al�m de estarem envolvidas na modula��o da forma��o e da reabsor��o �ssea. c) modela��o e remodela��o Uma vez que o osso se encontre totalmente formado, sua forma e estrutura s�o continuamente renovadas e modificadas pelos processos de modela��o e remodela��o. Na modela��o, a qual tem lugar principalmente durante o crescimento, novo osso � formado em uma localiza��o diferente daquela onde o osso foi destru�do, resultando em mudan�a na forma do esqueleto. Isto permite n�o apenas o desenvolvimento de uma arquitetura normal durante o crescimento, mas tamb�m a modula��o desta arquitetura no adulto quando as condi��es mec�nicas se modificam. Al�m disso, esta � a causa do aumento em tamanho dos ossos durante a vida. Modela��o � o principal processo atrav�s do qual o esqueleto pode aumentar seu volume e massa. Na remodela��o, que � o principal processo no adulto, os 2 processos est�o associados em espa�o e tempo, de maneira que n�o h� mudan�a na forma do osso. A modela��o e a remodela��o promovem a substitui��o do osso velho pelo novo, permitindo a manuten��o da integridade mec�nica do esqueleto. A taxa de remodela��o se d� entre 2 a 10% da massa esquel�tica ao ano. Este pode ser um processo sist�mico ou localizado. O osso trabecular, representando 20% da massa esquel�tica, � respons�vel por 80% do turnover, enquanto o c�rtex, que representa 80% do osso, participa em 20% do turnover. E j� que a perda �ssea ocorre mais rapidamente no osso trabecular temos que as fraturas tendem a surgir principalmente nas v�rtebras, colo do f�mur e nas termina��es dos ossos longos. O ciclo de remodela��o �ssea consiste de 4 passos: a) ativa��o: o processo se inicia atrav�s de sinais f�sicos ou hormonais que fazem com que c�lulas precursoras, mononucleares, derivadas da medula se agrupem na superf�cie �ssea, onde elas se fundem em osteoclastos multinucleados, com citoplasma pleno de mitoc�ndrias, ribossomos livres, ves�culas, complexo de Golgi e lisossomas. Este processo � mediado pelos osteoblastos, que liberam v�rios mediadores qu�micos. Estes, por sua vez estimulam a s�ntese de v�rios fatores que promovem a prolifera��o de c�lulas hema-topoi�ticas. Um receptor nos osteoclastos e seus precursores, o receptor ativador do NFkB (RANK), interage com um grupo molecular nos osteoblastos (RANK-L), para estimular a forma��o e atividade dos osteoclastos. Os osteoblastos tamb�m produzem a osteoprotegerina, que � um fator inibit�rio, o qual quando em excesso bloqueia a reabsor��o �ssea (Hansen, 1991), e se em baixos n�veis promove a osteoporose. Os osteoblastos ainda s�o respons�veis pela produ��o do MCSF (macrophage colony-stimulating factor), o qual estimula a replica��o dos precursores do osteoclasto. b) reabsor��o: no osso cortical, os osteoclastos se fundem e iniciam o processo de escava��o de um t�nel. Eles criam um ambiente �cido, dissolvendo e digerindo a matriz org�nica e mineral do osso velho. Os principais horm�nios que modulam a reabsor��o �ssea s�o o PTH e a 1,25(OH)2D, ace-lerando o processo, e a calcitonina, que o diminui. Os estrog�nios nas mulheres e a testosterona nos homens inibem a reabsor��o �ssea. Assim � que na menopausa, bem como na orquiectomia, ocorre um aumento na reabsor��o �ssea, mediado possivelmente em parte por um aumento em IL-1, IL-3, IL-6 e IL-11, fator de necrose tumoral alfa e beta, e prostaglandinas (Martin et coll, 1998; Manolagas et Jilka, 1995). c) invers�o: a reabsor��o cessa quando a cavidade atinge certa profundidade; c�lulas derivadas de mon�citos formam uma superf�cie de cimento, que previne a eros�o �ssea adicional. d) forma��o: quando a reabsor��o termina, inicia-se a forma��o �ssea. Libera��o local de mediadores qu�micos, provavelmente TGFb e IGF1, atraem pr�-osteoblastos que se transformam em osteo-blastos e iniciam o processo de forma��o �ssea pela secre��o de col�geno novo e outros constituintes da matriz. Reabsor��o e forma��o est�o completas em 8 a 12 semanas, e em mais algumas semanas se completa a mineraliza��o. Sob condi��es normais, h� um equil�brio e a quantidade de osso permanece constante. Entretanto, a remodela��o �ssea, como outros processos biol�gicos, n�o � total-mente eficiente, podendo ocorrer um desequil�brio. d) metabolismo dos sais minerais d.1) c�lcio Um total de 1-2 kg de c�lcio est� presente em um adulto, sendo que pelo menos 98% est� no esqueleto. O c�lcio da fase mineral na superf�cie dos cristais est� em equil�brio com aquele do liquido extracelular, mas apenas uma pequena fra��o daquele total (0,5%) pode ser convertida. Sua concentra��o m�dia no plasma � de 9,0 mg%. No plasma o c�lcio est� presente em 3 formas: a) 40% combinados com as prote�nas, principalmente a albumina; b) 10% difundem-se pela membrana capilar, mas encontram-se combinados com outras subst�ncias do plasma e dos l�quidos intersticiais, como citrato e fosfato, de forma n�o-ioniz�vel; c) os outros 50% s�o difund�veis pela membrana capilar, e ioniz�veis. � este c�lcio i�nico que atua no cora��o, sistema nervoso e ossos, e seus n�veis s�o mantidos dentro de pequena margem de varia��o. O c�lcio aparece no fluido intracelular em menor quantidade que no extracelular. A diferen�a entre estas concentra��es se faz pelo transporte ativo. Ao avaliarmos o c�lcio s�rico total estamos medindo ao mesmo tempo o c�lcio ionizado junto com aquele que se liga �s prote�nas e o que se une a outros componentes. Em geral isto pode ser feito j� que � mais f�cil do que dosar o c�lcio ionizado, e em pacientes normais o c�lcio total reflete o ionizado. A concentra��o das prote�nas s�ricas � fundamental na determina��o da concentra��o i�nica, sendo que a maioria dos �ons se liga � albumina. A maioria do c�lcio intracelular est� sob a forma de complexos insol�veis. A quantidade de c�lcio livre na c�lula � muito baixa (0.1 � mol/l), e assim o gradiente entre o c�lcio plasm�tico e intracelular � de 10.000:1 (Palmieri, 1995). Os n�veis de c�lcio no l�quido extracelular s�o mantidos constantes atrav�s de mecanismos que continuamente acrescentam e retiram c�lcio. O c�lcio atinge o plasma seja pela absor��o intestinal ou pela reabsor��o �ssea. O c�lcio deixa o l�quido extracelular pela secre��o gastrintestinal, secre��o urin�ria, deposi��o �ssea e perdas pelo suor. Menos da metade do c�lcio da dieta � absorvido (Heaney, 1993) principalmente no delgado proximal. O transporte ativo � mais importante no intestino proximal (Lau et coll, 1989) e a absor��o limitada pela difus�o no intestino distal. Ambos os processos s�o influenciados pela vitamina D, e tamb�m a absor��o � diferente de acordo com a forma do c�lcio. O c�lcio � pouco absorvido no intestino, mas o fosfato tem �tima absor��o, exceto quando h� excesso de c�lcio na dieta. Pode ocorrer forma��o de fosfato de c�lcio, quase insol�veis, que n�o s�o absorvidos, e saem nas fezes. 5/6 do c�lcio ingerido sai pelas fezes; o outro 1/6 na urina. A efic�cia da absor��o de c�lcio no intestino diminui � medida que a ingest�o aumenta (mecanismo protetor contra intoxica��o), contudo � superado por uma ingest�o > 4 g/dia, levando � les�o renal e deposi��o ect�pica. A acidez g�strica � importante na biodisponibilidade do c�lcio, e em pacientes portadores de patologia g�strica com diminui��o desta acidez haver� menor absor��o deste elemento (Pak et coll, 1989). Tamb�m em n�vel intestinal, patologias que levam � maior alcaliniza��o regional podem interferir na solubiliza��o do c�lcio. Assim, o suco de laranja fortificado com c�lcio apresenta um maior balan�o positivo do que o leite e o queijo pasteurizado tamb�m fortificados. Por dia ocorre uma filtra��o glomerular de c�lcio em torno de 8,0 g, onde 7,8 g s�o reabsorvidos, sendo que 70% no t�bulo proximal, 20% na al�a ascendente, 5-10% no t�bulo distal, e o restante no t�bulo coletor. A reabsor��o tubular renal e a reabsor��o osteocl�stica promovem grande controle sobre o c�lcio s�rico, e a a��o do PTH sobre o t�bulo distal se d� de maneira bastante r�pida, em quest�o de minutos. No entanto para que ocorram ajustes nas taxas de absor��o intestinal via eixo PTH-vitamina D s�o necess�rias 24-48 h. Atentar para o fato de que na acidose metab�lica ocorre maior perda de c�lcio pela inibi��o tubular renal de c�lcio relativa � de s�dio no t�bulo distal. A maioria do c�lcio do filtrado glomerular � reabsorvida nos t�bulos proximais e ramos ascendentes das al�as de Henle. Nos t�bulos distais e ductos coletores a reabsor��o � seletiva, e depende da concentra��o de c�lcio sang��neo. A manuten��o do balan�o de c�lcio depende da efici�ncia da absor��o intestinal. E o aumento da reabsor��o �ssea pode proteger contra a defici�ncia de c�lcio no liquido extracelular, mesmo em estados cr�nicos de balan�o negativo. A ingest�o e absor��o respondem por apenas 25% da varia��o no balan�o de c�lcio, enquanto as perdas urin�rias respondem por 50%. V�rios fatores diet�ticos podem levar a maior perda de c�lcio urin�rio, principalmente a prote�na animal e o s�dio. A ingest�o de c�lcio de lactentes amamentados apenas com leite materno nos 6 primeiros meses � de 250 a 300 mg/dia, com uma absor��o fracionada de c�lcio de 55% (compar�vel com a absor��o no leite de soja) (Gertz et coll, 1995); menor absor��o fracionada (30%) se acha nas f�rmulas com leite de vaca. Em lactentes entre 6 e 12 meses, a ingest�o varia de 400 a 700 mg/dia; baixo peso ao nascer requer maior ingest�o. De 3 a 10 anos, ingest�o acima de 800 mg/dia provoca maiores taxas de ac�mulo nos ossos. Defici�ncia de c�lcio na inf�ncia leva a hipoplasia do esmalte e forma��o de c�ries. O ac�mulo de c�lcio no osso durante a pr�-adolesc�ncia � de 140-165 mg/dia, e pode ser t�o alto quanto 400-500 mg/dia na puberdade; a absor��o intestinal � muito eficiente e estimada em 40%. A ingest�o de c�lcio de 1.200-1.500 mg/dia nesta fase pode resultar em maior pico de massa �ssea no adulto (Heaney, 1995). Entretanto, apenas 25% dos garotos e 10% das garotas na adolesc�ncia costumam ingerir c�lcio nas quantidades recomendadas, devida � pobre ingest�o de frutas e verduras, e � alta ingest�o de bebidas com baixo teor de c�lcio, como os refrigerantes. O uso do c�lcio na alimenta��o di�ria � mais importante durante o crescimento e desenvolvimento para se adquirir o pico de massa �ssea determinado geneticamente. � tamb�m importante na manuten��o da massa �ssea na velhice. Uma vez atingido o pico, o turnover �sseo torna-se est�vel em homens e mulheres; nestas, as taxas de reabsor��o aumentam e a massa �ssea cai seguindo a redu��o de estrog�nio na menopausa. Nesta fase imediatamente ap�s a menopausa o c�lcio n�o tem muito efeito, j� que a perda �ssea se relaciona mais com a queda de estrog�nio do que com algum d�ficit em nutrientes. Em maiores de 65 anos, a absor��o intestinal de c�lcio � menor, pela defici�ncia de estr�genos na mulher, e � redu��o da produ��o renal de 1,25-di(OH)vitamina D associada � idade (Bell et coll, 1991). Entre 25-50 anos, as mulheres devem manter uma ingest�o de 1.0 g/dia, e na p�s-menopausa, 1,5 g/dia. Mas apenas metade desta popula��o ingere esta quantidade adequada. A gravidez � um estresse fisiol�gico sobre a homeostase esquel�tica da m�e; n�o h� associa��o entre paridade e massa �ssea. Mulheres na gravidez e quando amamentando devem ingerir ao menos 1,2 g/dia de c�lcio; e se forem adolescentes ingerir pelo menos 1.5 g/dia. Na lacta��o, 160-300 mg/dia de c�lcio materno � gasto na produ��o do leite materno. Um beb� a termo acumulou em torno de 30 g de c�lcio durante a gesta��o, a maioria tendo sido assimilada em seu esqueleto no terceiro trimestre. � importante compreender que a preven��o atrav�s de uma alimenta��o suficientemente ri-ca em c�lcio � a melhor arma contra a osteoporose, uma arma mais eficaz que a suplementa��o massiva com c�lcio colocada em pr�tica uma vez que se perceba o desgaste �sseo. d.2) vitamina D A 1,25-di(OH)vitamina D estimula o transporte ativo de c�lcio no delgado e c�lon. Os raios ultravioletas (Chapuy, 1992) atuam na pele sobre o 7-dehydrocholesterol formando a vitamina D end�gena; mas ela deve ser convertida no f�gado (hidroxilada) em 25-hidroxicolecalciferol (Chapuy et coll, 1983). Esta � uma convers�o controlada, o que possibilita conservar vitamina D para o futuro, pois ap�s convertida ela s� vai durar 2 a 5 dias no organismo, e na forma de vitamina D pode ser armazenada no f�gado por meses. Com o envelhecimento ocorre uma redu��o significativa nos estoques da pele do precursor 7-dehydrocholesterol, resultando numa acentuada redu��o na produ��o de vitamina D3. A capacidade da pele de uma pessoa de 70 anos de idade em produzir vitamina D3 � de apenas 30% daquela de um adulto (Holick, 1996). Enfim, nos rins h� a convers�o para 1,25 diidroxicolecalciferol, que � a forma ativa da vitamina D. Assim na aus�ncia dos rins a vitamina D � quase totalmente ineficaz. A convers�o final depende da a��o do PTH, fosfato e c�lcio. O �on c�lcio tem pequeno efeito para impedir a convers�o do 25-hidroxicolecalciferol em 1,25-diidroxicolecalciferol. A concentra��o plasm�tica de vitamina D3 � inversamente influenciada pela de c�lcio do plasma. A vitamina D3 leva � forma��o de uma prote�na fixadora de c�lcio a qual permanece nas c�lulas por semanas ap�s a remo��o do 1,25-diidroxicolecalciferol do organismo, causando assim um prolongado efeito sobre a absor��o de c�lcio (Guyton, 1989). Tamb�m o 1,25-diidroxicolecalciferol causa a for-ma��o de uma ATPase, estimulada pelo c�lcio, no bordo em escova das c�lulas epiteliais e origina uma fosfatase alcalina nas c�lulas epiteliais. Na aus�ncia de 1,25 (OH)2D3, apenas 7% do c�lcio ingerido ser� absorvido, refletindo a absor��o de c�lcio que n�o � mediada por horm�nios. Estudos em receptores de vitamina D sugerem um risco maior de osteoporose em indiv�duos apresentando espec�ficos halotipos, comum em caucasianos e raros em japoneses, o que diminui a efic�cia da vitamina D Nos EUA � comum a suplementa��o e a fortifica��o do leite com esta vitamina, permitindo a ingest�o adequada na inf�ncia. Na adolesc�ncia, quando o consumo destes produtos diminui, a ingest�o di�ria se torna inadequada, numa fase de m�ximo desenvolvimento �sseo. Recomenda-se 400-600 UI/dia para os adultos. A associa��o de c�lcio, 1,0 g/dia com 1,25(OH)2D, 500 UI/dia por um per�odo de 3 anos previne fraturas em n�vel vertebral de mulheres com osteoporose (Tilyard, 1992). A incid�ncia anual de fraturas de quadril aumenta exponencialmente com a idade, tornando-se um problema s�rio ap�s os 75 anos. A suplementa��o com c�lcio e vitamina D3 previne este aumento de risco relacionado com a idade, n�o apenas para fraturas de quadril mas tamb�m para outras fraturas n�o-vertebrais (Nydegger et coll, 1991). d.3) magn�sio Em torno de 60% do magn�sio corporal se acha no esqueleto, 27% na musculatura, e 6 a 7% em outras c�lulas, e menos de 1% est� fora das c�lulas (Shils, 1997). Altas doses de zinco como suplemento parece interferir com a absor��o de magn�sio. Uma suplementa��o com zinco de 142 mg/dia em adultos sadios significativamente diminuiu a absor��o de magn�sio (Spencer, 1994). A prote�na da dieta tamb�m pode afetar a absor��o de magn�sio. Um estudo mostrou que a absor��o de magn�sio est� diminu�da quando a ingest�o prot�ica for menor que 30 g/dia, e que aumenta se a ingest�o for maior (Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes, 1997). A forma ativa da vitamina D (calcitriol) pode aumentar a absor��o intestinal de magn�sio em pequena quantidade. Entretanto, a absor��o de magn�sio n�o parece ser calcitriol-dependente como a absor��o de c�lcio e fosfato. Altas ingest�es de c�lcio n�o parecem afetar o balan�o do magn�sio. J� n�veis inadequados de magn�sio no sangue s�o conhecidos por resultarem em baixos n�veis de c�lcio sang��neo, resist�ncia ao PTH, e resist�ncia a alguns dos efeitos da vitamina D (Shils, 1999). A defici�ncia de magn�sio em indiv�duos sadios consumindo uma dieta balanceada � rara pois o magn�sio � abundante tanto nos alimentos de origem vegetal quanto animal e porque os rins conseguem limitar a excre��o urin�ria de magn�sio quando a ingest�o for baixa (Brody, 1999). Uma pobre ingest�o, problemas gastrintestinais como diarr�ia prolongada e processos inflamat�rios intestinais, e uma perda urin�ria aumentada de magn�sio podem contribuir para a deple��o de magn�sio, a qual � freq�entemente encontrada em alcoolistas. Diversos estudos mostram que a popula��o idosa tem relativamente menores ingestas di�rias de magn�sio. J� que a absor��o intestinal de magn�sio tende a diminuir e a excre��o urin�ria a aumentar nos idosos, ingest�es sub-�timas de magn�sio na dieta podem aumentar o risco de deple��o deste elemento nesta faixa et�ria ((Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes, 1997). Na defici�ncia de magn�sio, o primeiro sinal � a hipomagnesemia. Com o evoluir do quadro ocorre tamb�m hipocalcemia mesmo com ingest�o adequada de c�lcio na dieta. A hipocalcemia � persistente apesar do aumento na secre��o de PTH. Geralmente o aumento na secre��o de PTH resulta na mobiliza��o de c�lcio �sseo e normaliza��o da calcemia. Com o evoluir da deple��o de magn�sio, a secre��o de PTH diminui. Junto com a hipomagnesemia, sinais de severa defici�ncia de magn�sio incluem hipocalcemia, hipocalemia, reten��o de s�dio, baixos n�veis sang��neos de PTH, sintomas neurol�gicos e musculares (tremores, espasmos musculares, tetania), perda de apetite, n�usea, v�mitos e altera��es de personalidade (Shils, 1999). Quantidades di�rias recomendadas (RDA) para o magn�sio: . homens de 19-30 anos: 400 mg/dia . mulheres de 19-30 anos: 310 mg/dia . homens acima de 30 anos: 420 mg/dia . mulheres acima de 30 anos: 320 mg/dia Embora a diminui��o da densidade mineral �ssea seja a caracter�stica prim�ria da osteoporose, outras mudan�as osteopor�ticas na matriz de col�geno e nos componentes minerais do osso podem resultar em ossos fr�geis e mais suscept�veis a fraturas. O magn�sio responde por 1% do mineral �sseo e � conhecido por influenciar tanto a matriz quanto o metabolismo mineral �sseo. Enquanto decresce o conte�do de magn�sio no osso, cristais �sseos se tornam maiores e mais fr�geis (Sojka et Weaver, 1995). A hipomagnesemia resulta em baixos n�veis de c�lcio sang��neo, resist�ncia ao PTH e a alguns efeitos da vitamina D, o que em conjunto pode levar a aumento na perda �ssea. Um estudo recente com mais de 900 idosos de ambos os sexos observou que uma ingest�o diet�tica aumentada de magn�sio se associa com um aumento na densidade mineral �ssea no quadril nos dois sexos. Entretanto, como o magn�sio e o pot�ssio est�o presentes em v�rios dos mesmos alimentos estudados seus efeitos n�o t�m como serem separados (Tucker, 1999). Fontes alimentares de magn�sio: Por ser o magn�sio parte da clorofila, os vegetais de folhas verdes s�o ricos em magn�sio. Gr�os n�o refinados e castanhas tamb�m t�m alto conte�do de magn�sio. Carnes e leite apresentam valores intermedi�rios, enquanto alimentos refinados geralmente s�o pobres em magn�sio. A �gua � uma fonte vari�vel deste sal. Efeitos adversos n�o s�o vistos com a ingest�o de magn�sio presente em alimentos. Mas eles podem aparecer com o uso de suplemento de sais de magn�sio. A manifesta��o inicial � a diarr�ia, e a hipermagnesemia pode tamb�m levar � queda da press�o arterial. Os indiv�duos com les�o renal s�o mais propensos a tais altera��es. Intera��es de drogas: O magn�sio interfere com a absor��o de digoxina e nitrofuranto�na, potencialmente reduzindo sua efic�cia. Os bisfosfonatos e o magn�sio devem ser ingeridos no m�nimo 2 horas aparte, assim a absor��o dos bisfosfonatos n�o ser� inibida. J� que o magn�sio endovenoso pode acentuar os efeitos de certos relaxantes musculares usados durante anestesia, � aconselh�vel evitar suplementos de magn�sio algumas horas antes de cirurgias. Altas doses de furosemida ou hidroclorotiazida durante longos per�odos pode resultar em deple��o de magn�sio (Silverman, 1999), d.4) f�sforo Em torno de 32 mol (1 kg) est� presente no adulto normal, sendo que 85% se acha no esqueleto (Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes, 1997). O f�sforo � um componente �sseo importante na forma de um sal de fosfato de c�lcio chamado hidroxiapatita. Intera��es: . c�lcio e vitamina D: o f�sforo da dieta � prontamente absorvido no intestino delgado, e qualquer excesso absorvido � excretado na urina. A regula��o dos n�veis sang��neos de c�lcio e f�sforo se interrelaciona atrav�s das a��es do PTH e da vitamina D. Homeostasia do c�lcio e f�sforo Prote�nas sens�veis ao c�lcio nas gl�ndulas paratire�ides detectam os n�veis s�ricos de c�lcio. Em resposta a pequenas quedas na calcemia as gl�ndulas paratire�ides secretam PTH, o qual estimula a atividade da enzima 1-hydroxylase nos rins, resultando no aumento da produ��o de calci-triol, a forma biologicamente ativa da vitamina D. O calcitriol ativa o sistema de transporte dependente de vitamina D no intestino delgado, aumentando a absor��o do c�lcio e do f�sforo diet�tico. O calcitriol e o PTH agem no esqueleto para aumentar a mobiliza��o de c�lcio e f�sforo para a circula��o. Nos rins calcitriol e PTH aumentam a reabsor��o de c�lcio e aumentam a excre��o de f�sforo. O aumento da excre��o de f�sforo eleva os n�veis s�ricos de c�lcio trazendo-os ao normal pois altos n�veis sang��neos de fosfato suprimem a convers�o de vitamina D em sua forma ativa nos rins (Bringhurst, 1998). Os n�veis de f�sforo total s�o mais altos em crian�as e na p�s-menopausa. Ocorre uma varia��o circadiana dos n�veis de f�sforo, sendo mais baixos entre 9 e 12 h da manh�, e apresentando valores mais altos no come�o da tarde e ap�s a meia-noite. Durante uma deple��o de fosfato, a fosfat�ria diminui antes que haja decl�nio do f�sforo s�rico. O maior controle nos n�veis de f�sforo � exercido a n�vel renal. O f�sforo filtrado pelo glom�rulo � reabsorvido largamente no t�bulo proximal, e apenas 10-15% � excretado. Lembrar que a excre��o urin�ria do f�sforo normalmente reflete a ingest�o diet�tica. Quantidades di�rias recomendadas (RDA): . adultos de 19 - 50 anos: 700 mg/dia . adultos maiores que 50 anos: 580 mg/dia Fontes alimentares: O f�sforo est� presente na maioria dos alimentos j� que ele � um componente importante dos seres vivos. Leite e derivados e as carnes de origem animal s�o ricas fontes de f�sforo. O f�sforo � tamb�m um componente de v�rios polifosfatos usados como aditivos alimentares, e se encontra na maioria dos refrigerantes como �cido fosf�rico. O f�sforo presente nos gr�os de vegetais se encontra na forma de fosfato chamada �cido f�tico ou fitato. Em torno de 50% do f�sforo a partir do fitato � aproveitado pelos seres humanos. Os fermentos possuem fitases, assim os gr�os integrais incorporados aos p�es fermentados t�m um maior potencial de aproveitamento do f�sforo que os gr�os incorporados aos cereais matinais (Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes, 1997). Toxicidade: Intera��es medicamentosas: Os anti�cidos contendo alum�nio reduzem a absor��o do f�sforo diet�tico ao formarem fos-fato de alum�nio, o qual n�o � absorvido. Quando consumido em altas doses, os anti�cidos conten-do alum�nio podem produzir hipofosfatemia severa, tanto quanto agravar a defici�ncia de fosfato por outras causas. |
 |
 |